Living off the fat of the plant: Powdery mildew fungi thrive on host thylakoid lipids
Jan Wilhelm Hübbers

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
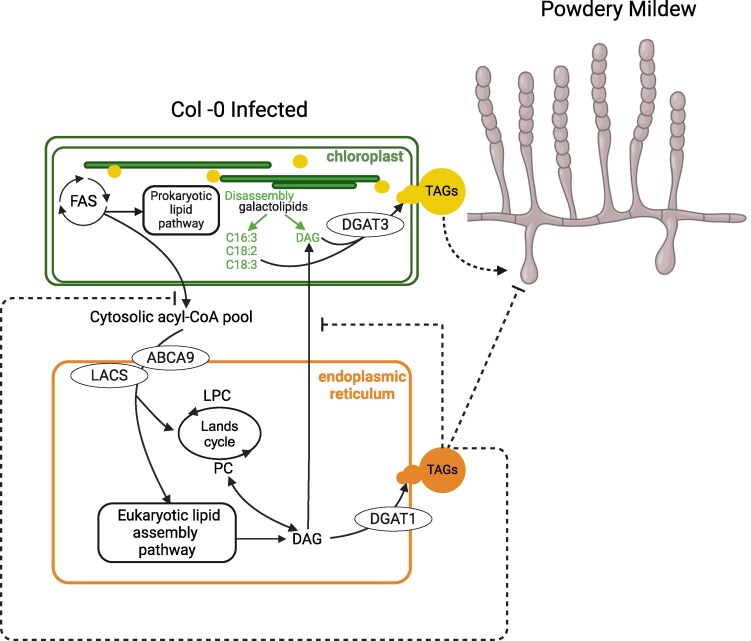 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant pathogens and resistance mechanisms · Powdery Mildew Fungal Diseases · Plant Parasitism and Resistance
“Eat healthy fats!” is a familiar mantra to promote a balanced diet. It highlights the human need for polyunsaturated fatty acids such as omega-3. Biochemically, omega-3 describes a double bond within a fatty acid chain 3 atoms away from the terminal methyl group. A common example of an omega-3 fatty acid is α-linolenic acid, a carboxylic acid with an 18-carbon chain and 3 cis double bonds (18:3). Essential fats such as α-linolenic acid are vital for human metabolism, and we typically obtain these lipids from plant-based foods like nuts, seeds, and oils.
Interestingly, in this respect, humans have something in common with powdery mildew fungi. These notorious plant parasites are named after the eponymous symptoms they cause on the surface of above-ground plant parts. As obligate biotrophs, powdery mildew fungi rely on living host cells for nutrients, which they extract using specialized feeding structures called haustoria. These structures form 1 day after infection and resemble tiny, inflated laboratory gloves within host epidermal cells. By 3 to 7 days post infection, the fungi produce spores on the plant surface that create the characteristic powdery appearance (Kuhn et al. 2016). The spores are filled with plant-derived lipid bodies that provide energy reservoirs for future infections (Jiang et al. 2017).
In new work, Hang Xue, Johan Jaenisch, and colleagues (Xue et al. 2025) unmasked how powdery mildew fungi manipulate host plant metabolism to extract thylakoid lipids for their own benefit. Using the Arabidopsis thaliana-Golovinomyces orontii pathosystem, the authors investigated the accumulation of lipids in infected rosette leaves. They observed a 3.5-fold increase in triacylglycerols (TAGs) in infected wild-type A. thaliana plants 12 days post inoculation, with a notable enrichment of unsaturated fatty acids, including 18:2 and 18:3. The authors then searched for 16:3 fatty acids, which are specific to the chloroplast. TAGs containing these fatty acids were indeed enriched in infected leaves, suggesting chloroplasts as the primary lipid source for powdery mildew fungi.
TAGs are storage lipids composed of 3 fatty acids attached to a central glycerol backbone. In eukaryotes, the final and rate-limiting step of TAG synthesis is catalyzed by diacylglycerol transferases (DGAT) in the endoplasmic reticulum (Xu et al. 2020). The authors tested whether loss-of-function of the canonical A. thaliana DGATs, DGAT1 and DGAT2, hinders powdery mildew spore production. Strikingly, after infection with G. orontii, dgat2 mutants showed no significant difference in spore accumulation, while dgat1 mutants showed enhanced powdery mildew spore development compared with Col-0 wild-type plants (see Figure).
