Accurate identification of Enterococcus lactis causing bacteraemia by matrix-assisted laser desorption ionization-time of flight mass spectrometry
Marhami Fahriani, Geoffrey W. Coombs, Princy Shoby, Haley Hood, Denise A. Daley, Christopher A. Mullally, Shakeel Mowlaboccus

TL;DR
This study creates a new database to accurately identify Enterococcus lactis using a mass spectrometry technique, preventing misidentification with E. faecium.
Contribution
A custom MALDI Biotyper® database is developed for rapid and accurate identification of E. lactis in clinical settings.
Findings
The custom database correctly identified all E. lactis and E. faecium isolates at the species level.
84.2% of E. lactis isolates were identified at the high probable species level using the database.
Abstract
Introduction.Enterococcus faecium clade B has recently been re-classified as Enterococcus lactis. Although E. lactis was previously associated with food products and probiotics, the recent re-classification has prompted the need for the accurate identification of this species and re-interpretation of its disease-causing ability. Since the re-classified E. lactis can currently only be identified by molecular techniques such as whole-genome sequencing, we constructed a MALDI Biotyper® custom database to rapidly identify and differentiate E. lactis causing bacteraemia from E. faecium. Hypothesis/Gap statement. The re-classification of E. faecium clade B as E. lactis warrants the development of rapid and accurate identification methods to distinguish these species, particularly in clinical settings where E. lactis may be misidentified as E. faecium. Aim. The aim of this study was to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
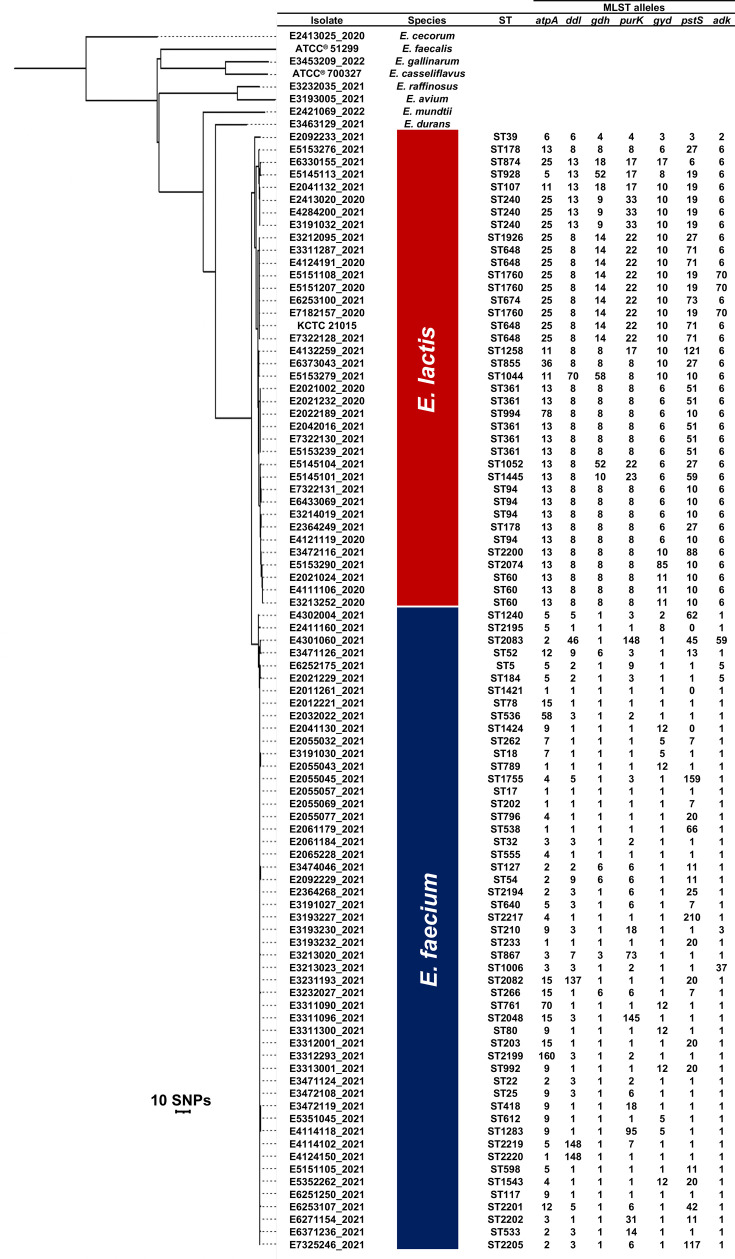 Fig. 1
Fig. 1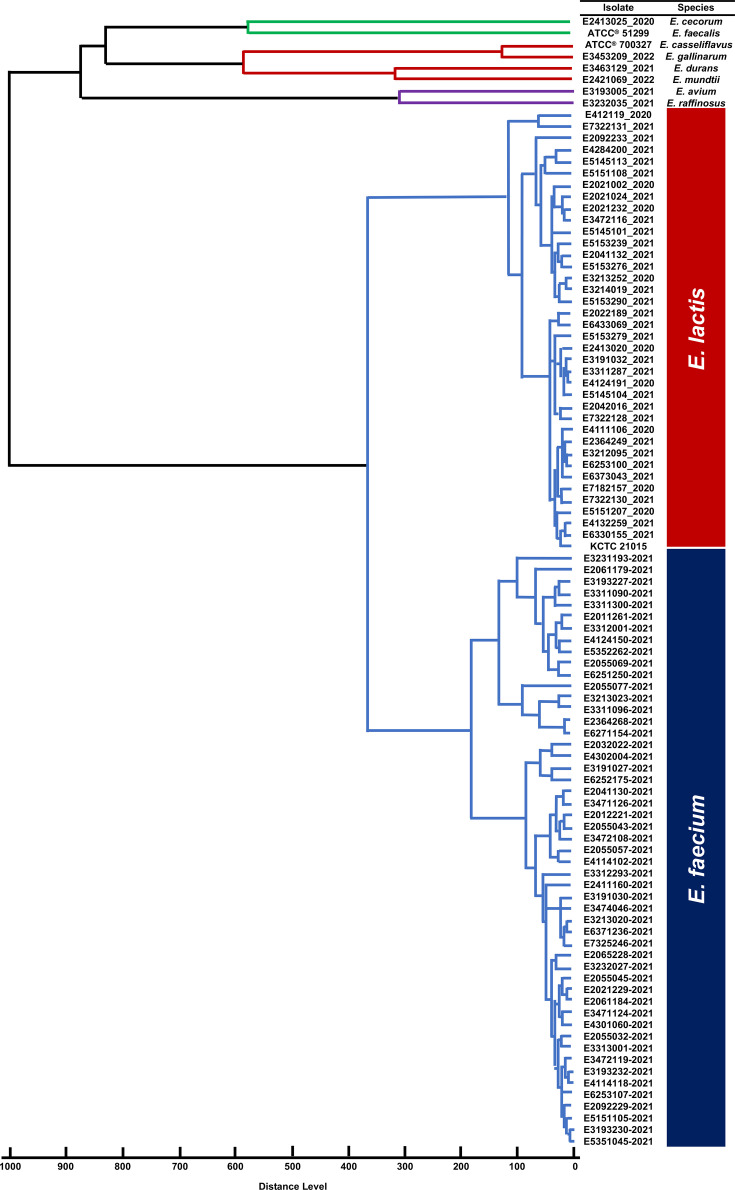 Fig. 2
Fig. 2| Sample | Source | Year | ST* | The BioTyper reference library (2022) | Elac_AGAR_MF (This study) | ||
| eDT† | Score | eDT† | Score | ||||
|
| |||||||
| KCTC 21015 | KCTC | – | ST648 |
| 2.30 |
| 2.45 |
| E2021002-2020 | AESOP | 2020 | ST361 |
| 2.11 |
| 2.26 |
| E2021232-2020 | AESOP | 2020 | ST361 |
| 2.19 |
| 2.49 |
| E2413020-2020 | AESOP | 2020 | ST240 |
| 2.34 |
| 2.48 |
| E3213252-2020 | AESOP | 2020 | ST60 |
| 2.28 |
| 2.56 |
| E4111106-2020 | AESOP | 2020 | ST60 |
| 2.2 |
| 2.41 |
| E4121119-2020 | AESOP | 2020 | ST94 |
| 2.02 |
| 2.51 |
| E4124191-2020 | AESOP | 2020 | ST648 |
| 2.29 |
| 2.63 |
| E5151207-2020 | AESOP | 2020 | ST1760 |
| 2.28 |
| 2.51 |
| E7182157-2020 | AESOP | 2020 | ST1760 |
| 2.26 |
| 2.55 |
| E2021024-2021 | AESOP | 2021 | ST60 |
| 2.27 |
| 2.51 |
| E2022189-2021 | AESOP | 2021 | ST994 |
| 2.25 |
| 2.44 |
| E2041132-2021 | AESOP | 2021 | ST107 |
| 2.1 |
| 2.1 |
| E2042016-2021 | AESOP | 2021 | ST361 |
| 2.33 |
| 2.5 |
| E2092233-2021 | AESOP | 2021 | ST39 |
| 2.24 |
| 2.33 |
| E2364249-2021 | AESOP | 2021 | ST178 |
| 2.25 |
| 2.53 |
| E3191032-2021 | AESOP | 2021 | ST240 |
