Functional analysis of type II chalcone isomerase (CHI) genes in regulating soybean (Glycine max L.) nodule formation
Xinyue Wang, Jingwen Li, Yuxue Zhou, Jinhao Zhang, Le Wang, Yajing Liu, Xuguang Yang, Hongshuang Han, Qingyu Wang, Ying Wang

TL;DR
This study explores how specific chalcone isomerase genes in soybeans affect nodule formation, which is important for nitrogen fixation.
Contribution
The study identifies functional differences among three GmCHI gene variants in promoting soybean nodulation and isoflavone production.
Findings
GmCHI1A and GmCHI1B1 promote soybean nodulation, with GmCHI1A having a stronger effect.
Overexpression of GmCHI1A increases daidzein and genistein levels, while GmCHI1B1 increases glycitein.
Functional differentiation among homologous GmCHI genes suggests potential for improving nitrogen fixation in soybeans.
Abstract
Biological nitrogen fixation (BNF) is the most cost-effective and environmentally benign method for nitrogen fertilization. Isoflavones are important signaling factors for BNF in leguminous plants. Whether chalcone isomerase (CHI), the key enzyme gene in the flavonoid synthesis pathway, contributes to soybean (Glycine max) nodulation has not yet been fully clarified. In the present study, we identified the functions of three types of GmCHI for BNF using a hairy root system. The results showed that GmCHI1A and GmCHI1B1 positively increased nodulation while GmCHI1B2 did not, with the GmCHI1A gene having a greater effect than GmCHI1B1. Meanwhile, the daidzein and genistein contents were significantly increased in composite plants overexpressing GmCHI1A and reduced in composite plants, thus interfering with GmCHI1A. However, overexpression of GmCHI1B1 significantly increased the content of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
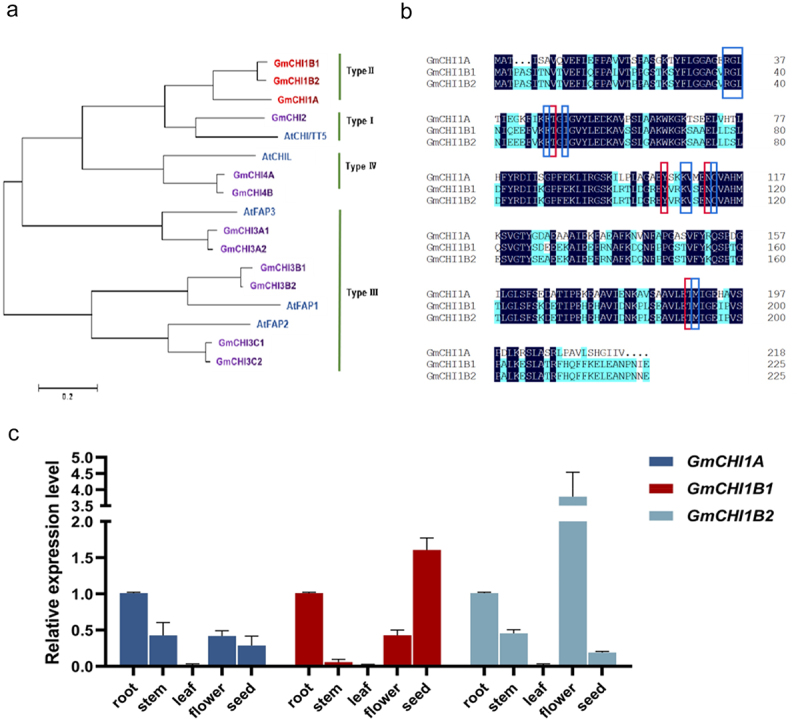 Figure 1
Figure 1 Figure 2
Figure 2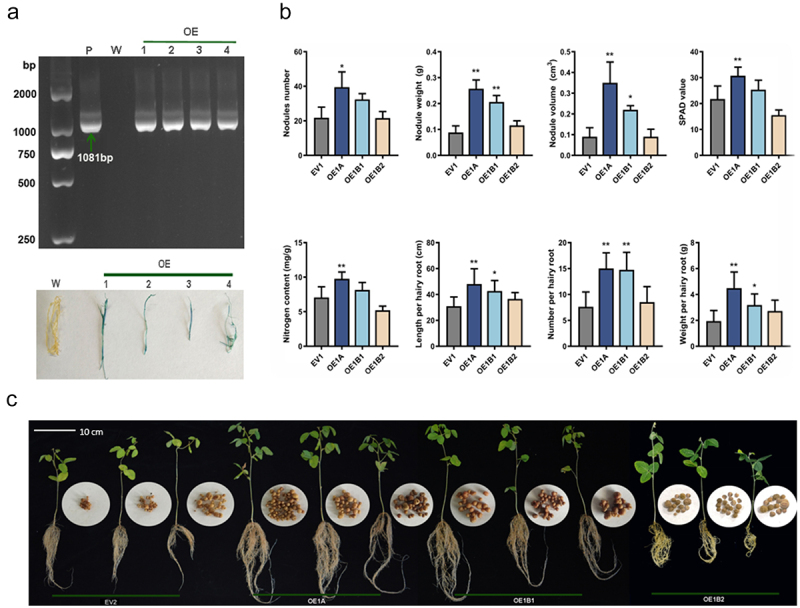 Figure 3
Figure 3 Figure 4
Figure 4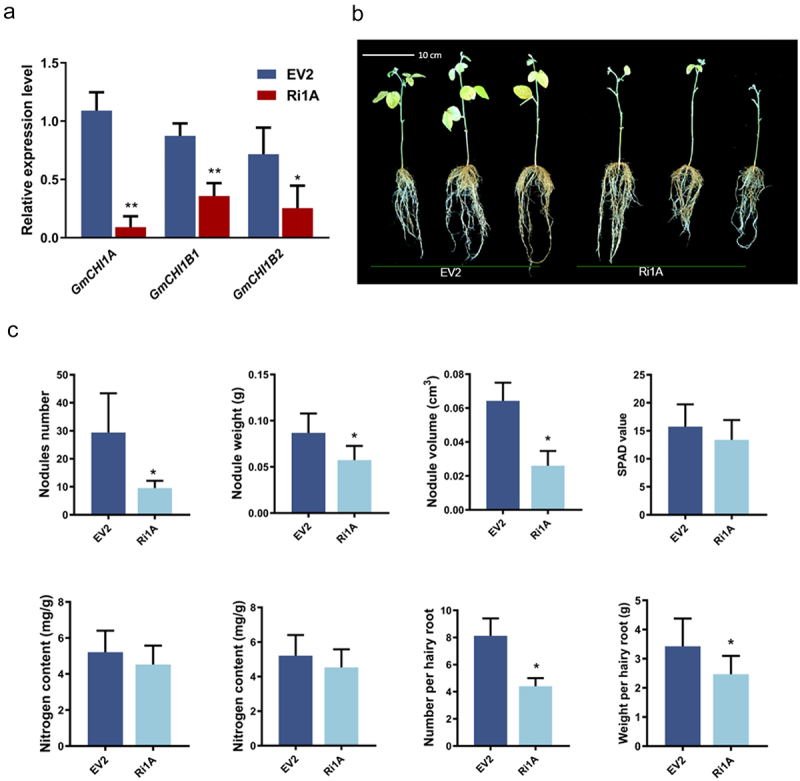 Figure 5
Figure 5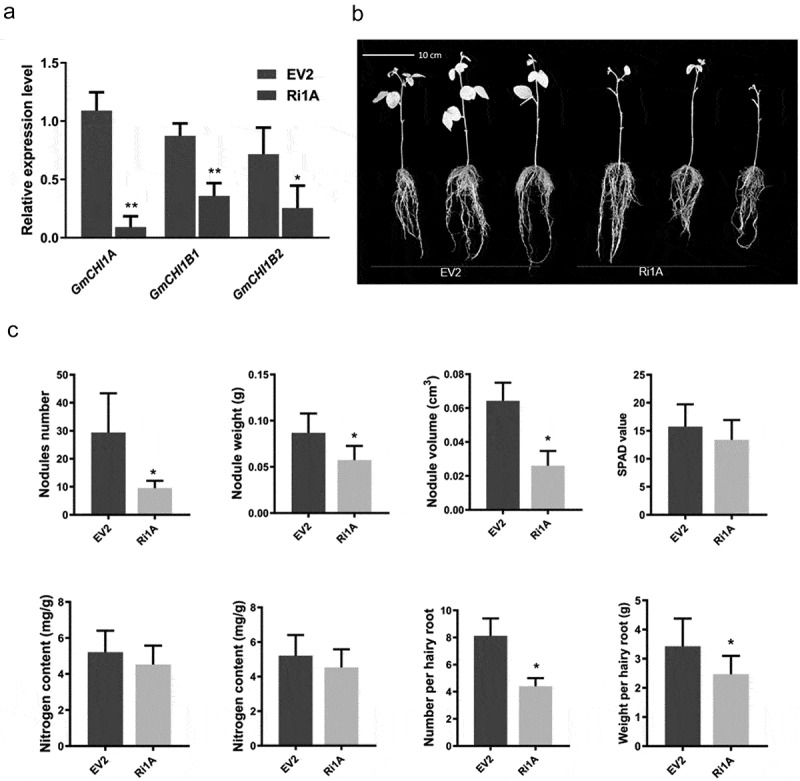 Figure 6
Figure 6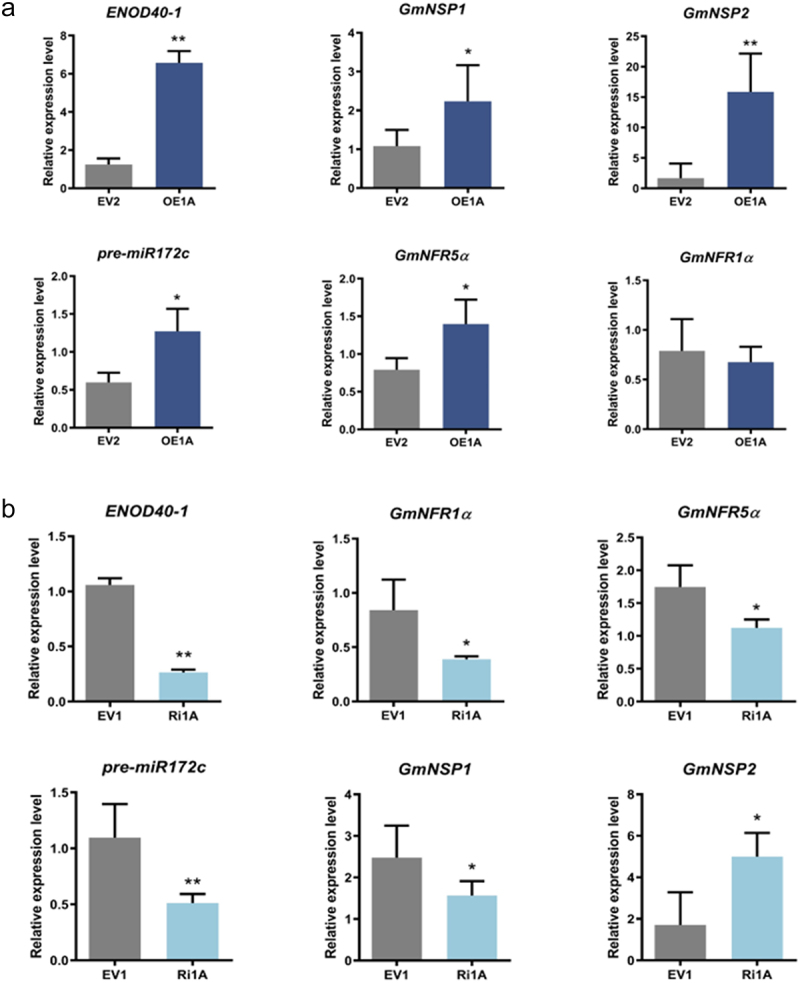 Figure 7
Figure 7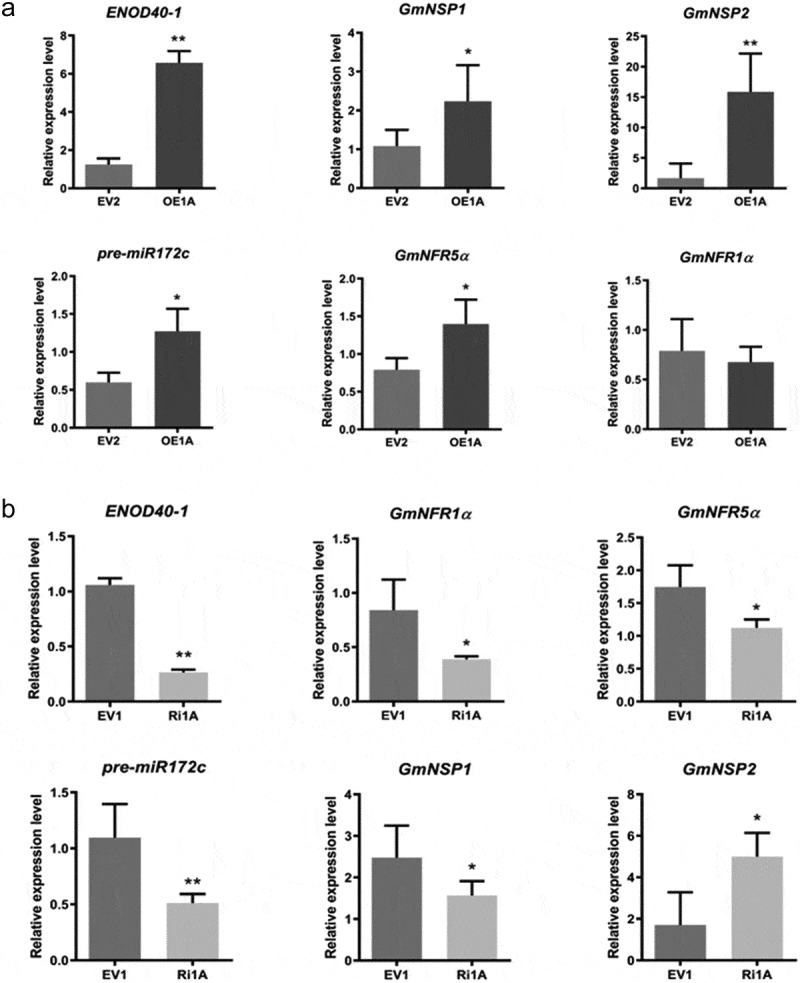 Figure 8
Figure 8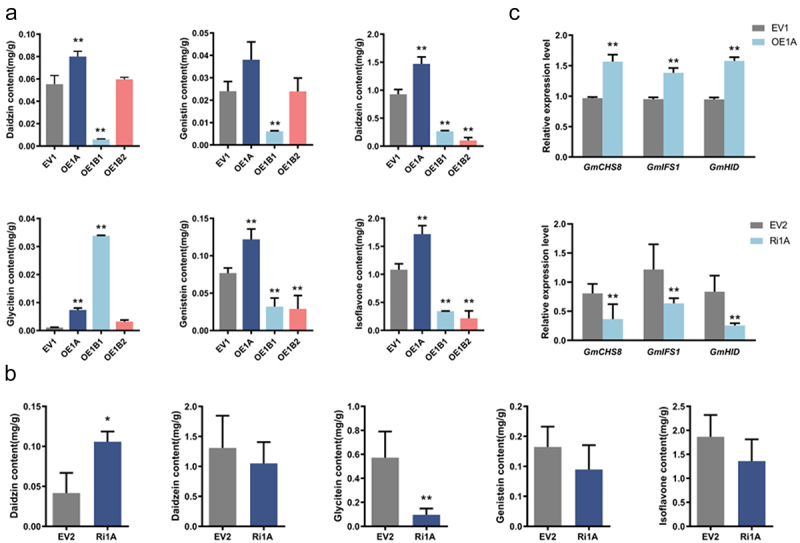 Figure 9
Figure 9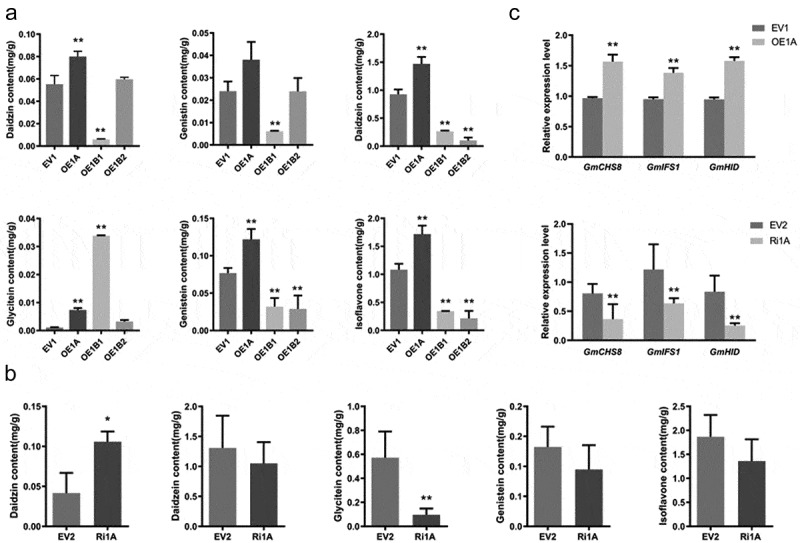 Figure 10
