Disruption of the EmrAB-TolC efflux pump of Escherichia coli induces global metabolic changes in multiple growth conditions
Dana E. Harmon, Klay Adamson, Joseph Wiersma
TL;DR
Deleting a bacterial efflux pump in E. coli changes multiple metabolic pathways under different growth conditions.
Contribution
Shows that the EmrAB-TolC efflux pump affects metabolic homeostasis in E. coli across various conditions.
Findings
Deleting EmrB impacts TCA cycle, carbohydrate, and amino acid metabolism.
Metabolic changes occur in both standard and nutrient stress conditions.
EmrAB-TolC helps adapt metabolism to cellular needs.
Abstract
Multidrug efflux pumps are transporters that are important for the removal of exogenous toxic molecules in bacteria. Recently, efflux pumps have been implicated in the regulation of metabolic homeostasis in Escherichia coli . Here, we investigated the contribution of EmrAB-TolC to metabolism in various conditions. Deletion of EmrB led to changes in several metabolic pathways, both in standard growth conditions and during nutrient stress. The pathways impacted include the tricarboxylic acid (TCA) cycle, and carbohydrate and amino acid metabolism. Our findings suggest that EmrAB-TolC contributes to maintaining metabolic homeostasis and adapts metabolism based on cellular needs.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
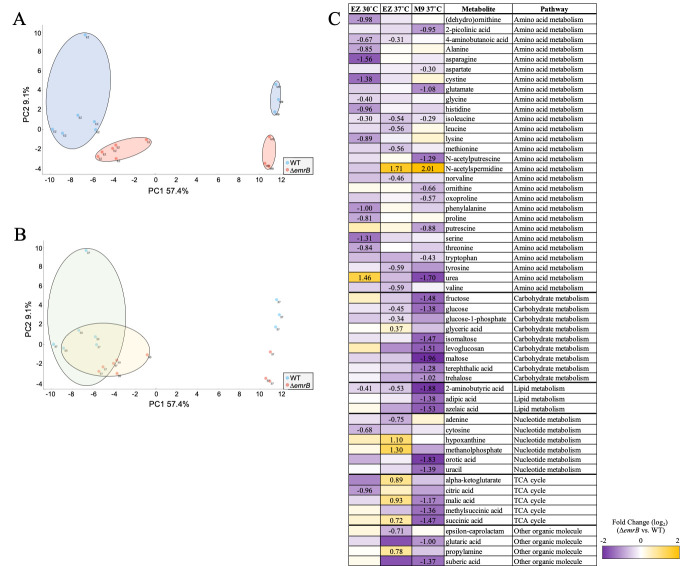 Figure 1
Figure 1|
|
|
|
|
BW25113 |
(parental) F-λ-Δ
|
CGSC, Keio collection (Baba 2006) |
|
DH8003 |
BW25113 ∆
|
This study |
- —American Association of University Women (United States)https://ror.org/05hrczs03
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNuclear reactor physics and engineering · Nuclear Materials and Properties · Particle accelerators and beam dynamics
Description
Efflux pumps are transporters found in all domains of life. Those found in bacteria are important for the extrusion of toxic molecules, such as bile salts, dyes, and various classes of antibiotics (Li et al., 2015). Because these pumps contribute to antibiotic resistance, they are often referred to as multidrug efflux pumps, referencing their well-characterized role in reducing the susceptibility of bacteria to antibiotics (Du et al., 2018; Li et al., 2015). The main and most well characterized multidrug efflux pump of E. coli , AcrAB-TolC, has been implicated in control of physiological processes such as metabolism, virulence, signaling, stress responses and motility (Cauilan et al., 2019; Harmon & Ruiz, 2022; Maldonado et al., 2023; Nishino et al., 2006; Padilla et al., 2010; Ruiz & Levy, 2014; Webber et al., 2009). Multidrug efflux pumps also exhibit extensive “functional redundancy” with different efflux pumps displaying overlapping substrate profiles, regulation, and possible roles in regulating homeostasis (Goetz et al., 2022; Teelucksingh et al., 2020; Teelucksingh et al., 2022) These observations suggest that other efflux pumps separate from AcrAB-TolC, may also play a role in the efflux of endogenous metabolites and contribute to maintenance of metabolic homeostasis in E. coli . However, relatively little is known about the endogenous functions of the other TolC-dependent efflux pumps. To address this gap in knowledge, we investigated the role of EmrAB-TolC in regulating the accumulation of metabolites.
The EmrAB-TolC efflux pump is a member of the major facilitator superfamily of efflux pumps found widely distributed in Gram-negative bacteria (Teelucksingh et al., 2020). This pump consists of three components: the substrate specificity subunit EmrB, which transports molecules across the inner membrane, the periplasmic adaptor protein EmrA, and TolC which acts as an outer membrane channel for EmrAB and a number of other efflux pumps found in E. coli (Teelucksingh et al., 2020; Yousefian et al., 2021). EmrAB-TolC has been implicated in resistance to several toxic molecules including dyes, antibiotics, bile salts, detergents, and proton motive force uncouplers (Furukawa et al., 1993; Lomovskaya & Lewis, 1992; Sulavik et al., 2001; Thanassi et al., 1997), as well as in the efflux of endogenously produced free fatty acids (Lennen et al., 2013).