| 2.24 |
| 2.45 |
| E3212095-2021 | AESOP | 2021 | ST1926 |
| 2.16 |
| 2.43 |
| E3214019-2021 | AESOP | 2021 | ST94 |
| 2.35 |
| 2.55 |
| E3311287-2021 | AESOP | 2021 | ST648 |
| 2.16 |
| 2.42 |
| E3472116-2021 | AESOP | 2021 | ST2200 |
| 2.22 |
| 2.44 |
| E4132259-2021 | AESOP | 2021 | ST1258 |
| 2.36 |
| 2.53 |
| E4284200-2021 | AESOP | 2021 | ST240 |
| 2.27 |
| 2.47 |
| E5145101-2021 | AESOP | 2021 | ST1445 |
| 2.14 |
| 2.22 |
| E5145104-2021 | AESOP | 2021 | ST1052 |
| 2.3 |
| 2.46 |
| E5145113-2021 | AESOP | 2021 | ST928 |
| 2.21 |
| 2.34 |
| E5151108-2021 | AESOP | 2021 | ST1760 |
| 2.28 |
| 2.45 |
| E5153239-2021 | AESOP | 2021 | ST361 |
| 2.38 |
| 2.56 |
| E5153276-2021 | AESOP | 2021 | ST178 |
| 2.14 |
| 2.00 |
| E5153279-2021 | AESOP | 2021 | ST1044 |
| 2.12 |
| 2.53 |
| E5153290-2021 | AESOP | 2021 | ST2074 |
| 2.28 |
| 2.37 |
| E6253100-2021 | AESOP | 2021 | ST674 |
| 2.22 |
| 2.63 |
| E6330155-2021 | AESOP | 2021 | ST874 |
| 2.24 |
| 2.54 |
| E6373043-2021 | AESOP | 2021 | ST855 |
| 2.25 |
| 2.4 |
| E6433069-2021 | AESOP | 2021 | ST94 |
| 2.02 |
| 2.2 |
| E7322128-2021 | AESOP | 2021 | ST648 |
| 2.17 |
| 2.33 |
| E7322130-2021 | AESOP | 2021 | ST361 |
| 2.26 |
| 2.2 |
| E7322131-2021 | AESOP | 2021 | ST94 |
| 2.09 |
| 2.46 |
|
| |||||||
| E2011261-2021 | AESOP | 2021 | ST1421 |
| 2.26 |
| 2.20 |
| E2012221-2021 | AESOP | 2021 | ST78 |
| 2.29 |
| 2.29 |
| E2021229-2021 | AESOP | 2021 | ST184 |
| 2.41 |
| 2.48 |
| E2032022-2021 | AESOP | 2021 | ST536 |
| 2.46 |
| 2.51 |
| E2041130-2021 | AESOP | 2021 | ST1424 |
| 2.45 |
| 2.44 |
| E2055032-2021 | AESOP | 2021 | ST262 |
| 2.5 |
| 2.52 |
| E2055043-2021 | AESOP | 2021 | ST789 |
| 2.47 |
| 2.50 |
| E2055045-2021 | AESOP | 2021 | ST1755 |
| 2.43 |
| 2.43 |
| E2055057-2021 | AESOP | 2021 | ST17 |
| 2.54 |
| 2.56 |
| E2055069-2021 | AESOP | 2021 | ST202 |
| 2.33 |
| 2.46 |
| E2055077-2021 | AESOP | 2021 | ST796 |
| 2.21 |
| 2.22 |
| E2061179-2021 | AESOP | 2021 | ST538 |
| 2.29 |
| 2.29 |
| E2061184-2021 | AESOP | 2021 | ST32 |
| 2.53 |
| 2.51 |
| E2065228-2021 | AESOP | 2021 | ST555 |
| 2.12 |
| 2.07 |
| E2092229-2021 | AESOP | 2021 | ST54 |
| 2.36 |
| 2.44 |
| E2364268-2021 | AESOP | 2021 | ST2194 |
| 2.38 |
| 2.35 |
| E2411160-2021 | AESOP | 2021 | ST2195 |
| 2.44 |
| 2.48 |
| E3191027-2021 | AESOP | 2021 | ST640 |
| 2.49 |
| 2.50 |
| E3191030-2021 | AESOP | 2021 | ST18 |
| 2.42 |
| 2.36 |
| E3193227-2021 | AESOP | 2021 | ST2217 |
| 2.51 |
| 2.48 |
| E3193230-2021 | AESOP | 2021 | ST210 |
| 2.47 |
| 2.51 |
| E3193232-2021 | AESOP | 2021 | ST233 |
| 2.38 |
| 2.34 |
| E3213020-2021 | AESOP | 2021 | ST867 |
| 2.42 |
| 2.42 |
| E3213023-2021 | AESOP | 2021 | ST1006 |
| 2.35 |
| 2.33 |
| E3231193-2021 | AESOP | 2021 | ST2082 |
| 2.31 |
| 2.28 |
| E3232027-2021 | AESOP | 2021 | ST266 |
| 2.51 |
| 2.55 |
| E3311090-2021 | AESOP | 2021 | ST761 |
| 2.31 |
| 2.44 |
| E3311096-2021 | AESOP | 2021 | ST2048 |
| 2.52 |
| 2.51 |
| E3311300-2021 | AESOP | 2021 | ST80 |