Figure 10- —National Natural Science Foundation of China10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLegume Nitrogen Fixing Symbiosis · Agronomic Practices and Intercropping Systems · Soybean genetics and cultivation
Introduction
Nitrogen is a critical element for all organic life forms, and symbiotic nitrogen fixation contributes approximately 20 million tons annually.^1^ Most plants need to take up nitrogen from the soil, although more than 90% of the nitrogen in the soil is organic and cannot be directly absorbed and used by plants.^2^ Biological nitrogen fixation (BNF) is the most cost-effective and environmentally benign method of nitrogen fertilization, and high levels of nitrogen fixation increase legume productivity and contribute to sustainable agriculture.^3^ Among the various nitrogen fixation processes, symbiosis between rhizobia and legumes is considered the most effective BNF system, accounting for more than half of the global nitrogen fixation.^4^ The development of nodules in legumes is a highly intricate process that is synergistically regulated at both the physiological and molecular levels by plants and rhizobia. Studies have shown that BNF begins with flavonoid compounds secreted by legume roots, after which rhizobia are attracted to and stimulated to produce nodulation factors.^5^ These nodulation factors are sensed by receptor protein kinases (LysM RLK and LRR RLK) in the roots,^6^ initiating a symbiotic signaling cascade that leads to rhizobial infection and cortical cell division. Eventually, cortical cells divide to form rhizobial progenitors, which develop into mature rhizomes, which execute efficient nitrogen fixation^7^
Flavonoids are important signaling molecules in the symbiosis between rhizobia and legumes,^8^ and they attract rhizobacteria, synthesize nodulation factors, stimulate nodulation symbiosis signal transduction, participate in rhizobial organogenesis, and regulate rhizome formation.^9^ Flavonoids secreted by the root system of legumes are primarily found in soybeans (Glycine max), alfalfa (Medicago sativa), peas (Pisum sativum), and wood beans (Cajanus cajan).^10^ As common flavonoids secreted by the root systems of legumes, soy isoflavones have been demonstrated to possess physiological activities, including the induction of root nodule formation.^11^