Given that EmrAB-TolC has been previously demonstrated to be involved in the extrusion of free fatty acids (Lennen et al., 2013), we reasoned that this pump may also recognize and extrude other metabolites and/or contribute to global metabolism. Therefore, we used untargeted metabolomics to uncover metabolic changes that occur in a mutant of E. coli deleted for the substrate-binding subunit emrB (∆ emrB ) and compared it to the wild-type (WT) strain. We compared total metabolite levels in the extracts of cells grown in standard conditions (EZ rich media, 37˚C), as well as temperature-stress (EZ rich media, 30˚C), and nutrient-stress (M9 minimal media, 37˚C) growth conditions, to determine if the addition of temperature or nutrient stress revealed a specialized role for EmrAB-TolC in metabolism for these specific metabolic states.
The total levels of endogenous metabolites for both WT and ∆ emrB mutant E. coli were measured in the three media conditions, and the results compared using a primary component analysis ( Figure 1A ) to determine if the differences we observed in the metabolomics data were due to strain, media, or temperature differences. As anticipated, we found that media composition had a substantial impact on global metabolism, with cells grown in EZ rich media, having a distinct metabolome from cells grown in M9 minimal salts. We also observed that independent clustering of the WT and emrB -deleted strains suggests that the EmrAB-TolC efflux pump also exerts an influence on metabolism. Interestingly, we found a greater degree of overlap between the metabolic changes that occur between 30 and 37˚C in cells grown in rich media, suggesting a far smaller metabolic effect caused by temperature ( Figure 1B ).
Comparing the accumulation of metabolites between the WT and ∆ emrB mutant, we found that the ∆ emrB mutant had significant differences in the accumulation of several metabolites ( Figure 1C ). Notably, these metabolites varied in the different conditions, suggesting that the contribution of the pump changes in different metabolic circumstances. For example, in rich media at 37˚C, we found the statistically significant ( p ≤ 0.05) accumulation of three TCA cycle intermediates, including alpha-ketoglutarate (1.85-fold), succinic acid (1.65-fold), and malic acid (1.90-fold) in cell extracts. These findings suggest that the efflux activity of EmrAB-TolC, either directly or indirectly, controls the levels of metabolites in this critical central metabolic pathway in standard, non-stressed conditions. These findings are consistent with observations made for AcrAB-TolC, where it was shown that the TCA cycle intermediates fumaric acid and malic acid both accumulate in *E. coli * deleted for *acrB * (Cauilan et al., 2019).
In both rich and minimal media culture at 37˚C, we also observed accumulation of N8-spermidine in the ∆ emrB strain (3.3-fold and 4.0-fold, respectively). Previous reports have shown that the enzyme that produces this metabolite, spermidine- N- acetyltransferase, is more active in nutrient poor conditions (Fukuchi et al., 1994). Consistent with this, we observed a strong increase in total amounts of N8-spermidine produced by WT in EZ versus M9 (3.7-fold), as well as by the ∆ emrB mutant (4.6-fold) *. * As to why we observe an overaccumulation in the ∆ emrB strain versus the WT, remains unclear, but it has been shown that increased production of monoacetylated spermidine occurs when cells experience a variety of chemical and physical stresses (Carper et al., 1991). It is possible that the loss of EmrAB-TolC efflux activity leads a general stress response that induces the production of N8-spermidine in cells deleted for emrB or, more tantalizingly, acetylated spermidine may be a substrate of the EmrAB-TolC pump .
Here we present the findings of a metabolomics study, in which we investigated the contribution of the EmrAB-TolC efflux pump to metabolism in E. coli in standard, low-temperature, and nutrient-limited conditions. The authors of this study recognize that the complexity of biological systems requires the careful interpretation of metabolomic analysis findings. Therefore, any results from this study that suggest a role for EmrAB-TolC in the control of a specific metabolic pathway require further investigation to confirm these observations. Nevertheless, the global changes observed upon disruption of EmrAB-TolC provides compelling evidence for its ability to shape metabolite pools, and highlights the potential role of EmrAB-TolC as a regulator of metabolic homeostasis in both standard and nutrient limited conditions. This is the first metabolomics study investigating the EmrAB-TolC efflux pump and is the first in E. coli to examine how a single efflux pump regulates metabolism in multiple growth conditions. Our findings suggest that EmrAB-TolC is involved in shaping intracellular metabolite profiles, both in standard growth conditions and during nutrient limitation. This could be attributed to the ability of EmrAB-TolC to efflux substrates and/or intermediates from several metabolic pathways, including the TCA cycle, carbohydrate metabolism, and amino acid biosynthesis and degradation, with EmrAB-TolC effectively acting as a regulator of cellular metabolism. Alternatively, the metabolic changes we observed in this study may not be solely due to the direct loss of EmrAB-TolC efflux activity. Studies of AcrAB-TolC have shown that the periplasmic adapter protein AcrA can interact with two other efflux pumps, AcrD and AcrF, in addition to AcrB (Elkins & Nikaido, 2002, 2003). Therefore, in the absence of EmrB, it is possible that EmrA may have alternate associations with other pumps, which could lead to secondary effects on metabolism. While experimentally evaluating these possibilities goes beyond the scope of this study, these questions are important topics for follow up studies to further understand the complex interplay between the TolC-dependent efflux pumps and their impact on global metabolism. Other metabolomics studies, in addition to this one, suggest a broad role for efflux pumps as regulators of metabolism (Cauilan et al., 2019; Wang-Kan et al., 2021). The insights provided by this study of the EmrAB-TolC efflux pump provides further evidence for the significant impact the TolC-dependent efflux pumps play in the ability of *E. coli * to adapt to diverse environments.