| 2.31 |
| 2.27 |
| E3312001-2021 | AESOP | 2021 | ST203 |
| 2.33 |
| 2.34 |
| E3312293-2021 | AESOP | 2021 | ST2199 |
| 2.52 |
| 2.49 |
| E3313001-2021 | AESOP | 2021 | ST992 |
| 2.41 |
| 2.45 |
| E3471124-2021 | AESOP | 2021 | ST22 |
| 2.27 |
| 2.44 |
| E3471126-2021 | AESOP | 2021 | ST52 |
| 2.23 |
| 2.32 |
| E3472108-2021 | AESOP | 2021 | ST25 |
| 2.53 |
| 2.56 |
| E3472119-2021 | AESOP | 2021 | ST418 |
| 2.53 |
| 2.54 |
| E3474046-2021 | AESOP | 2021 | ST127 |
| 2.45 |
| 2.43 |
| E4114102-2021 | AESOP | 2021 | ST2219 |
| 2.57 |
| 2.58 |
| E4114118-2021 | AESOP | 2021 | ST1283 |
| 2.41 |
| 2.48 |
| E4124150-2021 | AESOP | 2021 | ST2220 |
| 2.57 |
| 2.62 |
| E4301060-2021 | AESOP | 2021 | ST2083 |
| 1.73 |
| 2.03 |
| E4302004-2021 | AESOP | 2021 | ST1240 |
| 2.27 |
| 2.37 |
| E5151105-2021 | AESOP | 2021 | ST598 |
| 2.43 |
| 2.38 |
| E5351045-2021 | AESOP | 2021 | ST612 |
| 2.5 |
| 2.53 |
| E5352262-2021 | AESOP | 2021 | ST1543 |
| 2.42 |
| 2.33 |
| E6251250-2021 | AESOP | 2021 | ST117 |
| 2.4 |
| 2.37 |
| E6252175-2021 | AESOP | 2021 | ST5 |
| 2.37 |
| 2.34 |
| E6253107-2021 | AESOP | 2021 | ST2201 |
| 2.45 |
| 2.46 |
| E6271154-2021 | AESOP | 2021 | ST2202 |
| 2.57 |
| 2.56 |
| E6371236-2021 | AESOP | 2021 | ST533 |
| 2.41 |
| 2.43 |
| E7325246-2021 | AESOP | 2021 | ST2205 |
| 2.43 |
| 2.46 |
|
| |||||||
| E3193005-2021 | AESOP | 2021 |
|
| 2.25 |
| 2.17 |
| E2413025-2020 | AESOP | 2020 |
|
| 2.21 |
| 2.15 |
| E3463129-2021 | AESOP | 2021 |
|
| 2.2 |
| 2.21 |
| E3453209-2022 | AESOP | 2022 |
|
| 2.05 |
| 2.24 |
| E2421069-2022 | AESOP | 2022 |
|
| 2.25 |
| 2.18 |
| E3232035-2021 | AESOP | 2021 |
|
| 2.15 |
| 2.22 |
| ATCC 700327 | ATCC | – |
|
| 2.13 |
| 2.15 |
| ATCC 51299 | ATCC | – |
|
| 2.15 |
| 2.12 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial Identification and Susceptibility Testing · Antimicrobial Resistance in Staphylococcus · Infective Endocarditis Diagnosis and Management
Background
Enterococcus faecium, a Gram-positive bacterium, is a member of the ESKAPE organisms. ESKAPE consists of six virulent nosocomial bacterial pathogens (E. faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa and Enterobacter species) that have developed resistance mechanisms to multiple classes of antibiotics [1]. In 2024, the World Health Organization included vancomycin-resistant E. faecium on the high-priority list of bacteria, for which it is crucial to conduct research and develop strategies to prevent and control antimicrobial resistance [2]. Vancomycin-resistant E. faecium poses a public health challenge due to persistent infections and multidrug resistance, which limit treatment options for E. faecium infections [3].
