Unveiling new species of Phragmidiaceae (Basidiomycota, Pucciniales) on rosaceous plants from Guizhou, China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
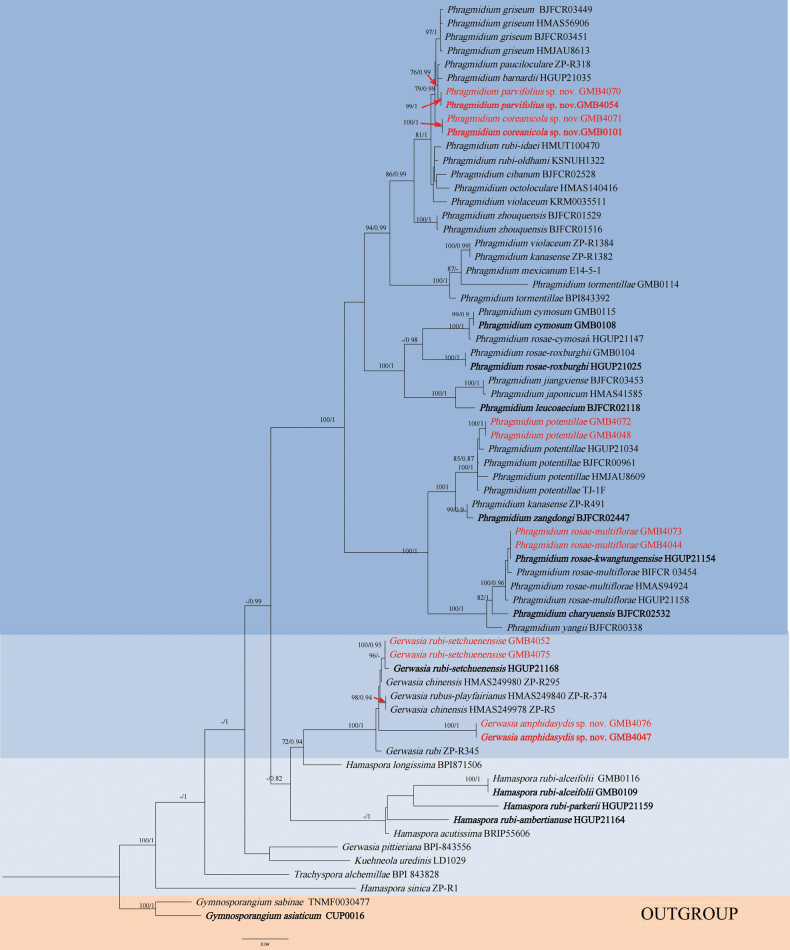 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5| Species | Specimen No. | Host | Province, Country | GenBank Accession No. | References | |
|---|---|---|---|---|---|---|
|
|
| |||||
|
| HMAS249978 (ZP-R5) |
| Yunnan, China |
|
|
|
|
| HMAS249980 (ZP-R295) |
| Yunnan, China |
|
|
|
|
| BPI 843556 | USA |
|
|
| |
|
| (ZP-R345) |
| Sichuan, China | – |
|
|
|
| GMB4047* |
| Guizhou, China |
|
| This study |
|
| GMB4076 |
| Guizhou, China |
|
| This study |
|
| GMB4052 |
| Guizhou, China |
|
| This study |
|
| GMB4075 |
| Guizhou, China |
|
| This study |
|
| HGUP21168* |
| Guizhou, China |
|
|
|
|
| HMAS249840 (ZP-R 1374) |
| Guangxi, China |
|
|
|
|
| CUP-0016* |
| Japan |
|
|
|
|
| TNM F0030477 |
| Bulgaria: Sofia |
|
|
|
|
| BRIP:55606 |
| Philippines | – |
|
|
|
| ZP-R1 |
| Guangdong, China |
|
|
|
|
| BPI 871506 |
| South Africa | – |
|
|
|
| HGUP21164* |
| Guizhou, China |
|
|
|
|
| HGUP21159* |
| Guizhou, China |
|
|
|
|
| GMB0109* |
| Guizhou, China |
|
|
|
|
| GMB0116 |
| Guizhou, China |
|
|
|
|
| LD1029 | New York, America |
|
|
| |
|
| HGUP21035 |
| Guizhou, China |
|
|
|
|
| BJFC: |
| China |
|
|
|
|
| BJFC: |
| China |
|
|
|
|
| GMB0115 |
| Guizhou, China |
|
|
|
|
| GMB0108* |
| Guizhou, China |
|
|
|
|
| GMB0101* |
| Guizhou, China |
|
| This study |
|
| GMB4071 |
| Guizhou, China |
|
| This study |
|
| HMAS56906 |
| Beijing, China |
|
|
|
|
| BJFCR 03451 | – | Beijing, China |
|
|
|
|
| BJFCR03449 |
| Beijing, China |
|
|
|
|
| HMAS41585 |
| Fujian, China |
|
|
|
|
| BJFCR 03453* |
| Jiangxi, China |
|
|
|
|
| ZP-R1382 |
| Yunnan, China |
|
|
|
|
| ZP-R491 |
| Xinjiang, China | – |
|
|
|
| BJFCR 02118* | Yunnan, China |
|
|
| |
|
| E14_5_1 |
| Slovenia, Forestry |
|
|
|
|
| HMAS140416 |
| China |
|
|
|
|
| ZP-R318 |
| Guangxi, China |
|
|
|
|
| HGUP21034 |
| Tasmania, Australia |
|
|
|
|
| HMJAU8609 |
| China |
|
|
|
|
| BJFCR 00961 |
| Beijing, China |
|
|
|
|
| TJ-1F |