Isoflavone biosynthesis begins with the deamination of phenylalanine, which is catalyzed by phenylalanine ammonia-lyase (PAL) to form cinnamic acid. Cinnamic acid is then hydroxylated by cinnamate-4-hydroxylase (C4H) to form p-coumaric acid, which is subsequently activated by 4-coumarate-CoA ligase (4CL) to produce p-coumaroyl-CoA, a common precursor in flavonoid biosynthesis.^12^ In the later stages of this pathway, p-coumaroyl-CoA is converted into isoliquiritigenin or naringenin chalcone by chalcone reductase (CHR) and chalcone synthase (CHS). These chalcones are then transformed by chalcone isomerase (CHI) into liquiritigenin or naringenin. Finally, isoflavone synthase (IFS) and 2-hydroxyisoflavone dehydratase (HID) catalyze the formation of isoflavonoid aglycones, such as daidzin, genistein, and glycitein.^13,14^ This biosynthetic pathway is predominantly found in legumes.^15^ The two principal isoflavonoids in soybeans are daidzein and genistein, which play pivotal roles in the formation of nodules and regulation of nodule ratios.^16^ Isoflavones are key molecules in symbiotic signaling between legumes and rhizobia that induce the production of nodulation factors (Nod factors) by rhizobia, which in turn initiates nodule formation.
CHI is the second rate-limiting and first reported enzyme in flavonoid biosynthesis. It catalyzes intramolecular cyclization reactions converting bicyclic chalcones into tricyclic (2S)-flavanones.^17^ Expression levels of CHI in plants have a crucial positive effect on flavonoid metabolism.^18^ Previous studies have divided the CHI family genes into four different types (types I-IV). Both type I and type II proteins are CHIs that present enzymatic activity to catalyze the stereospecific isomerization of chalcones to yield their corresponding flavanones.^19^ Notably, this cyclization reaction can also proceed spontaneously, although greater catalytic efficiency (107-fold) occurs with the aid of these CHIs. Type I CHIs are widely present in vascular plants and involved in the production of (2S)-naringenin (5,7,40 -trihydroxyflavanone) by exclusively isomerizing naringenin chalcone (4,20,40,60 -tetrahydroxychalcone). Type II CHIs appear to be specific to legumes and are primarily responsible for isoflavonoid production.^20^ Compared with these type I and II CHIs, type III and type IV CHIs are also found in plants but do not exhibit CHI activity due to the substitution of several catalytic core residues.^21^
