Cocaine- and amphetamine-regulated transcript (CART) peptide-positive neuron populations in the enteric nervous system of the porcine descending colon depend on age and gender
Krystyna Makowska, Kainã R. C. Fagundes, Renata de Britto Mari, Sławomir Gonkowski

TL;DR
This study shows that the number of CART-positive neurons in the gut's nervous system varies with age and gender in pigs.
Contribution
The study reveals age- and gender-dependent changes in CART-positive neurons in the porcine colonic enteric nervous system.
Findings
Age significantly affects the number of CART-positive neurons in the myenteric plexus of the porcine colon.
Sex differences in CART-positive neuron populations are more pronounced in adult pigs.
CART-positive neurons may be involved in neuroplasticity processes influenced by sex hormones.
Abstract
The enteric nervous system (ENS) is a complex structure located in the wall of the gastrointestinal tract. One of the less-known active substances found in the ENS is cocaine- and amphetamine-regulated transcript peptide (CART). It is known that CART-positive enteric neurons take part in the reactions to pathological stimuli, but knowledge of physiological stimuli-dependent changes in their population is extremely limited. The aim of the present study was to investigate the age- and gender-dependent diversities in the distribution of CART-positive neurons in the porcine colonic ENS using the double immunofluorescence technique. The obtained results have shown that age affects the number of CART-positive neurons in the colonic ENS and the character and intensity of age-caused changes depend on the type of the enteric plexus, and the most visible changes have been noted in the myenteric…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
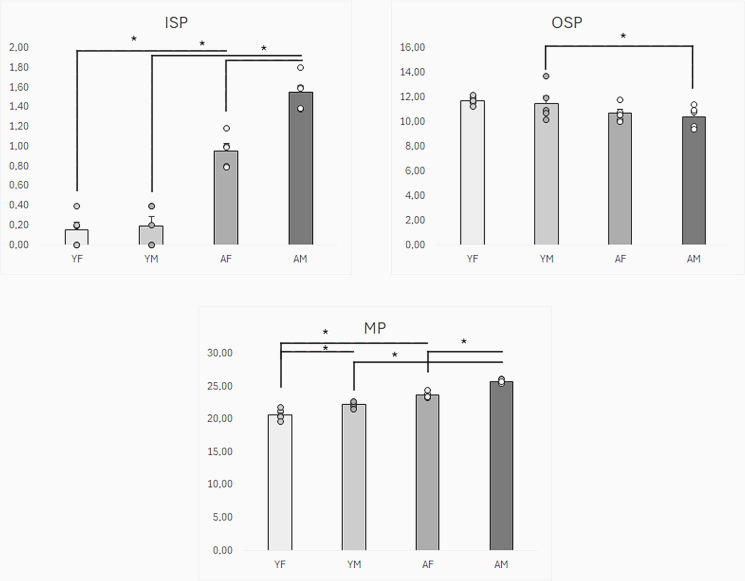 Fig 1
Fig 1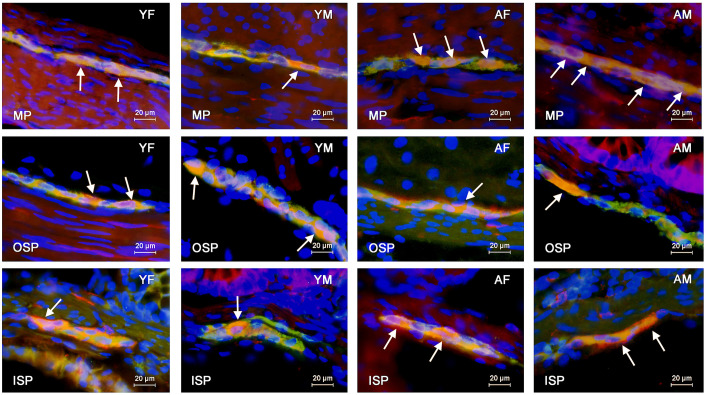 Fig 2
Fig 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGastrointestinal motility and disorders · Congenital gastrointestinal and neural anomalies · Neuropeptides and Animal Physiology
Introduction
The enteric nervous system (ENS) which is situated in the wall of the gastrointestinal tract. takes part in the regulation of many gastric and intestinal functions [1,2]. Due to the millions of neurons constituting the ENS and its high degree of independence from the central nervous system (CNS), it is often called the “second” or “gut brain” [3–5].