| China |
|
|
|
|
| GMB4048 |
| Guizhou, China |
|
| This study |
|
| GMB4072 | Guizhou, China |
|
| This study | |
|
| HGUP21158 |
| Guizhou, China |
|
|
|
|
| BIFCR 03454 |
| Jiangxi, China |
|
|
|
|
| HMAS94924 |
| Zhejiang, China | – |
|
|
|
| GMB4044 |
| Guizhou, China |
|
| This study |
|
| GMB4073 |
| Guizhou, China |
|
| This study |
|
| HGUP21025* |
| Guizhou, China |
|
|
|
|
| GMB0104 |
| Guizhou, China |
| – |
|
|
| HGUP21154* |
| Guizhou, China |
| – |
|
|
| HGUP21147 |
| Guizhou, China |
|
|
|
|
| HMUT100470 |
| Chongqing, China |
|
| – |
|
| KSNUH1322 |
| South Korea |
|
|
|
| HMJAU8613 | – | Changchun, China |
|
|
| |
|
| GMB4054* |
| Guizhou, China |
|
| This study |
|
| GMB4070 |
| Guizhou, China |
|
| This study |
|
|
| Guizhou, China |
| – |
| |
|
| BPI 843392 | USA: Maryland | – |
|
| |
|
| KRM 0035511 | Germany |
|
|
| |
|
| ZP-R1384 |
| Yunnan, China |
|
|
|
|
| BJFCR 00338 |
| Beijing, China |
|
|
|
|
| BJFC: |
| Tibet, China |
|
|
|
|
| BJFCR01516 |
| Yunnan, China |
|
|
|
|
| BJFCR01529 |
| Yunnan, China |
|
|
|
|
| BPI 843828 |
| Switzerland |
|
|
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Yeasts and Rust Fungi Studies · Plant Pathogens and Fungal Diseases
Introduction
Pucciniales (Basidiomycota, Pucciniomycetes) represents approximately 25% of basidiomycete fungi and constitutes one of the most prevalent fungal groups, parasitizing leaves, fruits, and branches of plants, thereby inhibiting normal growth and development of plants and impacting yield and quality (Aime and McTaggart 2021; Zhao et al. 2022a). To date, seven suborders (i.e., Araucariomycetineae, Melampsorineae, Mikronegeriineae, Raveneliineae, Rogerpetersoniineae, Skierkineae, and Uredinineae) and 18 families (i.e., Araucariomycetaceae, Coleosporiaceae, Crossopsoraceae, Gymnosporangiaceae, Melampsoraceae, Milesinaceae, Ochropsoraceae, Phakopsoraceae, Phragmidiaceae, Pileolariaceae, Pucciniaceae, Pucciniastraceae, Raveneliaceae, Rogerpetersoniaceae, Skierkaceae, Sphaerophragmiaceae, Tranzscheliaceae, and Zaghouaniaceae) have been reported (Aime et al. 2017; Aime and McTaggart 2021; Zhao et al. 2022b).