The soybean CHI family also contains four subfamilies: Type I: GmCHI2; Type II: GmCHI1A, GmCHI1B1, GmCHI1B2; Type III: GmCHI3A1, GmCHI3A2, GmCHI3B1, GmCHI3B2, GmCHI3C1, GmCHI3C2; and Type IV: GmCHI4A, GmCHI4B.^22,23^ Type I CHIs is expressed predominantly in floral tissues, whereas Type II CHIs is root-specific and regulates enzymatic activity in isoflavonoid synthesis.^24^
Therefore, in this study, we investigated the effects of type II CHIs on soy isoflavone content and nodulation. Composite plants transformed with GmCHI1s using a soybean hair-rooting technology system were obtained, the main functional genes were screened, and the role of GmCHI1s in the symbiotic nitrogen fixation process of soybean rhizobia was elucidated.
Materials and Methods
Plant Materials and Plant Growth Conditions
2.1.
Soybean cultivar Williams 82 was grown in an artificial climate chamber at Jilin University, Changchun, China, under a 16 h light (25°C)/8 h (22°C) diurnal cycle with 75% relative humidity. The roots, stems, leaves, flowers, and mature seeds were collected for cloning and expression analysis of GmCHI1s (GmCHI1A, GmCHI1B1 and GmCHI1B2). All collected samples were immediately plunged in liquid N_2_ and then were stored at − 80°C prior to RNA isolation. Three independent biological replicates (at least ten replicates) were used for RNA extraction.
Cloning of GmCHI1s Genes
2.2.
TRIzol reagent was used to extract root RNA from Williams 82. RNA solution was reverse transcribed into cDNA using a PrimeScript™ II 1st Strand cDNA Synthesis Kit (Takara, Changchun, China)^25^. The open reading frames (ORFs) of GmCHI1s were amplified from root cDNA using primers (Table S1). Polymerase chain reaction (PCR) was performed as follows: 94°C for 8 min, followed by 30 cycles of 94°C for 30 s, 50/58°C for 30 s, and 72°C for 1 min followed by a final extension at 72°C for 8 min. The amplified fragments were cloned into the pMD-18T cloning vector (TaKaRa, Changchun, China) and sequenced for confirmation (Kumei Technology Co., Ltd., China).
Bioinformatics Analysis of the GmCHI1s Gene Sequences
2.3.