Based on whole-genome sequences, E. faecium isolates are categorized into two phylogenetically distinct clades: clade A isolates, which are typically hospital-associated, and clade B isolates, which are typically community-associated [4]. Clade A isolates are further divided into subclade A1, consisting of clinical isolates, and subclade A2, consisting of animal-associated isolates. Unlike the multidrug-resistant clade A isolates, the non-hospital clade B isolates are typically susceptible to antibiotics. Furthermore, clade B isolates, often isolated as commensals, have fewer virulence genes and mobile genetic elements than subclade A1 isolates and thus have a smaller genome size [5].
Recently, E. faecium clade B isolates have been reclassified as a different species, Enterococcus lactis [4]. E. lactis, first reported in 2012 from Italian raw milk cheese, is closely related to E. faecium with 99.4% similarity based on 16S rRNA gene sequence analysis [6]. E. lactis is frequently isolated in milk and dairy products [67]. The benefits and safety of E. lactis as a probiotic product for humans have previously been studied. The probiotic E. lactis strain IITRHR1 has a cytoprotective potential against drug-induced liver injury [8], and the probiotic E. lactis strain IW5 has anticancer properties against several human cancer cell lines [9]. E. lactis may also inhibit the growth of pathogenic bacteria, including Escherichia coli O26, S. aureus, Bacillus cereus, K. pneumoniae, Shigella flexneri, Streptococcus mutans and Listeria monocytogenes [9]. In the food industry, E. lactis has been used as a starter [10], probiotic culture [11] and animal feed additive [12]. Although previously presumed safe, E. lactis, which is susceptible to ampicillin, can still cause severe disease in humans [13]. Therefore, due to potential safety concerns, in 2022, E. lactis was removed from the European Food Safety Authority’s Qualified Presumption of Safety [14]. Furthermore, E. lactis can harbour antimicrobial resistance genes – for example, E. lactis strain E843, isolated in China from faecal samples of pigs, harbours the poxtA gene, which encodes resistance to the last-line drug linezolid [15].
In Australia, a nationwide antimicrobial resistance surveillance programme on enterococcal bloodstream infections, known as the Australian Enterococcal Surveillance Outcome Programme (AESOP), has been performed by the Australian Group on Antimicrobial Resistance (AGAR) since 2013. The programme involves 33 laboratories servicing 55 hospitals from all Australian states and mainland territories. In addition to antimicrobial susceptibility testing, whole-genome sequencing (WGS) is performed on all isolates identified by the laboratories as E. faecium. In AESOP 2021, of the 1,297 enterococcal bloodstream infections reported, 523 (40.3%) were identified as E. faecium bacteraemia cases by the AGAR laboratories. However, WGS identified 28 (2.16%) E. lactis isolates that were incorrectly reported as E. faecium by the laboratories [16]. Similarly, in AESOP 2022, 29 of the 1,535 (1.89%) enterococcal isolates were misidentified as E. faecium by the AGAR laboratories and were confirmed to be E. lactis following WGS [14].