According to estimations, there are approximately 35,000 higher plant species in China, which are categorized into 454 families and 3,818 genera (Wang et al. 2015). It is further estimated that between 1,700 and 8,800 species of rust fungi may exist within the country, posing a significant threat to the health and productivity of these plant species (Zhao et al. 2021). More than 8,400 rust taxa have been documented worldwide (Zhao et al. 2021; Sun et al. 2022). Based on their morphological characteristics and host associations, 1,200 species from 71 genera across 15 families have been documented in China. Among them, more than 70 species of Phragmidium have been identified. (Dai 1979; Zhuang et al. 1998, 2003, 2005, 2012, 2021; Ji et al. 2019, 2020; Zhao et al. 2021; Sun et al. 2022, 2024). Plants are susceptible to Coleosporiaceae, Melampsoraceae, Phragmidiaceae, and Pucciniaceae, which are the dominant families of rust taxa in China (Zhao et al. 2021; Zhao et al. 2022b; Sun et al. 2024).
The genus Phragmidium (Pucciniales: Phragmidiaceae) was established by Link (1816). Phragmidiaceae comprises a diverse array of species, encompassing 14 genera and an estimated 200 species, primarily targeting the economically vital family Rosaceae (examples of hosts in Rosaceae: Rosa, Rubus, Potentilla, Sanguisorba, Duchesnea, and Acaena) (Zhao et al. 2021; Sun et al. 2024). A total of 115 Phragmidium species have been described (Cummins 1931; Arthur 1934; Wahyuno et al. 2001; Cummins and Hiratsuka 2003; Zhuang et al. 2012; Yang et al. 2015; Ali et al. 2017; Zhao et al. 2021; Sun et al. 2022; Ya 2024). Among these, 38 species specifically target and parasitize members of the genus Rosa (Ya 2024), and at least 39 Phragmidium species have been reported in China (Wei 1988; Zhuang et al. 2003, 2012; Yang et al. 2015; Liu et al. 2018).
The susceptibility of Rosaceae plants to Phragmidiaceae infections underscores the importance of understanding the biology, ecology, and management strategies of these fungal pathogens (Dietel 1905; Yun et al. 2011; Sun et al. 2022). During the field investigation of rust fungi on medicinal plants in Guizhou Province, China, three potential new species were found belonging to the genera Gerwasia and Phragmidium of Phragmidiaceae, which infect the members of Rosaceae, as well as one species that infects a new host. We conducted a phylogenetic analysis using multi-locus (ITS and LSU) phylogeny and morphological characteristics to better understand their taxonomic position. Descriptions, illustrations, and phylogenetic analysis results of all the novel species and the new host record are provided.
Materials and methods
Sample collection and preservation
Rust-infected specimens were collected in Guizhou Province, China, from August to November of each year from 2021 to 2023. All host and habitat information of specimens was recorded (Rathnayaka et al. 2024). Photographs of the infected plants were taken using a camera (Canon G15, Corporation, Tokyo, Japan). The samples were kept in blotting papers and were brought to the laboratory for examination. The collected specimens were partly kept in a refrigerator at 4 °C for spare parts and partly pressed and air-dried to make pressed specimens (Wei and Wang 2011; Wu et al. 2023). All specimens were deposited at the herbarium of Guizhou Medical University (GMB) and Kunming Institute of Botany, Chinese Academy of Sciences (KUN-HKAS).
Morphological characterization
Macroscopic characteristics were observed under a stereomicroscope (Olympus SZ61), and photographs were taken with a digital camera (Canon 700D) fitted with a light microscope (Nikon Ni). The infected portions were examined, and photographs were taken as described by Wu et al. (2023). More than 30 measurements were noted for each type of teliospores, urediniospores, and paraphyses for each sample using the Tarosoft (R) Image Frame Work v. 0.9.0.7. The images were arranged using Adobe Photoshop CS6 (Adobe Systems, the USA).
DNA extraction and PCR amplification
The infected portion of the rust fungus was scraped using a sterilized scalpel. DNA extraction was carried out following the manufacturer’s protocols for the Biomiga Fungal gDNA Kit. The DNA samples were kept at –20 °C. The rust-specific primer pairs Rust2inv (Aime 2006) and ITS4rust (Pfunder et al. 2001) were used for the PCR amplification of the regions of internal transcribed spacer (ITS), whereas the universal primer pairs LR0R (Hopple and Vilgalys 1999) and LR6 (Vilgalys and Hester 1990) were used for the large subunit ribosomal (LSU). The composition of a 25 μL PCR mixture comprised the following: 9.5 μL of double-distilled water, 12.5 μL of PCR Master Mix, 1 μL of each primer, as well as 1 μL of template DNA. The qualified PCR products were verified through 1.5% agarose gel electrophoresis, stained with GoldenView, and subsequently submitted to Sangon Co. China for sequencing (Zhao et al. 2016).