Gene sequences of GmCHI1s were searched and compared using the NCBI database (https://www.ncbi.nlm.nih.gov/). GmCHI1s protein sequences were analyzed using the ProtParam software (http://web.expasy.org/protparam/). The amino acid sequences of GmCHI1s genes were compared and analyzed using the DNAMAN software. A phylogenetic tree was constructed using the neighbor-joining method in MEGA6.0 software. Bootstrapping values were presented from 1000 repeated calculations, and bootstrap values of > 70% were recommended.
Construction of Hairy Root Expression Vector
2.4.
Using the double-enzyme digestion method, the GmCHI1s genes were recombined to construct an overexpression vector (pCHF1301) and interference vector (pFGC5941). The overexpression vector pCHF1301 was digested with the restriction endonucleases bamH1 and Pst1, and the interference vector pFGC5941 was digested with the restriction endonucleases Nco1 and Asc1. Specific primers were designed, and the corresponding cleavage junction was inserted at the 3’ end of the primers, and the plasmid pMD18-T-GmCHI1s containing the complete coding sequence of the GmCHI1s genes was used as a template for PCR amplification. The product was purified, recovered, ligated with the pMD18-T vector, and sequenced for confirmation (Kumei Technology Co. Ltd., China). The correct fragment was ligated into the appropriate vector using T4 ligase. The recombinant plasmid was then transformed into Escherichia coli (DH5α) and verified by colony PCR. The correct recombinant plasmids pCHF1301-GmCHI1s and pFGC5941-GmCHI1A were transformed into the Agrobacterium rhizogenes strain K599 using the freeze-thaw method (Weigel & Glazebrook, 2006).^26^
GmCHI1s Transformation into Composite Plants
2.5.
Williams 82 seeds were grown in pre-sterilized wet vermiculite at a depth of 5 cm for germination, 7 day old soybean seedlings were injected at the cotyledonary node locus with A. rhizogenes (K599) medium with a recombinant expression vector,^27^ and a null control was established. Hairy roots appeared after two weeks and were transplanted after subtracting the primary roots. The slow-growing rhizobium Bradyrhizobium japonicum USDA110 was also cultured at 28°C in Yeast Mannitol Agar (YMA) medium. Approximately 50 mL of a suspension with an optical density of approximately 0.05 to 0.1 at 600 nm (OD_600_) adjusted to 0.08 to 0.1 was applied to each plant.
Phenotyping of Transgenic Plants
2.6.
Four weeks after rhizobial application, the phenotypic data of hairy roots, nodules, and leaves were collected. The Soil Plant Analysis Development (SPAD) chlorophyll meter and nitrogen content (in mg/g) of the leaves of the soybean composite plants were determined, and the number of hairy roots, length of roots, weight of roots, number of rhizomes, weight of rhizomes, and volume of rhizomes of the composite plant roots were measured. Eight seedlings were analyzed for each transformation vector and control.
Isoflavones Extraction and High Performance Liquid Chromatography (HPLC) Analysis
2.7.
The fresh weight of soybean composited plant transgenic root samples to approximately 0.5 g was weighed, 80% methanol (analytical purity) extract was added to a mortar and the sample was grinded to a homogenate. Then, the sample was transferred to a 10 mL volumetric flask to set the volume. The sample was ultrasonicated at 50°C (frequency 40 kHz, power 300 W) to assist in extraction for 40 min. Every 10 min, the sample will be taken out of the sample up and down the upside down mixing. Ultrasonic centrifugation was then performed at 12,000 rpm and room temperature for 10 min. After sonication, the supernatant was removed, passed through a 0.45-μm filter membrane to obtain the solution to be tested, and stored at 4°C for testing.
The solvents and distilled water were of HPLC grade and degassed before use. A Phenomenex C18 column (Shimadzu LC-20A, Japan, 150 mm × 4.6 mm, 5.0 μm) was utilized for HPLC. The sample solution (40 µL) was aspirated, and the isoflavone content of each component was determined using liquid chromatography, with three replicates performed for each sample.
Quantitative RT-PCR (qRT-PCR) Analysis of Gene Expression
2.8.
The GmCHI1s gene expression patterns in various tissues were examined using qRT-PCR. The primers used for qRT-PCR are listed in Table S1. For analysis of tissue-specific expression, total RNA was isolated from different organs (seeds, roots, stems, leaves, and flowers) and transgenic hairy roots using RNA plant Plus Reagent (Tiangen, Changchun, China) according to the manufacturer’s protocol. cDNA was reverse transcribed using M-MLV reverse transcriptase (TaKaRa, Changchun, China). The qRT-PCR was performed using SYBR Green I dye (TaKaRa, Changchun, China) on a real-time PCR machine (Applied Biosystems 7500, Foster City, USA). Soybean GmActin was used as an internal control.
Statistical Analysis
2.9.
Samples were subjected to statistical analysis using IBM SPSS19.0 and GraphPad Prism 8.0 software. Data were presented as the mean ± SD. Statistical significance among the treatments was evaluated using one-way analysis of variance (ANOVA) and Student’s t-test. Asterisks or different lowercase letters indicate significant differences according to p < .05 or p < .01.
Result
Sequence and Expression Analysis of GmCHI1s in Soybean
3.1.
GmCHI1A, GmCHI1B1, and GmCHI1B2 were cloned by PCR using soybean Williams 82 cDNA as a template. Bioinformatic analysis showed that GmCHI1A, GmCHI1B1, and GmCHI1B2 were the most recently identified type II GmCHIs (see Figure 1a). GmCHI1A contained an entire ORF of 657 bp that encodes 218 amino acids. The ORFs of GmCHI1B1 and GmCHI1B2 both contained 681 bp, which encoded 226 amino acids. Conserved structural domains of these three genes showed 87.32% homology. GmCHI1B1 and GmCHI1B2 had higher homology in the conserved protein domain, with a value of 94.46%. All three genes retained all residues of the catalytically active sites of the CHI protein, which are related to the catalytic ability of trihydroxychalcone and tetrahydroxychalcone (see Figure 1b). Figure 1.Sequence and expression analysis of GmCHI1s. (a) A phylogenetic tree containing GmChI1s and 14 other species of CHI proteins was obtained using the neighbor joining method of the MEGA software. (b) Comparison of the protein sequences of GmChI1s using DNAMAN software. (c) The expression of GmChI1s in roots, stems, leaves, flowers and mature seeds.