The enteric neurons form intramural ganglia and are often interconnected with each other by a dense network of fibres and create ganglionated plexuses [6–10]. In the large intestine of the domestic pig, the ENS is built of three types of enteric plexuses: 1) myenteric plexus (MP) located between circular and longitudinal muscular fibres; 2) outer submucous plexus (OSP) – located in the submucosal layer, laid over the circular muscle fibres and 3) inner submucous plexus (ISP) – located in the submucosal layer, beneath the mucosal layer.
The enteric neurons are characterised by high morphological, functional and neurochemical diversity [11]. Till now, several dozens of substances acting as neurotransmitters, neuromodulators, enzymes or transporters have been described in the enteric neurons [12–14]. One of these substances is cocaine- and amphetamine-regulated transcript (CART) peptide [11].
CART peptide occurs in both the central and peripheral nervous system of various mammal species, including humans, and it is considered to be one of the most important factors involved in the regulation of food intake and anorexigenic actions [14–16]. Moreover, CART plays a part in the development of the nervous system and has neuroprotective properties [17,18]. CART has also been identified in the enteric nervous system in all the enteric plexuses in various segments of the gastrointestinal tract of a diversity of mammal species [14]. CART in the intestine is probably involved in the regulation of peristalsis and secretory activity, but not through a direct effect on the intestinal wall but through the CNS [19]. Locally, the CART-positive enteric neurons are also involved in the plastic modulations of the enteric neurons under the impact of physiological and pathological stimuli [20,21]. However, the exact roles of CART in the intestine still remain unexplained.
The enteric neurons can respond to a wide range of physiological and pathological factors [22–24]. These changes concern the structure and functions of neurons but primarily manifest themselves in a change in their neurochemical characteristics and fluctuation in intraneuronal synthesis, transport and secretion of active factors [24–26]. The majority of studies focus on the changes in the ENS under pathological stimuli, including gastrointestinal and systemic diseases, inflammatory processes and toxic substances in the food [21,22,27–30]. The ENS can also change under physiological factors, including age, gender or diet, but the knowledge about these issues is much more limited [31–35].
Previous results have described changes in the number of enteric neurons containing CART exposed to diverse pathological factors, which strongly suggests that CART–positive neurons in the gastrointestinal tract take part in adaptive and/or protective processes [21,36–38]. On the other hand, information about the participation of CART–positive enteric neurons in adaptive processes during aging is extremely scanty and is limited to only one study, which describes age-dependent changes in the degree of co-localisation of calcitonin gene-related peptide and CART in the same enteric neurons [39]. However, to date, the influence of age and/or gender on the percentage of enteric neurons containing CART has not been studied at all.
It should be pointed out that the choice of the animal species and segment of the digestive tract was not random. It is relatively well known that the ENS in the domestic pig shows great similarities to human ENS in terms of organisation, biochemistry and neurochemical characterisation of neurons [40,41]. Therefore, physiological and pathological processes and neuroplasticity reactions in the enteric neurons are similar in both these mammal species, and the domestic pig is considered to be an optimal animal model for studying the human ENS [42,43]. In turn, the descending colon is a segment of the digestive tract where many diseases can develop. In many pathological processes occurring in the descending colon, e.g., Crohn’s disease and cancer, changes in the ENS have been noted [44–46], and the risk of these processes often depends both on the age and gender of individuals [29].
Given the above, the aim of the present study was to investigate for the first time the correlation between age and gender and the number of neurons containing CART in the ENS localised in the wall of the descending colon of the domestic pig. The results obtained in the present work may contribute to a better understanding of the roles of CART in neuroplasticity processes occurring during the development and aging of the ENS.