Phylogenetic analyses
All sequences were obtained in ABI file format and deposited in the GenBank (Table 1). The consensus sequences were blasted in GenBank using the BLAST algorithm. The similar sequences were retrieved from the GenBank database. The molecular phylogeny was inferred from a combined dataset of ITS and LSU sequences. The reference sequences retrieved from open databases originated from recently published literature (Zhao et al. 2021; Wu et al. 2023; Sun et al. 2024). All the ambiguous nucleotides were trimmed using BioEdit software v.7.0.5.3 and TrimAL (Hall 1999). Sequences were aligned using the MAFFT v.7.110 online tool (Katoh and Standley 2013). The alignments are available in TreeBASE (www.treebase.org/treebase-web/home.html) under ID31754 for LSU and ITS rDNA sequences. The maximum likelihood (ML) analysis was implemented in RAxML v.8.2.12 using the GTRGAMMA substitution model with 1,000 bootstrap replicates (Stamatakis 2015). The phylogenetic analyses were also performed using Bayesian inference in MrBayes v.3.2.1 (Ronquist et al. 2015) online. The Markov chain Monte Carlo (MCMC) sampling in MrBayes v.3.2.2 (Ronquist et al. 2015) was used to determine the Bayesian posterior probabilities (BYPP). Six simultaneous Markov chains were run for 3,000,000 generations, and trees were sampled every 1,000^th^ generation. All analyses were run on the CIPRES Science Gateway v.3.3 web portal (Miller et al. 2010; Huelsenbeck and Ronquist 2001). The phylogenetic tree was visualized by FigTree v.1.4.3 (Rambaut 2012).
Results
Phylogeny
In this study, 14 samples from twelve host plant species were collected in Guizhou Province. Through morphological and molecular systematic studies, a total of 6 species were identified, including three new species, one new host species, and two known species.
For the final phylogenetic analyses, taxa were selected based on their morphological and phylogenetic affinities, largely following the approach of Zhao et al. (2021). Both the RAxML and BYPP analyses produced similar overall tree topologies with no significant differences. The alignment includes 67 species (comprising one family: Phragmidiaceae, six genera: Phragmidium, Gerwasia, Hamaspora, Kuehneola, Trachyspora, and the outgroup genus Gymnosporangium) and contains 1245 characters, including gaps (ITS: 398 bp, LSU: 847 bp). Among these, three novel species have been identified (Gerwasiaamphidasydis sp. nov., Phragmidiumparvifolius sp. nov., and P.coreanicola sp. nov.). Additionally, one species (G.rubi-setchuenensise) was reported for the first time on Rubusbuergeri.
Taxonomy
Family Phragmidiaceae Corda, Icon. Fung. (Prague) 1: 6. 1837
Genus Gerwasia Racib. Bull. Acad. Sci. Lett. Cracovie, Cl. Sci. Math. Nat. Sér. B, Sci. Nat. 3: 270. 1909
Gerwasia
amphidasydis
Taxon classificationFungiPuccinialesPhragmidiaceae
Q. F. Zhang, Q. Z. Wu & Q. R. Li sp. nov.
72625E3E-7BB9-51D7-8BF5-55B5325D1A33
854995
Type.
China • Guizhou Province, Zunyi City, Kuangkuoshui Nature Reserve (28°12'40"N, 107°10'22"E), 2227 m a.s.l., on leaves of Rubusamphidasys. 1 November 2022, Q. Z. Wu and Q. F. Zhang (holotype GMB4047, isotype KUN-HKAS144247);
Etymology.
The epithet refers to the host species, Rubusamphidasys Focke ex Diels., from which the holotype was collected.
Description.
Spermogonia, Aecia, and Telia not found. Uredinia 0.2–0.8 mm diam. produced on the abaxial leaf surface, scattered to gregarious, hypophyllous, covered by peridium, small, rounded, light yellow, or orange-yellow. Urediniospores 29–41 × 22–29 μm (av. = 34 × 26 μm, n = 30), globose to subglobose or ovoid, golden, yellow-brown, wall 1.2–2.5 µm thick at sides, hyaline, prominent sparsely echinulate, markings elongated longitudinally, 1.3–3.1 µm in distance, pore obscure, germ pores inconspicuous. Pedicel broken; paraphyses not seen.