qRT-PCR was used to detect the expression levels of GmCHI1s in the roots, stems, leaves, flowers, and seeds. The results showed that the expression of these three genes was observed in all tested samples. However, their expression patterns differed among organs. The highest level of GmCHI1A gene was observed in the roots, which was two times the expression in stems, flowers and seeds, and the lowest expression was observed in the leaves; The highest level of GmCHI1B1 gene was detected in the seeds, which was 1.5 times the expression in the roots and 3 times of the expression in the flowers, and the lowest expression was observed in the stems and leaves; The highest level of GmCHI1B2 expression was observed in the flowers, which was 2.5 times higher than that in the roots and 7 times higher than that in the stems. The expression of GmCHI1A was higher in the roots than in the other organs, implying that it was a possible that GmCHI1A works mainly in roots (see Figure 1c).
Overexpression of GmCHI1s in Soybean Resulted in a Differential Increase in Root Nodulation
3.2.
To determine whether the overexpression of GmCHI1s affects nodulation in soybeans, we generated transgenic plants overexpressing GmCHI1s using the soybean hairy rooting technique (see Figure 2a). The control consisted of soybeans transformed with an pCHF-1301 empty vector (EV1). The phenotypic characteristics of the composite plants were investigated 30 d after rhizobial infection. The aboveground leaf and belowground root growth statuses of overexpression plants were better than those of the control group. The results showed that overexpressing GmCHI1s affected soybean nodulation and growth to varying degrees (see Figure 2c). The nodule number, weight, and volume significantly increased in the roots of the overexpressing GmCHI1A (OE1A) plants. Simultaneously, the SPAD value and nitrogen content in the leaves hair root length, hair root weight, and hair root number also increased significantly. Plants overexpressing GmCHI1B1 (OE1B1) showed significantly increased nodule weight, nodule volume, hair root length, hair root weight, and hair root number (see Figure 2b). Plants overexpressing GmCHI1B2 (OE1B2) did not show significantly effects any phenotypic characteristics. Plants overexpressing GmCHI1A (OE1A) were superior to those overexpressing GmCHI1B1 (OE1B1) in terms of various indices. This suggests that GmCHI1A may be a pivotal gene affecting nodulation in soybeans with type II CHIs (see Figure 2). Figure 2.The effect of overexpression of GmCHI1s genes on root nodulation formation in soybean. (a) PCR test results of GUS gene and GUS staining results of the overexpressing GmChI1s roots (P: positive plasmid; W: negative control; 1: empty pCHF-gus hairy root; 2~4: experimental group pCHF-CHI1s hairy root); (b) phenotypic data statistics of comparing between overexpressing transgenic composite plants and control (** and *indicate significance at p ≤ .01 and p ≤ .05 respectively). (c) Phenotypic map of soybean overexpressing transgenic composite plants and control.
RNA Interference (RNAi) of GmCHI1s Genes Disrupts Nodulation and Growth in Soybean
3.3.
To further confirm the effect of GmCHI1A, an interference vector for GmCHI1A was constructed, and composite plants interfering with GmCHI1A (Ri1A) were obtained using the soybean hair-rooting technique. The control group was transfected with the empty vector, pCHF-1301 (EV2). The expression of GmCHI1B1 and GmCHI1B2 did not show a compensatory mechanism after interference with GmCHI1A; however, both showed a decreasing trend (see Figure 3a). The aboveground portion of Ri1A plants exhibited indications of stunting to some extent, as evidenced by the smaller leaf area, yellowish leaf color, and early leaf abscission (see Figure 3b). The phenotypic data showed that the aboveground biomass (e.g., chlorophyll SPAD value and nitrogen content) and hairy root length of Ri1A showed a decreasing trend compared with the control, although the difference was not significant. The number of hairy roots, weight of hairy roots, number of rhizomes, weight of rhizomes, and volume of rhizomes were significantly lower in Ri1A plants than the control (see Figure 3c). This indicated that interference with the GmCHI1A gene affected the formation of soybean nodules and had a certain effect on growth and development (see Figure 3). Figure 3.The effect of interfering GmCHI1A gene on root nodulation formation in soybean. (a) Relative expression of GmChI1s in transformed hairy roots of interfering GmCHI1A. (b) phenotype of the transformed hairy roots of interfering GmCHI1A. (c) phenotypic comparison between transgenic composited plants with interfering GmCHI1A and control (** and *indicate significance at p ≤ .01 and p ≤ .05 respectively).
GmCHI1A Influences the Expression Levels of Genes in the Soybean Nodulation Signaling Pathway
3.4.
The expression of relevant nodulation pathway genes was detected using qRT-PCR. The results showed that the expression levels of relevant nodulation signaling pathway genes, especially Early Nodulin 40–1(ENOD40–1), Glycine max Nodule factor receptor 5α (GmNFR5α), Glycine max Nodule factor receptor 1α (GmNFR1α), microRNA172c precursor (pre-miR172c), Glycine max Nodulation signaling pathway 1 (GmNSP1), and Glycine max Nodulation signaling pathway 2 (GmNSP2), were significantly higher in OE1A compared with the control, although no significant difference was observed in the expression of GmNF1α (see Figure 4a). The expression of relevant nodulation signaling pathway genes, including ENOD40–1, GmNF5α, pre-miR172C, GmNSP1, and GmNF1α, was significantly lower in Ri1A than in the control group, whereas the expression of GmNSP2 was significantly higher in Ri1A (see Figure 4b). Therefore, except for GmNF1α and GmNSP2, all detected genes on the nodulation signaling pathway were co-expressed with GmCHI1A. This suggests that GmCHI1A positively regulates the expression of genes related to the nodulation signal transduction pathway, which coincides with the rhizome development and root isoflavone content of the transgenic plants, as previously investigated (see Figure 4). Figure 4.Alterations in GmCHI1A expression affect the transcript levels of nodule-specific genes. (a) Relative expression levels of related genes in transgenic roots overexpressing GmCHI1A. (b) relative expression levels of related genes in transgenic roots interfering GmCHI1A. data are means±sd. Asterisks above the columns represent statistically significant differences (*p < .05, **p < .01; Duncan’s test).