Unlike DGAT1 and DGAT2, A. thaliana DGAT3 has an N-terminal chloroplast transport signal. The authors confirmed chloroplast localization using fluorophore-labeled DGAT3 in Nicotiana benthamiana and found that DGAT3 knockdown or knockout reduced powdery mildew spore production by 20% to 50%. Notably, DGAT3 prefers 18:2 and 18:3 substrates (Hernández et al. 2012), which were considerably reduced in chloroplast TAGs of infected dgat3 mutants compared with infected wild-type leaves.
Plants deprived of DGAT3 did not show constitutive activation of plant defense. Thus, the authors concluded that reduced spore production on dgat3 mutants is due to decreased TAG production in chloroplasts. By contrast, DGAT1 reduces spore production by limiting substrates for chloroplast TAG synthesis (see Figure) and/or by contributing to the presence of endoplasmic reticulum-derived lipid droplets, containing defense compounds.
In this work, the authors uncovered the role of DGAT3 in obligate biotrophy and lipid metabolism. Among other findings, their results open the interesting perspective of leveraging powdery mildew infection to study plant lipid metabolism. This approach may also support engineering plant TAG profiles enriched in fatty acids essential for a balanced human diet.
Recent related articles in The Plant Cell
Dong et al. (2025) used single-cell mass spectrometry and gene editing to produce an automated pipeline to engineer and profile increased lipid production in plant cells and whole plants. Huebbers et al. (2024) uncovered isoform-specific interactions between MILDEW RESISTANCE LOCUS O (MLO) proteins and exocyst component EXO70 proteins, affecting both modulation of trichome cell wall biogenesis and powdery mildew susceptibility. Dodds et al. (2024) reviewed the main principles of plant immunity, emphasizing key scientific milestones. Ostermeier et al. (2024) reviewed thylakoid membrane architectures in phototrophs, aiming to define the underlying principles guiding the evolution of these bioenergetic membranes.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Dodds PN, Chen J, Outram MA. Pathogen perception and signaling in plant immunity. Plant Cell. 2024:36(5):1465–1481. 10.1093/plcell/koae 02038262477 PMC 11062475 · doi ↗ · pubmed ↗
- 2Dong J, Croslow SW, Lane ST, Castro DC, Blanford J, Zhou S, Park K, Burgess S, Root M, Cahoon E, et al Enhancing lipid production in plant cells through automated high-throughput genome editing and phenotyping. Plant Cell. 2025; 10.1093/plcell/koaf 026/PMC 1185030139899469 · doi ↗ · pubmed ↗
- 3Hernández ML, Whitehead L, He Z, Gazda V, Gilday A, Kozhevnikova E, Vaistij FE, Larson TR, Graham IA. A cytosolic acyltransferase contributes to triacylglycerol synthesis in sucrose-rescued Arabidopsis seed oil catabolism mutants. Plant Physiol. 2012:160(1):215–225. 10.1104/pp.112.20154122760209 PMC 3440200 · doi ↗ · pubmed ↗
- 4Huebbers JW, Caldarescu GA, Kubátová Z, Sabol P, Levecque SCJ, Kuhn H, Kulich I, Reinstädler A, Büttgen K, Manga-Robles A, et al Interplay of EXO 70 and MLO proteins modulates trichome cell wall composition and susceptibility to powdery mildew. Plant Cell. 2024:36(4):1007–1035. 10.1093/plcell/ko ad 31938124479 PMC 10980356 · doi ↗ · pubmed ↗
- 5Jiang Y, Wang W, Xie Q, Liu N, Liu L, Wang D, Zhang X, Yang C, Chen X, Tang D, et al Plants transfer lipids to sustain colonization by mutualistic mycorrhizal and parasitic fungi. Science. 2017:356(6343):1172–1175. 10.1126/science.aam 997028596307 · doi ↗ · pubmed ↗
- 6Kuhn H, Kwaaitaal M, Kusch S, Acevedo-Garcia J, Wu H, Panstruga R. Biotrophy at its best: novel findings and unsolved mysteries of the Arabidopsis-powdery mildew pathosystem. Arabidopsis Book. 2016:14:e 0184. 10.1199/tab.018427489521 PMC 4957506 · doi ↗ · pubmed ↗
- 7Ostermeier M, Garibay-Hernández A, Holzer VJC, Schroda M, Nickelsen J. Structure, biogenesis, and evolution of thylakoid membranes. Plant Cell. 2024:36(10):4014–4035. 10.1093/plcell/koae 10238567528 PMC 11448915 · doi ↗ · pubmed ↗
- 8Xu C, Fan J, Shanklin J. Metabolic and functional connections between cytoplasmic and chloroplast triacylglycerol storage. Prog Lipid Res. 2020:80:101069. 10.1016/j.plipres.2020.10106933127353 · doi ↗ · pubmed ↗