Additional material examined.
China • Guizhou Province, Zunyi City, Kuangkuoshui Nature Reserve, (28°12'38"N, 107°10'21"E) 2214 m a.s.l., on the leaves of Rubusamphidasys (Rosaceae). 1 November 2022, Q. Z. Wu and Q. F. Zhang (GMB4076).
Notes.
Gerwasiaamphidasydis was the first species of Gerwasia described on Rubusamphidasys. Our phylogenetic analyses showed that G.amphidasydis formed a separate branch (Fig. 1). Morphologically, G.amphidasydis and G.rubi exhibit similar spines. Moreover, the difference between G.amphidasydis and G.rubi is that the former has bigger urediniospores (29–41 × 22–29 μm vs. 22–33 × 16–26 µm) (Ito 1950; Hiratsuka et al. 1992). Gerwasiaamphidasydis and G.guanganensis have similar uredinia and urediniospores; however, G.guanganensis has longer spine distances compared to G.amphidasydis (4.0–6.0 µm vs. 1.3–3.1 µm) (Zhao et al. 2021). Gerwasiaamphidasydis is distinguishable from G.rubi-setchuenensise by having larger urediniospores (29–41 × 22–29 μm vs. 18–29 × 15–22 μm) and a thinner wall (1.2–2.5 µm vs. 2.1–3.2 μm) (Sun et al. 2024).
RAxML tree of the family Phragmidiaceae based on rDNA ITS and LSU sequences. ML bootstrap supports (≥75%) and Bayesian posterior probability (≥0.90) are indicated as ML/BYPP. The tree is rooted to G.sabinae and G.asiaticum (Yang et al. 2015; Aime et al. 2018). All species newly studied are indicated in red, with novel species highlighted in bold red. Type materials were highlighted in bold.
Gerwasiaamphidasydis (Holotype GMB4047) A–D host and its habitat E–F uredinia under a stereomicroscope G–K urediniospores. Scale bars: 1 mm (D, F); 0.5 mm (E); 10 μm (G–K).
Additionally, the LSU sequences of Gerwasiaamphidasydis also differ from that of G.rubi with 93.74% similarity and from G.rubi-setchuenensis with 90.56% similarity. The ITS sequence for Gerwasiarubi is not available in the NCBI database, whereas the ITS sequence similarity between Gerwasiaamphidasydis and G.rubi-setchuenensis is 98.56%.
Genus Phragmidium Link, Mag. Gesell. Naturf. Freunde, Berlin 7: 30. 1816
Phragmidium
coreanicola
Taxon classificationFungiPuccinialesPhragmidiaceae
Q. F. Zhang, Q. Z. Wu & Q. R. Li sp. nov.
0E3E9EE1-38C7-59E6-A93F-00A48BC63344
855005
Type.
China • Guizhou Province, Guiyang City, Campus of Guizhou Medical University (26°22'48"N, 106°37'30"E), 1911 m a.s.l., on leaves of Rubuscoreanus (Rosaceae), 7 October 2021, Q. Z. Wu (holotype GMB0101, isotype KUN-HKAS144249).
Etymology.
The epithet refers to the host species, RubuscoreanusMiq.var.coreanus, from which the holotype was collected.
Description.
Spermogonia and Aecia not found. Uredinia 0.1–0.7 mm diam., produced on the abaxial leaf surface, scattered to gregarious, hypophyllous, yellow spots, scattered, irregular patches. Urediniospores 20–29 × 14–25 μm (av. = 24 × 21 μm, n = 30), globose to subglobose or broadly elliptical to ellipsoidal, wall 0.8–2.1 μm thick (av. = 1.4 μm, n = 30), inconspicuous or smooth at the base; inclusions orange-yellow or pale-yellow; germ pores 2–3, sub-equatorial. Telia 0.1–0.9 mm diam., hypophyllous, dark brown to black, clustered or scattered, bacilliform. Teliospores 107–167 × 25–35 μm (av. = 134 × 30 μm, n = 30), cylindrical, 5–7 cells, often 6, reddish-brown to opaque, rounded at the apex, rounded or somewhat attenuate at the base, not or slightly constricted at the septum, pedicels sub-hyaline, persistent, 47–96 × 12–19 μm (av. = 68 × 16 μm, n = 30), with a swollen base that gradually shows orange-yellow contents towards the lower end. Pedicel broken; paraphyses not seen.