In Australian diagnostic microbiology laboratories, matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS) is frequently used for bacterial species identification [17]. MALDI-TOF MS is a cost-effective and easy-to-perform assay that accurately identifies pathogenic organisms with minimal hands-on time and optimal turnaround time [18]. However, the two MALDI-TOF MS systems used in diagnostic microbiology laboratories, the MALDI Biotyper^®^ (Bruker Daltonics, Germany) and VITEK^®^ MS (bioMérieux, France), do not have E. lactis reference spectra in their databases and consequently are not able to distinguish E. lactis from E. faecium [17]. In this study, we aim to construct a MALDI Biotyper^®^ in-house database to rapidly identify and differentiate E. lactis from E. faecium.
Methods
Bacterial isolates
A total of 97 isolates representing ten different Enterococcus species (E. avium, E. casseliflavus, E. cecorum, E. durans, E. faecalis, E. faecium, E. gallinarum, E. lactis, E. mundtii and E. raffinosus) were included. All isolates were cultured on blood agar and incubated at 37 °C overnight. E. cecorum was incubated in the presence of 5% CO_2_. The clinical * E. faecium* and E. lactis isolates were obtained from bacteraemia cases reported in the 2020 and 2021 AESOP surveys. The E. faecalis (ATCC^®^ 51299) and E. casseliflavus (ATCC^®^ 700327) reference strains were obtained from the American Type Culture Collection (ATCC). The E. lactis reference strain KCTC 21015 was obtained from the Korean Collection for Type Cultures (KCTC).
Whole-genome sequencing
Bacterial genomic DNA was extracted using the DNeasy^®^ Blood and Tissue kit (Qiagen, Germany) according to the manufacturer’s instructions. Genomic DNA was quantified using a Qubit^™^ 3.0 Fluorometer (Thermo Fisher Scientific Inc., USA). DNA libraries were prepared using the Nextera XT Library Preparation kit (Illumina Inc., USA) and sequenced on the Illumina NextSeq^™^ 500 platform (Illumina Inc., USA) using 150 bp paired-end chemistry. The raw sequence reads have been deposited in the sequence read archive under BioProject ID PRJNA1062579.
Bioinformatics analysis
Raw sequence reads were trimmed using Trimmomatic version 0.39 and assembled de novo using SPAdes version 3.15.4 [19]. The sequence type (ST) of each E. faecium and E. lactis isolate was determined in silico using the E. faecium multilocus sequence typing (MLST) scheme described by Homan et al. [20]. Species identification was performed using the Species ID tool on the PubMLST website. Species ID uses an in silico ribosomal MLST approach based on 53 loci to identify the genus and species of bacterial isolates [21].
Phylogenetic reconstruction
Single nucleotide polymorphisms (SNPs) were identified by aligning the sequence reads of each isolate to the chromosome of the E. faecium strain SRR24 (accession no. CP038996.1) or E. lactis KCTC 21015 (accession no. CP065211.1) using Snippy version 4.6.0 [22]. Phylogenetic trees were constructed using the neighbour-joining algorithm based on the SNP alignment using 200 bootstrap replicates, and the dendrogram was visualized and annotated on the interactive Tree of Life (iTOL) website [23].
Sample preparation for spectral database construction
For the construction of the spectral database library, proteins were extracted from bacterial colonies using the ethanol/formic acid/acetonitrile protocol as recommended by the manufacturer. Briefly, pure bacterial colonies were transferred to 300 µl of HPLC-grade water using a 1 µl inoculation loop and mixed with 900 µl absolute ethanol. The suspension was then centrifuged for 2 min at 13,300 r.p.m. and the supernatant was removed. After air-drying for 5 min at room temperature, the pellet was resuspended in 25 µl of 70% formic acid and 25 µl of 100% acetonitrile, followed by centrifugation for 2 min at 13,300 r.p.m. A 1 µl volume of the supernatant was then applied to a single empty spot on the MALDI target plate and allowed to dry at room temperature. The spots were overlaid with 1 µl of α-cyano-4-hydroxycinnamic acid (HCCA) matrix solution and allowed to dry at room temperature. The target plate was loaded onto the Bruker Microflex^®^ LT/SH bench-top mass spectrometer.
MALDI-TOF MS parameters
Analysis was performed using the Bruker Microflex^®^ LT/SH bench-top mass spectrometer (Bruker Daltonics GmbH, Bremen, Germany) with FlexControl software version 3.4. Calibration and quality control were performed using the protein extract from E. coli strain DH5α as a test standard. Data were obtained in automatic mode by collecting 40 laser shots with 40% laser intensity. Spectra were recorded in a positive linear mode (ion source one voltage=20.00 kV; ion source two voltage=18.20 kV; lens voltage=6.00 kV; laser frequency=60 Hz; mass range=2,000–20,000 Da).
Analysis of raw spectra for E. lactis main spectrum profiles
The main spectrum profiles (MSPs) of 29 E. lactis isolates (the KCTC 21015 reference isolate and 28 isolates from AESOP 2021) were used to construct the in-house database. In total, 36 replicates of the spectra of each isolate were generated. Raw spectra quality was evaluated using FlexAnalysis software version 3.4 as per the manufacturer’s recommendation (Bruker Daltonics GmbH, Bremen, Germany). Baseline subtraction and smoothing were performed. Background noises, such as flatline spectra, outliers, dramatic mass shifts and anomalies, were deleted. At least 20 high-quality spectra per isolate were selected and transferred to the MBT Compass Explorer^®^ version 4.1.100 to create a single main spectrum for each E. lactis isolate.