Methods
Strains
The DH8003 was generated from JW2661 (Keio collection; (Baba et al., 2006)) by removal of the *kan * cassette as described previously (Datsenko & Wanner, 2000).
Untargeted metabolomics
Three biological replicates of the parental and ∆ emrB mutants were cultured with aeration in either EZ rich defined medium supplemented with 0.2% glucose (Teknova) at either 37˚C or 30˚C, or in M9 minimal salts (MP Biomedicals) supplemented with 0.2% glucose and 1 mM MgSO 4 , at 37˚C to mid-exponential (OD 600 of 0.35-0.4). Two milliliters of each culture were flash frozen in liquid nitrogen and shipped on dry ice to the West Coast Metabolomics Center (WCMC) at UC Davis for analysis by untargeted metabolomics. The cell extracts were prepared using the Matyash extraction procedure (Matyash et al., 2008), derivatized with MSTFA/FAMEs, and analyzed using the WCMC ALEX-CIS GCTOF MS (automated liner exchange-cold injection system-gas chromatography time of flight-mass spectrometry) platform. Quantification of each metabolite was reported by WCMC as peak height, following the established protocol within the BinBase (rtx5) algorithm established by the core facility. We further normalized the peak heights for all metabolites to the optical density (600 nm) measurements for the individual samples from which the peaks were derived. Finally, we determined the average peak height for each metabolite and present the data as the log 2 -fold difference between the ∆ emrB mutant and the WT. To determine statistical significance, we used the student’s t-test (two independent samples with equal variance and a two tailed distribution) in Microsoft Excel 365 to compare the abundance of a given metabolite in the ∆ *emrB * to that found in the WT. Those that had a p- value ≤ 0.05 were considered significant.
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Baba T Ara T Hasegawa M Takai Y Okumura Y Baba M Datsenko KA Tomita M Wanner BL Mori H 2006221 Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: the Keio collection.Mol Syst Biol 22006.00082006.000810.1038/msb 410005016738554 PMC 1681482 · doi ↗ · pubmed ↗
- 2Carper SW Willis DG Manning KA Gerner EW 199175 Spermidine acetylation in response to a variety of stresses in Escherichia coli.J Biol Chem 266190021-925812439124412061318 · pubmed ↗
- 3Cauilan A Ramos K Harmon DE Ruiz C 2019517 Global effect of the Acr AB-Tol C multidrug efflux pump of Escherichia coli in cell metabolism revealed by untargeted metabolomics.Int J Antimicrob Agents 5410924-857910510710.1016/j.ijantimicag.2019.05.01531108224 · doi ↗ · pubmed ↗
- 4Datsenko KA Wanner BL 200066 One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products.Proc Natl Acad Sci U S A 97120027-84246640664510.1073/pnas.12016329710829079 PMC 18686 · doi ↗ · pubmed ↗
- 5Du D Wang-Kan X Neuberger A van Veen HW Pos KM Piddock LJV Luisi BF 201891 Multidrug efflux pumps: structure, function and regulation.Nat Rev Microbiol 1691740-152652353910.1038/s 41579-018-0048-630002505 · doi ↗ · pubmed ↗
- 6Elkins CA Nikaido H 2002121 Substrate specificity of the RND-type multidrug efflux pumps Acr B and Acr D of Escherichia coli is determined predominantly by two large periplasmic loops.J Bacteriol 184230021-91936490649810.1128/JB.184.23.6490-6499.200212426336 PMC 135441 · doi ↗ · pubmed ↗
- 7Elkins CA Nikaido H 200391 Chimeric analysis of Acr A function reveals the importance of its C-terminal domain in its interaction with the Acr B multidrug efflux pump.J Bacteriol 185180021-91935349535610.1128/JB.185.18.5349-5356.200312949086 PMC 193755 · doi ↗ · pubmed ↗
- 8Fukuchi J Kashiwagi K Takio K Igarashi K 199499 Properties and structure of spermidine acetyltransferase in Escherichia coli.J Biol Chem 269360021-925822581225858077207 · pubmed ↗