Phragmidiumcoreanicola (Holotype, GMB0101) A–C host and its habitat D–E uredinia and telia F, G urediniospores H–K teliospores. Scale bars: 0.5 mm (D, E); 10 μm (F–K).
Additional material examined.
China • Guizhou Province, Qingzhen City (26°34'58"N, 106°28'28"E), 1972 m a.s.l., on leaves of Rubuscoreanus (Rosaceae), 7 October 2021, Q.Z. Wu (GMB4071).
Notes.
Phragmidiumcoreanicola formed a separate branch in our phylogenetic analyses (Fig. 1). Morphologically, P.coreanicola differs from P.griseum in having slightly wider urediniospores (14–25 μm vs. 13–21 μm) and larger teliospores (107–167 × 25–35 μm vs. 50–125 × 18–28 μm) (Liu et al. 2018). Additionally, P.coreanicola is reported on Rubuscoreanus, whereas P.griseum was found on Rubuscrataegifolius. Phragmidiumcoreanicola differs from P.cibanum, which is reported on Rubusniveus, by having bigger urediniospores (20–29 × 14–25 μm vs. 17–20 × 18–19 μm) and larger teliospores (107–167 × 25–35 μm vs. 80–100 × 20–30 μm) (Wei 1988; Hiratsuka et al.1992; Liu et al. 2018). Phragmidiumcoreanicola has the same host species as P.rubi-coreani in Guiyang City. However, P.coreanicola has larger teliospores (107–167 × 25–35 μm vs. 29–74 ×14–37 µm) (Sun et al. 2022). The morphological comparison between P.coreanicola and P.pauciloculare shows that the uredinia of P.coreanicola are larger than those of P.pauciloculare (0.1–0.7 mm diam vs. 0.2–0.3 mm diam), and teliospores of P.coreanicola are also larger than those of P.pauciloculare (107–167 × 25–35 μm vs. 35–111 × 18–27 µm) (Wei 1988).
Furthermore, the ITS sequences of P.coreanicola and P.griseum exhibit significant differences, with a similarity of 87.33%. However, the LSU sequences of P.coreanicola and P.griseum have little variation, sharing a similarity of 99.38%.
Phragmidium
parvifolius
Taxon classificationFungiPuccinialesPhragmidiaceae
Q. F. Zhang, Q. Z. Wu & Q. R. Li sp. nov.
751ADE41-8F22-523A-8907-1B2CB804FA77
855006
Type.
China • Guizhou Province, Guiyang City, Huaxi District (26°43′27.3″N, 106°67′14.4″E), 1,114 m a.s.l., on leaves of Rubusparvifolius (Rosaceae), 3 November 2022, Q. Z. Wu and Q. F. Zhang (holotype GMB4054, isotype KUN-HKAS144250).
Etymology.
The epithet refers to the host species, Rubusparvifolius L., from which the holotype was collected.
Description.
Spermogonia, Aecia and Telia not found. Uredinia 0.3–0.8 mm diam., produced on the abaxial leaf surface, scattered to gregarious, hypophyllous, rounded to irregular, powdery, orange, pulverulent, at first covered by the epidermis, later, not surrounded by host epidermis; Urediniospores 18–32 × 12–24 μm (av. = 22 × 18 μm, n = 30), globose, oblong, orange, wall 1.1–1.7 μm thick (av. = 1.3 μm, n = 30) at sides, regularly echinulate with stout spines; germ pores 2–3, supra-equatorial. Paraphyses 49–83 × 10–19 μm (av. = 65 × 15 μm, n = 30), hyaline, curved.
Phragmidiumparvifolius (Holotype, GMB4054) A–D host and its habitat E, F uredinia under a stereomicroscope G, H urediniospores and paraphyses I–L urediniospores M paraphyses. Scale bars: 1 mm (D); 0.5 mm (E, F); 10 μm (G–M).
Additional material examined.
China • Guizhou Province, Guiyang City, Huaxi District (26°43′59.7″N, 106°67′66.5″E), 1114 m a.s.l., on leaves of Rubusparvifolius (Rosaceae), 3 November 2022, Q. Z. Wu and Q. F. Zhang (GMB4070).
Notes.