Materials and methods
The investigation was performed on the tissues collected from domestic pigs slaughtered at commercial slaughterhouses using a typical method of slaughter (carbon dioxide). All experimental protocols were approved by the Local Ethical Committee for Animal Experiments in Olsztyn working at the University of Warmia and Mazury in Olsztyn, according to Act for the Protection of Animals for Scientific or Educational Purposes of 15 January 2015 (Official Gazette 2015, No. 266), applicable in the Republic of Poland (based on the consent No 28/2013 of 22 May 2013 and 65/2013/DLZ of 27 Nov 2013). During this experiment, all methods were carried out in accordance with relevant guidelines and European and Polish regulations. Moreover, the study was carried out in compliance with the ARRIVE guidelines.
The fragments of the descending colon with a length of about 2 cm and located about 30 cm before the anus were collected from 20 animals divided into four groups (n = 5 in each group): 1) YF group – young female pigs before puberty (about 10 weeks old); 2) AF group – adult female pigs after puberty (7–8 months old); 3) YM group – young male pigs before puberty (about 10 weeks old) and 4) AM – adult male pigs after puberty (7–8 months old). Male pigs were castrated at the age of about one week.
The intestinal fragments were collected immediately after the death of the animals during bowel removal in post-slaughter processing. Just after sampling, tissues were put into a solution of 4% buffered paraformaldehyde (pH 7.4) for 1 h to perform immersive fixation. The fragments of the intestine were then rinsed in a phosphate buffer (0.1 M, pH 7.4) for three days at 5 °C with a daily exchange of the buffer. After this period, tissues were put in an 18% phosphate-buffered sucrose solution and stored at 5 °C for at least three weeks until further study. After this period, the fragments of the colon were frozen at − 22 °C, cut perpendicular to the intestinal lumen into 14-µm-thick sections with a microtome (Microm, HM 525, Walldorf, Germany) and mounted on the microscopic slides.
Intestinal sections were subjected to the typical double immunofluorescence method described previously by Makowska and Gonkowski [39]. In brief, the method consisted of the following stages: 1) drying the sections at room temperature for 45 min; 2) incubation with “blocking” solution (10% goat serum, 0.1% bovine serum albumin (BSA), 0.01% NaN_3_, Triton X-100, and thimerosal in PBS) for 1 h to prevent non-specific staining; 3) overnight incubation with the mixture of two antibodies: mouse antibody directed against protein gene product 9.5 (PGP 9.5, used as a pan-neuronal marker obtained from Biogenesis Ltd., Poole, UK, (working dilution 1:1000) and rabbit antibody against CART peptide purchased from Phoenix Pharmaceuticals, Inc., Burlingame, CA, USA (working dilution 1:16000); 4) one-hour incubation with a mixture of species-specific secondary antisera: donkey anti-mouse IgG conjugated with Alexa fluor 488 and donkey anti-rabbit IgG conjugated with Alexa fluor 546 (both from Invitrogen, Carlsbad, CA, USA, both in a working dilution of 1:1000); 5) covering the sections with buffered glycerol and coverslips. The information about antibodies used I the present study is summarized in Table 1.
Table 1: The list of primary and secondary antibodies used in this study.
All of the above-mentioned stages were performed at room temperature. Stages no. 2,3, and 4 were performed in a humid chamber. Between each stage of labelling, colon sections were rinsed in PBS for 30 min with a PBS change every 10 min. Moreover 4′,6-diamidino-2-phenylindole (DAPI) obtained from Sigma obtained from Sigma-Aldirch (St Louis, MO, USA) in working dilution 1 µ /ml was used to visualise cell nuclei.
Checking the specificity of the staining performed consisted of typical testing, including 1) pre-absorption of antibodies with appropriate antigens: PGP 9.5 (AbD Serotec, Kidlington, UK) or CART (Phoenix Pharmaceuticals, Burlingame, CA, USA) at a concentration of 20 μg/mL, overnight at room temperature; 2) omission test and 3) replacement of primary antibodies by non–immune sera. Negative staining was obtained during the pre-absorption of antibodies after applying the above tests.