GmCHI1s Regulate Soybean Isoflavone Accumulation
3.5.
The isoflavone content of the soybean roots was determined using HPLC. The data showed that overexpression of each GmCHI1s gene had a different effect on the isoflavone content of soybean roots. The daidzin, genistin, daidzein, genistein, and glycitein contents as well as the total isoflavone content were significantly increased in OE1A compared to the control. This suggests that overexpression of the GmCHI1A gene in the root can positively regulate isoflavone components. Compared with the control, OE1B1 showed a highly significant increase in glycitein content but a decrease in the rest of the components and total isoflavone content, which may be due to competition for synthesis substrates among the isoflavone components (see Figure 5a). This suggested that GmCHI1B1 plays a critical role in the synthesis of glycitein. After the interference of GmCHI1A, Ri1A showed significant downregulation of glycitein content and a nonsignificant reduction of daidzein, genistein, and total isoflavone contents. The daidzin content of Ri1A showed a certain degree of backfilling (see Figure 5b). Taken together, these results showed that GmCHI1A had a positive regulatory effect on all isoflavone fractions and total isoflavone content, GmCHIB1 positively regulated soybean flavin synthesis but did not affect total soybean isoflavone content, and GmCHI1B2 had some effect on isoflavone content (see Figure 5). Figure 5.Analysis of isoflavone content and relative expression of related genes in interference and overexpression composite plants. (a, b) quantitative determination of the genistein, genistin, daidztein, daidztin, glycitein content using HPLC methods in overexpression and RNAi transgenic hairy roots. Total isoflavone content is equal to the sum of genistein, genistin, daidztein, daidztin and glycitein contents. Data are means±sd. Asterisks above the columns represent statistically significant differences. (c) qRT-PCR detected the expression level of GmCHS8, GmIFS1, GmHID in overexpression GmCHI1A and RNAi transgenic hairy roots of composite plants respectively. (*p < .05, **p < .01; Duncan’s test).
GmCHI1A Regulates Isoflavone Biosynthetic Gene Expression in the Roots
3.6.
Given that OE1A demonstrated a notable elevation in isoflavone content relative to the control, an investigation was conducted to ascertain whether the expression of genes associated with soybean isoflavone synthesis had undergone alterations in the transgenic roots. To this end, the expression of isoflavone biosynthesis genes, including Glycine max Isoflavone synthase 1 (GmIFS1), Glycine max Chalcone synthase 8 (GmCHS8), and Glycine max Hydroxyisoflavanone dehydratase (GmHID), was evaluated by qRT-PCR. The expression of GmIFS1, GmCHS8, and GmHID was markedly elevated in OE1A cells relative to that in EV1 cells (see Figure 5c). This finding suggests that the overexpression of GmCHI1A in the root lines led to the upregulation of isoflavone synthesis-related genes, thereby increasing the isoflavone content of the root lines. In contrast, the expression of GmIFS1, GmCHS8, and GmHID was significantly reduced in the transgenic hairy roots with interfering GmCHI1A (Ri1A) compared to the null control (EV1) (see Figure 5c), suggesting that interfering with GmCHI1A genes in the root system exerted an inhibitory effect on the genes related to isoflavone synthesis. Changes in the expression of isoflavone synthesis-related genes in the transgenic plants coincided with changes in isoflavone content (see Figure 5).
Discussion
BNF by legumes and rhizobia is an efficient, nonpolluting, and sustainable method for plants to obtain nitrogen.^28^ Isoflavones secreted by soybean roots can stimulate signal transduction in symbiotic nodules. This study was focused on CHI, a key enzyme in isoflavone synthesis, to investigate the effect of CHIs on isoflavone synthesis and nodule development in soybean.
Isoflavone synthesis is regulated by the phenylpropane metabolic pathway, in which CHI is involved in the synthesis of intermediates. Isoliquiritigenin and naringenin chalcones are catalytically synthesized by CHI into flavanones (liquiritigenin and naringenin). Liquiritigenin and naringenin are important prerequisites for the synthesis of other metabolites, with liquiritigenin representing a substrate for the biosynthesis of glycitein and daidzein and naringenin representing a substrate for the synthesis of genistein and various anthocyanins.^29^ CHI has been studied extensively and its mutants in aster (Callistephus chinensis) and carnation (Dianthus caryophyllus) lead to yellowish flower petals, while its mutant in Arabidopsis has leads to a change in seed coat color.^30^ Overexpression of CHI in Scutellaria baicalensis, Glycyrrhiza uralensis, and Arachis hypogaea leads to increased production of flavonoids in hairy roots.^31,32^ In soybean, there are three CHI1 genes, GmCHI1A, GmCHI1B1, and GmCHI1B2. Expression analysis showed that these three genes were biased in different organs. GmCHI1A was expressed at the highest levels in the roots, while GmCHI1B1 and GmCHI1B2 were expressed at the highest levels in the seeds and flowers, respectively. We investigated the function of these three genes in the roots and found that GmCHI1A promoted the accumulation of all isoflavone components while GmCHI1B1 promoted the accumulation of glycitein. These results suggest that GmCHI1s regulate isoflavone synthesis in soybeans and have evolved to be functionally differentiated.
The nodulation of legume roots is a process in which the host plant interacts with rhizobacteria and involves a complex series of regulatory networks that incorporate many genes. In M. truncatula, knockdown of MtENOD40–1 expression resulted in a 50% reduction in nodule number, suggesting that ENOD40 is involved in nodule initiation.^33^ Research has confirmed that NSP1 and NSP2 interact to form a complex, which is enhanced by Nod factor perception and necessary for the proper development of nodules.^34^ NF1α and NF5α are two highly related lipo-oligochitin LysM type receptor kinase genes that are presumed to be critical nodulation inducing (Nod) factor receptors.^35,36^ Additionally, miR172c has been shown to play an essential role in the control of nodulation through the GmNINa–miR172c–GmNNC1 regulatory network.^37^ In this study, ENOD40–1, NF5α, miR172c, NSP1, and NSP2 were significantly increased after overexpression of GmCHI1A while ENOD40–1, NF1α, NF5α, miR172c, and NSP1 were significantly decreased after interference with GmCHI1A. These results are consistent with those of previous studies. This suggests that GmCHI1A is involved in nodule formation and affects the intensity of nodule signaling.