Phylogenetically, P.parvifolius formed a sister branch to P.barnardii Plowr. & G. Winter (HGU21035), which was also reported on Rubusparvifolius (Fig. 1). Morphologically, P.parvifolius can be easily differentiated from P.barnardii by its larger urediniospores (18–32 × 12–24 μm vs. 16–19 × 15–18 µm) and larger paraphyses (49–83 × 10–19 μm vs. 26–39 × 10–13 µm) (Winter 1886; McTaggart et al. 2016; Sun et al. 2022). In terms of urediniospore size, P.parvifolius is similar to P.griseum (Dietel) Syd. However, P.parvifolius differs from P.griseum by having relatively larger paraphyses (49–83 × 10–19 μm vs. 34–70 × 7–16 μm) and by its host, Rubusparvifolius vs. Rubuscrataegifolius (Wei 1988; Hiratsuka et al. 1992; Liu et al. 2018; Sun et al. 2024). Additionally, P.parvifolius differs from P.pauciloculare by its larger urediniospores (18–32 μm vs. 13–20 μm) (Wei 1988; Hiratsuka et al. 1992). Phragmidiumparvifolius and P.kanas have the same urediniospores and paraphyses, but P.parvifolius has no teliospores, whereas P.kanas has them (Zhao et al. 2021).
Furthermore, the morphological comparison between P.parvifolius and P.coreanicola (this study) shows that the urediniospores of P.parvifolius are larger than those of P.coreanicola (18–32 μm vs. 20–29 μm), and P.parvifolius has no teliospores, whereas P.coreanicola has them. The ITS and LSU sequence similarities of P.parvifolius with P.coreanicola are 97.57% and 99.22%.
Gerwasia
rubi-setchuenensis
Taxon classificationFungiPuccinialesPhragmidiaceae
J.E. Sun, Yong Wang bis & K.D. Hyde, (2024)
0B748040-F67A-5F99-A29D-23FAA12DCB04
Host.
Rubusbuergeri Miq.
Description.
Spermogonia, aecia, and telia unknown. Uredinia 0.4–1.0 mm diam., hypophyllous, pulverulent, golden, scattered, irregular, surrounded by host epidermis. Urediniospores 24–30 × 19–25 µm (av. = 27.2 × 22.3 µm, n = 30), subglobose or fusiform, inclusions golden or bright yellow; wall 1.4–2.9 µm thick (av. = 2.0 μm, n = 30), colorless, irregularly elongated verrucae.
Materials examined.
China • Guizhou Province, Zunyi City, Xishui County (28°49'38"N, 106°41'23"E), 1223 m a.s.l., on the leaves of Rubusbuergeri Miq. (Rosaceae), 3 November, 2022, Q. Z. Wu and Q. F. Zhang (GMB4052); China • Guizhou Province, Zunyi City, Xishui County (28°33'43"N, 106°24'3"E), 1997 m a.s.l., on Rubusbuergeri (Rosaceae), 3 November 2022, Q. Z. Wu and Q. F. Zhang (GMB4075).
Gerwasiarubi-setchuenensis (GMB4052) A–D host and its habitat E–F uredinia under the stereomicroscope G–K urediniospores. Scale bars: 0.5 mm (E–F); 10 μm (G–L).
Notes.
In the phylogram (Fig. 1), our collections (GMB4052 and GMB4075) clustered with G.rubi-setchuenensis (HGUP21168). The morphological characteristics of our specimen are consistent with the original description of G.rubi-setchuenensis, and the DNA sequence aligns with that of G.rubi-setchuenensis HGUP21168 (ITS 100%; LSU 99.67%) (Sun et al. 2024). The only difference observed between the descriptions and figure of G.rubi-setchuenensis in Sun et al. (2024) is the size of the urediniospores. The urediniospores of G.rubi-setchuenensis (GMB4075) are slightly wider than those of G.rubi-setchuenensis (HGUP21168) (19–25 μm vs. 15–22 μm). This study identifies Rubusbuergeri as a new host for this fungus.
Discussion
The exploration of rust fungi in China began in the mid-19^th^ century, and to date, over 1200 rust taxa have been documented (Zhuang et al. 1998, 2005, 2021; Zhao et al. 2022a, b; Sun et al. 2024). Molecular techniques have significantly advanced fungal species identification, but accurately identifying rust fungi remains challenging, necessitating a comprehensive approach incorporating morphology, host specificity, and phylogenetic analyses (Sun et al. 2024). In China, over 70 species of Phragmidium have been described, although numerous species remain without molecular data. According to our literature review, approximately 22 species of Phragmidium have been reported in Guizhou (Cummins 1931; Zhuang et al. 2012; Aime et al. 2018; Aime and McTaggart 2021; Zhao et al. 2021; Sun et al. 2022, 2024). These studies emphasize the critical role of integrating morphological data, host specificity, and phylogenetic insights for a comprehensive understanding and accurate identification of rust fungi.