Intestinal fragments were analysed using an immunofluorescence microscope BX51 (Olympus, Tokyo, Japan) equipped with appropriate filters. To establish the percentage of enteric neurons containing CART, at least 500 cells immunoreactive to PGP 9.5 located in each type of enteric ganglia were evaluated for the presence of CART in each animal included in the study. The results were presented in percentages, and the number of analysed PGP 9.5-positive cells was considered 100%. To prevent double counting the same neurons, sections of the descending colon included in the analysis were located at least 200 μm from each other. The obtained results were presented as mean values ± SEM. A statistical analysis was performed with a T-Student test using GraphPad Prism version 9.2.0 (GraphPad Software, San Diego, California, USA). Differences were considered statistically significant at P < 0.05.
Results
Neurons containing CART were found in all types of colonic enteric plexuses in all groups of animals included in the study.
The largest percentage of CART-positive neurons was observed in the MP. In young female pigs, the percentage of such neurons amounted to 22.3 ± 0.2% of all cells immunoreactive to PGP 9.5 and was statistically significantly higher than the value noted in young male pigs, where the percentage of CART-positive cells achieved the level of 20.7 ± 0.4%. Moreover, the number of CART-like immunoreactive neurons in the MP increased with the growth and maturation of the organism, and this increase was more visible in male pigs. In adult pigs, the percentage of neurons containing CART amounted to 23.7 ± 0.2% and 25.8 ± 0.1% in females and males, respectively. These values were statistically significantly higher than these values in young animals of the appropriate gender. Moreover, contrary to young animals, the percentage of CART-positive cells noted in adult males was statistically significantly higher than that noted in females (Fig 1, Fig 2 Table 2).
Table 2: The percentage (mean ± SEM) of enteric neurons containing CART in the porcine descending colon under physiological conditions in young (10-week-old) females and males and adult (7–8-month-old) females and males.
Graphical comparison of the number of nerve cells immunoreactive to protein gene product (PGP 9.5) – used here as a pan-neuronal marker and cocaine- and amphetamine-regulated peptide (CART) in inner submucous (ISP), outer submucous (OSP) and myenteric plexus (MP) of porcine colon between young females (YF), young males (YM), adult females (AF) and adult males (AM).Statistically significant differences between groups are marked with * .
Distribution pattern of nerve cells immunoreactive to protein gene-product 9.5 (PGP 9.5) – used as a pan-neuronal marker – and cocaine- and amphetamine-regulated peptide (CART) in the myenteric (MP), outer submucous (OSP) and inner submucous (ISP) plexuses of porcine descending colon in young females (YF), young males (YM), adult females (AF) and adult males (AM).The images are the result of the overlap of both stainings. The arrows indicate neurons immunoreactive to both – PGP 9.5 and the other neuronal active substance studied.
A much smaller percentage of CART-positive neurons was observed in the OSP. In the young animals, these values amounted to 11.5 ± 0.6% and 11.7 ± 0.2% of all PGP 9.5-positive cells in young females and males, respectively. There were no statistically significant differences between genders in young animals. In adult females, the percentage of CART-positive cells amounted to 10.7 ± 0.3% and did not differ statistically significantly from that observed in young pigs of this gender. In turn, in adult males, the percentage of neurons immunoreactive to CART achieved the level of 10.4 ± 0.4% and was statistically significantly lower than that found in young males. There were no statistically significant differences between genders in adult animals (Fig 1).
In the ISP, CART-positive neurons were the least numerous. In young animals, the percentage of such neurons achieved the level of 0.2 ± 0.1% both in females and males. These values increased slightly with increasing age but still oscillated only around 1% of all PGP 9.5 – positive cells. In adult animals, the percentage of neurons containing CART amounted to 1.0 ± 0.1% and 1.6 ± 0.1%. Both in adult females and males, the values were statistically significantly higher than those noted in young animals of the same gender. Moreover, the difference in the percentage of CART-positive neurons between adult females and males was statistically significant (Fig 1). The results obtained in the present study are summarised in Table 2.