Identifying species using the in-house database
The extended direct transfer protocol commonly used by diagnostic laboratories was used for species identification. Briefly, one colony was lifted using a 1 µl sterile plastic loop and smeared as a thin film on a single empty spot on the target plate as described by Patel (2013) [24]. The spot was treated with 1 µl of 70% formic acid, allowed to dry at room temperature and overlaid with 1 µl of HCCA solution. Following a short drying period, the plate was loaded onto a Bruker Microflex^®^ LT/SH bench-top mass spectrometer. Prior to using the constructed in-house database, all isolates were identified using the BioTyper^®^ MBT Compass reference library (2022), which contains 11,897 main spectra of 4,274 species from 704 micro-organism genera.
The MALDI-TOF MS analysis results, expressed with a log score value (0.000–3.000), were indicative of the closest species matched: a score ≥2.300 was considered a high probable species identification; 2.000–2.299, a probable species identification; 1.700–1.999, a probable genus identification; and <1.700, no reliable identification. The log score value was calculated after determining the matching mass signal between the reference spectra and the unknown spectra (no match=0, complete match=1), and vice versa, and the symmetry of the matching signal intensity between the unknown and reference spectra (high symmetry yields a value close to 1, no symmetry yields a value close to 0). These three scores were then multiplied, normalized to a value of 1,000 and converted to common logarithm yielding a maximum value of 3 (i.e. log 1,000) as previously described [25].
An MSP dendrogram was generated using the MBT Compass Explorer^®^ software module version 4.1.100 and constructed using a correlation distance and average-linkage algorithm with a threshold value of 400 for a single organism. In the MSP dendrogram, the closeness of organisms is reflected by an arbitrary distance level. The distance level in an MSP dendrogram is normalized to a maximum value of 1,000.
Statistical analysis
Using WGS as the gold standard, the sensitivity, specificity, positive predictive value (PPV) and negative predictive value (NPV) of the MALDI-TOF MS identification were calculated accordingly [26], using the following definitions: true positive – both the MALDI-TOF MS and WGS results were E. faecium; false positive – only the MALDI-TOF MS result was E. faecium; false negative – only the WGS result was E. faecium; and true negative – no E. faecium was identified by either method.
Results
Genomic diversity of enterococcal isolates
A total of 97 isolates, including 38 E. lactis, 51 E. faecium and 8 non-E. faecium non-E. lactis (NFL) enterococci, were included in the study (Table 1). Each E. faecium isolate (n=51), selected from the AESOP 2021 survey, had a unique ST. The E. lactis isolates included the KCTC 21015 strain and all E. lactis isolates misidentified as E. faecium in the AESOP 2020 (n=9) and AESOP 2021 (n=28) surveys. The eight NFL enterococcal isolates included two strains from the ATCC^®^ collection, E. faecalis ATCC^®^ 51299^™^ and E. casseliflavus ATCC^®^ 700327^™^, and single isolates of E. avium, E. cecorum, E. durans, E. gallinarum, E. mundtii and E. raffinosus from the AESOP surveys. The species identity of each isolate was confirmed by WGS and the SpeciesID tool. The genomes of all isolates were aligned to the chromosome of the E. faecium strain SRR24 and a total of 587 SNPs were identified. An SNP phylogenetic tree was constructed to visualize the relatedness of the isolates, and although closely related, all E. lactis isolates clustered separately from the E. faecium isolates (Fig. 1). A similar phylogeny was observed when the genomes were aligned to the chromosome of the E. lactis strain KCTC 21015 (Fig. S1, available in the online Supplementary Material).
SNP-based phylogenetic tree of enterococcal genomes. The phylogeny was inferred based on 587 SNPs identified after aligning the genomes of E. faecium (n=51) (blue cluster), E. lactis (n=38) (red cluster) and non-E. faecium non-E. lactis enterococci (n=8) to the chromosome of the reference E. faecium strain SRR24 (accession no. CP038996.1) using a neighbour-joining algorithm. The non-faecium non-lactis enterococcal species included E. avium, E. casseliflavus, E. cecorum, E. durans, E. faecalis, E. gallinarum, E. mundtii and E. raffinosus. The ST and the MLST allele numbers were obtained from the PubMLST website using the MLST scheme designed by Homan et al. [20]. The scale bar represents the number of SNPs.
Among the 38 E. lactis isolates, 21 STs were identified, 7 of which were represented by more than one isolate: ST361 (n=5), ST94 (n=4), ST60 (n=3), ST240 (n=3), ST648 (n=4), ST1760 (n=3) and ST178 (n=2). Except for atpA allele 5 and gyd allele 8, the MLST alleles harboured by E. lactis and E. faecium in this study were species-restricted. The atpA allele 5 was identified in seven E. faecium isolates (ST5, ST184, ST598, ST640, ST1240, ST2195 and ST2219) and one E. lactis isolate (ST928), while the gyd allele 8 was identified in one E. faecium isolate (ST2195) and one E. lactis isolate (ST928).