Isoflavonoids are released from plant roots and act as signaling molecules, activating nod operons in compatible bacteria, which induce nodule formation and nitrogen fixation.^38^ Isoflavones contain 12 different isomers in four structural forms (aglycone, glucoside, 6″-O-acetylglucosides, and 6″-O-malonylglucosides). Daidzein, glycitein, and genistein belong to the aglycone type.^39^ In particular, the symbiotic interaction between Phaseolus vulgaris roots and Rhizobium leguminosarum is regulated by genistein, daidzein, and coumestrol.^40^ In soybeans, the main isoflavonoids perceived by rhizobacteria are daidzein and genistein.^41^ Most isoflavonoids in root secretions early in the growth of soybean plants are soybean daidzein derivatives.^16^ Ramongolalaina suggested that genistein is involved in controlling soybean nodule formation through a QTL analysis of genetic interrelationships between soybean nodule interactions and root-secreted genistein.^42^ In this study, the nodule number and daidzein and genistein content increased in composite plants overexpressing GmCHI1A and decreased in composite plants interfering with GmCHI1A, this suggests that GmCHI1A may influence nodule formation by regulating daidzein and genistein synthesis.
Composite plants overexpressing GmCHI1B1 also showed significant increases in nodule number, weight, and volume. Unexpectedly, only the glycitein content significantly increased in overexpressing GmCHI11B1 roots, whereas all other isoflavonoid fractions decreased. Previous studies have shown that glycitein belongs to the same type of aglycone structure as genistein and daidzein. Therefore, glycitein might have functions similar to those of genistein and daidzein. However, glycitein has only been studied for disease resistance and antioxidant purposes^43^ and rarely for nodule development. The results of this study showed that GmCHI1B1 affected nodule formation by regulating glycitein content. Therefore, whether glycitein can replace genistein and daidzein to promote plant nodule development should be determined.
Nitrogen fixation in soybeans not only provides nutrients to crops, but also contributes to the improvement of crop ecosystems and may provide a feasible method of resolving the multiple societal crises of population, food, energy, and environment. It is of great significance to analyze the molecular mechanism of the soybean-rhizobium symbiosis system, which can aid in the selection of soybean varieties and achieve fertilization requirements and environmental protection. The results of this study showed that GmCHI1A and GmCHI1B1 could be used as genes to enhance nitrogen fixation in soybeans and thus could be applied to improve soybean varieties.
Conclusion
In this study, GmCHI1A was expressed at the highest level in soybean roots, and its overexpression in soybean significantly promoted nodule development. However, interference with GmCHI1A in soybean reduced nodules. This demonstrates that GmCHI1A is the main effector gene that regulates nodule formation in type II GmCHIs. Overexpression of GmCHI1A increased the content of all the isoflavone fractions. However, overexpression of GmCHI1B1 increased the glycitein content, which also promoted nodule development. Therefore, this study is the first to propose that glycitein may affect soybean nodulation.
This study revealed the mechanism by which type II GmCHIs affect nodulation in soybeans and screened the main functional genes, providing a theoretical basis for subsequent research on BNF. Glycitein, an isoflavone that affects nodulation in soybeans, was discovered for the first time, providing new ideas for the subsequent enrichment of soybean nodulation signaling pathways.
Supplementary Material
Supplemental Material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Yuan S, Peng S. Exploring the trends in nitrogen input and nitrogen use efficiency for agricultural sustainability. Sustainability. 2017;9(10):1905. 10.3390/su 9101905. · doi ↗
- 2Han H, Tian Z, Fan Y, Cui Y, Cai J, Jiang D, Cao W, Dai T. Water-deficit treatment followed by re-watering stimulates seminal root growth associated with hormone balance and photosynthesis in wheat (Triticum aestivum L.) seedlings. Plant Growth Regul. 2015;77(2):201–10. doi: 10.1007/s 10725-015-0053-y. · doi ↗
- 3Schulte CCM, Borah K, Wheatley RM, Terpolilli JJ, Saalbach G, Crang N, De Groot DH, Ratcliffe RG, Kruger NJ, Papachristodoulou A, et al. Metabolic control of nitrogen fixation in rhizobium-legume symbioses. Sci Adv. 2021;7(31):eabh 2433. doi: 10.1126/sciadv.abh 2433.34330708 PMC 8324050 · doi ↗ · pubmed ↗
- 4Wang ET. Symbiosis Between Rhizobia and Legumes. In: Wang ET, Tian CF, Chen WF, Young JPW Chen WX. Ecology and Evolution of Rhizobia. Springer: Singapore; 2019. p. 3–19. doi:10.1007/978-981-32-9555-1_1. · doi ↗
- 5Xu P, Wang E. Diversity and regulation of symbiotic nitrogen fixation in plants. Curr Biol. 2023;33(11):R 543–59. doi: 10.1016/j.cub.2023.04.053.37279688 · doi ↗ · pubmed ↗
- 6Buendia L, Girardin A, Wang TM, Cottret L, Lefebvre B. Lys M Receptor-Like Kinase and Lys M Receptor-Like Protein Families: An Update on Phylogeny and Functional Characterization. Front Plant Sci. 2018;9:1531. doi: 10.3389/fpls.2018.01531.30405668 PMC 6207691 · doi ↗ · pubmed ↗
- 7Ferguson BJ, Indrasumunar A, Hayashi S, Lin M, Lin Y, Reid DE, Gresshoff PM. Molecular analysis of legume nodule development and autoregulation. J Integr Plant Biol. 2010;52(1):61–76. doi: 10.1111/j.1744-7909.2010.00899.x.20074141 · doi ↗ · pubmed ↗
- 8Sugiyama A. The soybean rhizosphere: metabolites, microbes, and beyond—A review. J Adv Res. 2019;19:67–73. doi: 10.1016/j.jare.2019.03.005.31341671 PMC 6630087 · doi ↗ · pubmed ↗