Discussion
During the present study, neurons containing CART have been noted in all types of enteric plexuses localised in the descending colon of both young and adult animals, which may confirm the multidirectional roles of this peptide in the digestive tract suggested by previous studies [17,28,47]. Both the present observation and previous investigations have shown the largest population of CART-positive neurons in the MP, which confirms that CART is a factor that, first of all, regulates intestinal motility [17,28,48,49]. In turn, the small number of CART-containing cells in the ISP suggests that this peptide plays a relatively minor role in intestinal secretion, although some previous studies have described the possibility of CART participation in the regulation of the gastrointestinal glands’ activity [50,51]. Moreover, the present study confirms intra-species differences in the distribution of CART in the enteric neurons [17,20,21,48]. However, it should be pointed out, that there are also clear differences in the percentage of CART-positive neurons between the present study and previous investigations performed on juvenile domestic pigs [14,48]. This fact strongly suggests that the synthesis and distribution of this peptide in the enteric neurons may depend on the race of animals, food and/or other not quite clear environmental factors.
It is commonly known that the ENS is a complex structure that may change under the influence of many pathological and physiological factors [22,24]. These changes include neuronal morphology, functions and neurochemical characterisation and reflect adaptive processes in response to acting stimuli [43,52,53]. Contrary to the influence of pathological and toxic factors on the ENS, the knowledge about changes in the enteric neurons caused by physiological factors (especially by maturation and aging) is rather scanty, but it is known that these changes may manifest themselves by modification of the number and shape of neurons and their ability to synthesise active substances [35,54,55].
The results obtained in the present study confirmed age-dependent changes in the population of the enteric neurons containing CART in the porcine colon. It should be underlined that maturation-induced changes in the intestine are multidirectional and concern the muscular layer, intramural blood vessels and intestinal secretory activity [56,57]. Moreover, maturation and aging directly affect the ENS, changing the length, number and appearance of axons and dendrites of the enteric neurons, as well as nerve electrophysiological properties and organisation of synapses [25,39].
The present results strongly suggest the participation of CART in adaptive processes connected with the development, maturation and aging of the large intestine. It is all the more likely that CART has been described as a factor participating in the development and differentiation of neuronal cells and synaptic connections within the central nervous system [58,59]. In light of this research, it is highly probable that CART may perform similar functions within the ENS. Age-dependent changes in the number of CART-positive neurons may also result from interactions between intestinal bacterial flora and the ENS. This thesis is supported by the understanding that the maturation process significantly affects the composition of the intestinal flora [60]. Additionally, research has shown that the microbiota can modulate the enteric neurons that contain CART [61].
The next reason for the increase in the percentage of enteric CART-positive neurons in older animals may be connected with the neurotrophic and protective functions of this peptide, as suggested by previous studies [62,63]. It is commonly known that throughout life, the intestinal wall is affected by many mechanical, chemical and stress stimuli [56,57,64]. Such a situation can lead to the development of inflammatory processes and various intestinal diseases in later life [65,66]. On the other hand, it is known that the ENS shows protective and regenerative abilities and protects the gastrointestinal tract in pathological states [4,10,43,53]. Therefore, the increase in the percentage of neurons containing CART in the MP and ISP noted in the present study may be connected with these processes because CART in some previous studies has been described as a protective factor [19,62,63,67]. However, in light of the facts mentioned above and noted in the present observation, the slight decrease in the number of CART–positive cells in the OSP of adult males in comparison to young males is not clear.
During the present study, gender-dependent differences in the percentage of CART-positive neurons in the colonic ENS were also noted, and these differences were more visible in adult animals. This fact confirms that the neurochemical characterisation of the enteric neurons depends on gender. Previous knowledge about the influence of gender on the organisation of the ENS is rather limited [39,68,69]. It is known that receptors for sex hormones are widely distributed in the gastrointestinal tract, including the enteric neurons and glial cells [70]. Previous studies have also reported that sex hormones affect the ENS. It was found that stimulation of oestrogen receptor β leads to neuroprotective reactions involving enteric glial cell proliferation and neuronal differentiation [71].