E. lactis isolates are misclassified as E. faecium using the current Bruker commercial database
The 51 E. faecium and 8 NFL enterococci were correctly identified at the species level using the BioTyper^®^ MBT Compass reference library (2022). However, all E. lactis isolates (n=38) were misidentified as E. faecium prior to the construction of the custom in-house database (Table 1).
Construction and validation of the Elac_AGAR_MF in-house spectral database
The custom in-house spectral database, named Elac_AGAR_MF (.btmsp file available in Supplementary Material), was constructed using the KCTC 21015 E. lactis isolate and the 28 E. lactis isolates, confirmed by WGS, from the AESOP 2021 survey [16]. To evaluate the performance and validate the constructed in-house spectral database, species identification of all 97 enterococcal isolates confirmed by WGS was re-assessed by MALDI-TOF after integrating the Elac_AGAR_MF database with the BioTyper^®^ MBT Compass reference library (2022).
All E. lactis isolates (n=38) were accurately identified as E. lactis using the custom in-house database, including 84.2% (n=32/38) identified at the high probable species level (score ≥2.300) and 15.8% (n=6/38) identified at the probable species level (score 2.000–2.299). All E. faecium isolates (n=51) were accurately identified as E. faecium, including 84.3% (n=43/51) identified at the high probable species level and 15.7% (n=8/51) identified at the probable species level. The NFL isolates (n=8/8) were also accurately identified at the species level. The species identity and log score value obtained for each isolate before and after integration of the custom Elac_AGAR_MF spectral database are provided in Table 1. The phyloproteomic tree constructed based on the MSP identification using the MBT Compass Explorer^®^ software showed E. faecium and E. lactis formed two distinct clusters, separate from the cluster containing the NFL isolates (Fig. 2).
Phyloproteomic tree of enterococcal isolates based on the MSP identification of Elac_AGAR_MF using MBT Compass Explorer®. A total of 97 enterococcal isolates that separated into E. faecium cluster (n=51, blue), E. lactis cluster (n=38, red) and non-E. faecium non-E. lactis enterococci (n=8). The MSP was constructed using correlation distance, average linkage and a threshold of 400 for a single organism.
Performance comparison before and after integrating the Elac_AGAR_MF in-house spectral database
The sensitivity, specificity, PPV and NPV for E. faecium identification by MALDI TOF MS using the BioTyper^®^ MBT Compass reference library (2022) were 100% (n=51/51), 17.4% (n=8/46), 57.3% (n=51/89) and 100% (n=8/8), respectively. Following the integration of the custom Elac_AGAR_MF in-house spectral database, the sensitivity, specificity, PPV and NPV for E. faecium identification were 100% each. Identification of E. lactis by MALDI-TOF MS could only be performed following the integration of the Elac_AGAR_MF custom database, and the sensitivity, specificity, PPV and NPV were 100% each (Table S1).
Discussion
The re-classification of E*. faecium* clade B as E. lactis has prompted the need for diagnostic techniques, other than WGS, to accurately distinguish E. lactis from E. faecium. In this study, we have successfully constructed a database for the rapid identification of E. lactis using the MALDI Biotyper^®^ (Bruker Daltonics, Germany), which can be implemented in diagnostic and research laboratories. Although the re-classification of E. faecium clade B as a different species (i.e. E. lactis) has been debated, phylogenetic and phyloproteomic data from our study support the idea that E. faecium and E. lactis isolates are dissimilar enough to be characterized as two separate groups. Furthermore, our data show that the Homan et al. [20] E. faecium MLST scheme can differentiate between E. faecium and E. lactis, given that the STs of our isolates was species-specific.
Unlike the study performed by Kim et al. [17] in which the investigators used three E. lactis reference strains to construct their in-house database on the MALDI Biotyper^®^, we constructed the Elac_AGAR_MF database using 29 genetically diverse E. lactis isolates representing 21 different sequence types. Kim et al. assessed the ability of the MALDI Biotyper^®^ to differentiate enterococcal strains isolated from fermented foods, while our study included clinical strains of enterococci. Our study included clinical * E. lactis* isolates retrieved from recent bacteraemia episodes, highlighting the need not to underestimate this species’ ability to cause severe disease. The antimicrobial susceptibility profile of E. lactis is different to that of E. faecium. Unlike most E. faecium, E. lactis isolates are typically susceptible to ampicillin as they do not harbour a pbp5-R allele [27]. Furthermore, vancomycin resistance in E. lactis is rare [28]. All E. lactis isolates in our collection were ampicillin-susceptible and vancomycin-susceptible [1629]. Therefore, resolving the misidentification of E. lactis as E. faecium is important to direct proper therapeutic options for E. lactis infections, which would include prescribing ampicillin, the treatment of choice for enterococcal infections caused by enterococci that lack mechanisms for high-level ampicillin resistance, thus limiting the overuse of vancomycin.
Two commercial MALDI-TOF MS platforms commonly used in Australian diagnostic laboratories are the MALDI Biotyper^®^ (Bruker Daltonics) and the VITEK^®^ MS (bioMérieux). Although mass spectrometry has revolutionized the field of diagnostic clinical microbiology for being a fast, reliable and inexpensive technique, the analytical performance of a mass spectrometer is significantly reliant on the diversity of reference spectra in its database. Currently, the commercial platforms do not contain an E. lactis reference spectrum in their database and thus misidentify E. lactis isolates as E. faecium (i.e. an ESKAPE organism), which, in the clinical setting, may lead to an unnecessary escalation in management of patients. In our study, the integration of the custom Elac_MF_AGAR database enhanced the performance of the MALDI BioTyper^®^, allowing for the accurate identification of E. lactis and E. faecium with a sensitivity and specificity value of 100%. Since we have shown the MALDI Biotyper^®^ can accurately identify E. lactis, a similar study is warranted to assess the ability of the VITEK^®^ MS to identify E. lactis to better equip our diagnostic laboratories.