In our investigation, three new species of Phragmidiaceae belonging to the genera Gerwasia and Phragmidium are introduced based on phylogenetic analysis of the ITS and LSU regions and morphological features. amphidasydis sp. nov., Phragmidiumcoreanicola sp. nov., and P.parvifolius sp. nov. infected Rubusamphidasys, Rubuscoreanus, and Rubusparvifolius, respectively. The host of the P.coreanicola is the same as that of P.rubi-coreani; however, P.coreanicola has larger teliospores (107–167 × 25–35 μm vs. 29–74 ×14–37 µm), and P.rubi-coreani possesses aeciospores (Sun et al. 2022). In addition, Rubusbuergeri was identified as a new host plant for Gerwasiarubi-setchuenensis. Previously, it was known only on Rubussetchuenensis (Sun et al. 2024). At the same time, we discovered some samples of Phragmidiumrosae‐multiflorae Dietel, and Phragmidiumpotentillae (Pers.) P. Karst. Here, we provide their sequences.
The hosts of Phragmidium discussed in this study are mainly from the genus Rubus within the Rosaceae family, yet P.coreanicola represents a new species on a previously reported host, while Gerwasiarubi-setchuenensis originates from a different host. This demonstrates the host specificity and species diversity of rust fungi (Wei 1988; Zhuang et al. 2012; Yang et al. 2015; Liu et al. 2018). Investigations into the interaction between plant hosts and pathogens suggest that the variety of pathogenic fungi may stem from host-switching or cooperative coevolutionary processes. These findings prompt inquiries concerning the linkage between plant hosts and Phragmidium, as well as the evolutionary dynamics at play (Zhao et al. 2016; McTaggart et al. 2015, 2016). Addressing these inquiries necessitates further research into Phragmidium in the future.
Supplementary Material
XML Treatment for Gerwasia amphidasydis
XML Treatment for Phragmidium coreanicola
XML Treatment for Phragmidium parvifolius
XML Treatment for Gerwasia rubi-setchuenensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aime MC (2006) Toward resolving family–level relationships in rust fungi (Uredinales).Mycoscience 47(3): 112–122. 10.1007/S 10267-006-0281-0 · doi ↗
- 2Aime MC Bell CD Wilson AW (2018) Deconstructing the evolutionary complexity between rust fungi (Pucciniales) and their plant hosts.Studies in Mycology 89: 143–152. 10.1016/j.simyco.2018.02.00229910520 PMC 6002339 · doi ↗ · pubmed ↗
- 3Aime MC Mc Taggart AR (2021) A higher-rank classification for rust fungi, with notes on genera.Fungal Systematics and Evolution 7(1): 21–47. 10.3114/fuse.2021.07.0234124616 PMC 8165960 · doi ↗ · pubmed ↗
- 4Aime MC Mc Taggart AR Mondo SJ Duplessis S (2017) Phylogenetics and phylogenomics of rust fungi.Advances in Genetics 100: 267–307. 10.1016/bs.adgen.2017.09.01129153402 · doi ↗ · pubmed ↗
- 5Ali B Sohail Y Mumtaz AS Berndt R (2017) Phragmidiumpunjabense, a new species of rust fungus on Rosabrunonii in the outer Himalayan ranges of Murree, Pakistan.Nova Hedwigia 105(3–4): 385–396. 10.1127/nova_hedwigia/2017/0418 · doi ↗
- 6Arthur JC (1934) Manual of the rust in United States and Canada, Hafner Publishing Company, New York, USA.
- 7Bradshaw MJ Carey J Liu M Bartholomew HP Jurick WM II Hambleton S Hendricks D Schnittler M Scholler M (2023) Genetic time traveling: Sequencing old herbarium specimens, including the oldest herbarium specimen sequenced from kingdom Fungi, reveals the population structure of an agriculturally significant rust.The New Phytologist 237(4): 1463–1473. 10.1111/nph.1862236385249 · doi ↗ · pubmed ↗
- 8Cummins GB (1931) Phragmidium species of North America: Differential teliospore and aecial characters.Mycologia 23(6): 433–445. 10.1080/00275514.1931.12017054 · doi ↗