Nevertheless, previous reports concerning gender-dependent differences within the ENS are inconclusive. Some of them have described such differences in the reaction of the ENS to some pathological and stress stimuli [72], as well as in the neurochemical characterisation of the enteric neurons [39]. In turn, other studies have not reported diversities in the organisation of the ENS between males and females [73]. The results obtained in the present study have confirmed gender-dependent differences in CART distribution in the enteric neurons. This is in agreement with previous reports on the central nervous system, where the knowledge about CART is better developed and gender-dependent diversity in the distribution and functions of structures containing CART have been reported [74].
Conclusions
To sum up, the results obtained in the present study have shown that the percentage of the neurons containing CART in the colonic ENS of the domestic pig depends not only on the type of the enteric plexus but also on the age of the animals. The presence of CART-positive neurons in all types of enteric plexuses confirms the multidirectional roles of CART in the regulation of large intestine functions. In turn, age-dependent changes in the number of CART-positive enteric neurons strongly suggest that this peptide plays a part in the processes connected with the development, maturation, and/or aging of the ENS and gastrointestinal tract. The results have also shown gender-dependent diversities in the distribution of CART in the enteric neurons. This fact shows that CART-positive neurons are influenced by sex hormones and suggests various exact functions of this peptide in the male and female gastrointestinal tract. However, it should be pointed out that clarification of all issues related to the exact functions of CART in the regulation of the functioning of the ENS in the large intestine requires further comprehensive research.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gabella G. Innervation of the gastrointestinal tract. Int Rev Cytol. 1979;59:129–93. doi: 10.1016/s 0074-7696(08)61662-9 226496 · doi ↗ · pubmed ↗
- 2Rao M, Gershon MD. Enteric nervous system development: what could possibly go wrong?. Nat Rev Neurosci. 2018;19(9):552–65. doi: 10.1038/s 41583-018-0041-0 30046054 PMC 6261281 · doi ↗ · pubmed ↗
- 3Furness JB, Callaghan BP, Rivera LR, Cho H-J. The enteric nervous system and gastrointestinal innervation: integrated local and central control. Adv Exp Med Biol. 2014;817:39–71. doi: 10.1007/978-1-4939-0897-4_3 24997029 · doi ↗ · pubmed ↗
- 4Makowska K. Chemically induced inflammation and nerve damage affect the distribution of vasoactive intestinal polypeptide-like immunoreactive (VIP-LI) nervous structures in the descending colon of the domestic pig. Neurogastroenterol Motil. 2018;30(11):e 13439. doi: 10.1111/nmo.13439 30109906 · doi ↗ · pubmed ↗
- 5Schneider S, Wright CM, Heuckeroth RO. Unexpected Roles for the Second Brain: Enteric Nervous System as Master Regulator of Bowel Function. Annu Rev Physiol. 2019;81:235–59. doi: 10.1146/annurev-physiol-021317-121515 30379617 · doi ↗ · pubmed ↗
- 6Morikawa S, Komuro T. Distribution of myenteric NO neurons along the guinea-pig esophagus. J Auton Nerv Syst. 1998;74(2–3):91–9. doi: 10.1016/s 0165-1838(98)00131-3 9915623 · doi ↗ · pubmed ↗
- 7Reiche D, Michel K, Pfannkuche H, Schemann M. Projections and neurochemistry of interneurones in the myenteric plexus of the guinea-pig gastric corpus. Neurosci Lett. 2000;295(3):109–12. doi: 10.1016/s 0304-3940(00)01617-7 11090986 · doi ↗ · pubmed ↗
- 8Zhang G-Q, Yang S, Li X-S, Zhou D-S. Expression and possible role of IGF-IR in the mouse gastric myenteric plexus and smooth muscles. Acta Histochem. 2014;116(5):788–94. doi: 10.1016/j.acthis.2014.01.011 24630395 · doi ↗ · pubmed ↗