While the ethanol/formic acid/acetonitrile protocol was used to prepare our custom library, the extended direct transfer protocol was used to validate the performance of the in-house library. The ethanol/formic acid/acetonitrile protocol, also known as the in-tube extraction method, yields high-quality spectra and is thus recommended for constructing custom libraries [30]. However, the extended direct transfer protocol is faster and is typically used in diagnostic laboratories to test bacterial isolates. Thus, we ensured the MALDI Biotyper^®^ yielded accurate results by validating our constructed database using the extended direct transfer protocol, allowing diagnostic laboratories to implement the Elac_AGAR_MF database onto their mass spectrometer without altering their sample preparation protocol.
Although our study accurately assigned the species of all isolates with a score >2.000, 84.3% of E. faecium and 84.2% of E. lactis were identified with a score ≥2.300 (high probable species level). The lower identification scores (score 2.000–2.299) observed in 22.7% (n=22/97) of isolates, while still sufficient for accurate identification, can be attributed to several factors. Variations in the amount of colony material and the homogeneity of the smear in this study can affect the spectral quality [3132]. Factors influencing the intensities of signal peaks, such as the age of bacterial culture, concentration and location of protein in the bacterial cell as well as level of protein expression, also play a significant role in the identification score [25]. Additionally, regular calibration of the MALDI-TOF MS device is essential to maintain high spectral quality [31].
Since our study only included E. lactis isolated from bacteraemia cases in Australia, clinical and non-clinical E. lactis from other geographical locations should be tested in the future to further validate the Elac_AGAR_MF database. Furthermore, more non-E. faecium non-E. lactis enterococcal strains, as well as non-enterococcal bacterial isolates, should be tested to ensure high sensitivity and high specificity of the Elac_AGAR_MF database.
Conclusions
We have shown that MALDI-TOF can be used to differentiate E. lactis from E. faecium in the clinical setting if E. lactis representatives are added to the spectral database. We urge commercial platforms to update their standard spectral database to incorporate * E. lactis* spectra to resolve the misidentification of E. lactis as E. faecium in the clinical setting where antibiotic prescription depends significantly on the accurate speciation of bacterial pathogens.
Availability of data and materials
The datasets supporting the conclusions of this article are included within the article and the Supplementary Material. The raw sequence reads have been deposited in the sequence read archive under BioProject ID PRJNA1062579.
supplementary material
10.1099/jmm.0.001995Uncited Supplementary Material 1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Boucher HW Talbot GH Bradley JS Edwards JE Gilbert D et al Bad bugs, no drugs: no ESKAPE! An update from the Infectious Diseases Society of America Clin Infect Dis 20094811210.1086/59501119035777 · doi ↗ · pubmed ↗
- 2World Health Organization WHO bacterial priority pathogens list, 2024: Bacterial pathogens of public health importance to guide research, development and strategies to prevent and control antimicrobial resistance Geneva: World Health Organization 2024 https://www.who.int/publications/i/item/9789240093461 accessed 27-November-2024
- 3Eisenberger D Tuschak C Werner M Bogdan C Bollinger T et al Whole-genome analysis of vancomycin-resistant Enterococcus faecium causing nosocomial outbreaks suggests the occurrence of few endemic clonal lineages in Bavaria, Germany J Antimicrob Chemother 2020751398140410.1093/jac/dkaa 04132083651 · doi ↗ · pubmed ↗
- 4Belloso Daza MV Cortimiglia C Bassi D Cocconcelli PS Genome-based studies indicate that the Enterococcus faecium Clade B strains belong to Enterococcus lactis species and lack of the hospital infection associated markers Int J Syst Evol Microbiol 20217110.1099/ijsem.0.00494834402778 · doi ↗ · pubmed ↗
- 5Lebreton F van Schaik W Mc Guire AM Godfrey P Griggs A et al Emergence of epidemic multidrug-resistant Enterococcus faecium from animal and commensal strainsm Bio 20134 e 00534-1310.1128/m Bio.00534-1323963180 PMC 3747589 · doi ↗ · pubmed ↗
- 6Morandi S Cremonesi P Povolo M Brasca M Enterococcus lactis sp. nov., from Italian raw milk cheeses Int J Syst Evol Microbiol 2012621992199610.1099/ijs.0.030825-022003038 · doi ↗ · pubmed ↗
- 7Sharma A Lavania M Singh R Lal B Identification and probiotic potential of lactic acid bacteria from camel milk Saudi J Biol Sci 2021281622163210.1016/j.sjbs.2020.11.06233732048 PMC 7938203 · doi ↗ · pubmed ↗
- 8Sharma S Chaturvedi J Chaudhari BP Singh RL Kakkar P Probiotic Enterococcus lactis IITRHR 1 protects against acetaminophen-induced hepatotoxicity Nutrition 20122817318110.1016/j.nut.2011.02.01221778036 · doi ↗ · pubmed ↗