Inferring replication states of bacteria and viruses in enrichment cultures via long-read sequencing
Sophie A Simon, André R Soares, Till L V Bornemann, Adrian Lange, Lea Griesdorn, Adrián Fuentes, Marie Dieckmann, Beate A Krok, S Emil Ruff, Michael Hügler, Cristina Moraru, Alexander J Probst

TL;DR
This study uses long-read sequencing to track replication in bacteria and viruses from enrichment cultures, revealing which microbes are active and how they replicate.
Contribution
A novel method combining long-read sequencing and BrdU labeling to simultaneously analyze replication and genome structure in enrichment cultures.
Findings
114 out of 118 high-quality genomes were found to be actively replicating in activated sludge enrichment cultures.
Some genomes showed BrdU-rich regions, suggesting prophages in the lytic cycle.
The method distinguishes between slow and rapidly replicating organisms based on BrdU incorporation and abundance changes.
Abstract
Most microorganisms cannot be cultured in isolation, necessitating sophisticated methods for studying their (eco)physiology. While numerous approaches can probe the activity of given microbes in enrichment cultures, no single technique can render simultaneous data on both metabolic capacities and mobile genetic elements. Here, we apply long-read sequencing to monitor the incorporation of non-canonical bases in genome-resolved metagenomic datasets and elucidate the replication patterns of both bacteria and phages. This technology enables the simultaneous reconstruction of both prokaryotic and viral genomes (alongside genomics downstream analyses like metabolic predictions), in addition to providing information regarding their replication in enrichment cultures. By spiking the base analog 5-bromo-2′-deoxyuridine (BrdU) into activated sludge microcosms, we determined that 114 of the 118…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
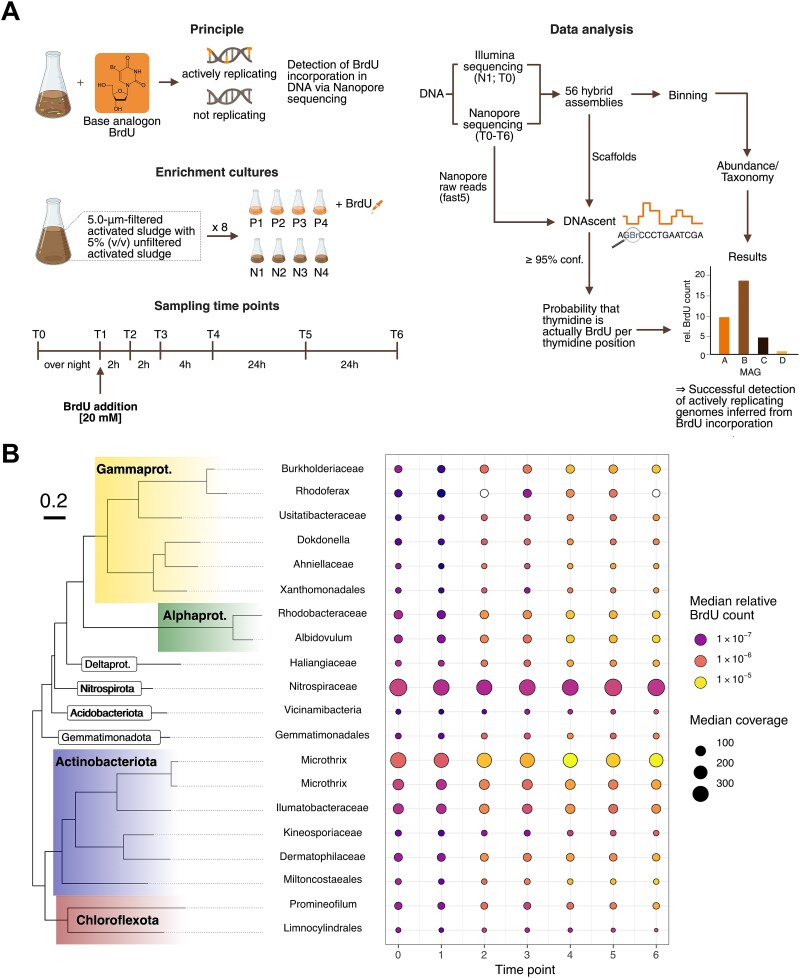 Figure 1
Figure 1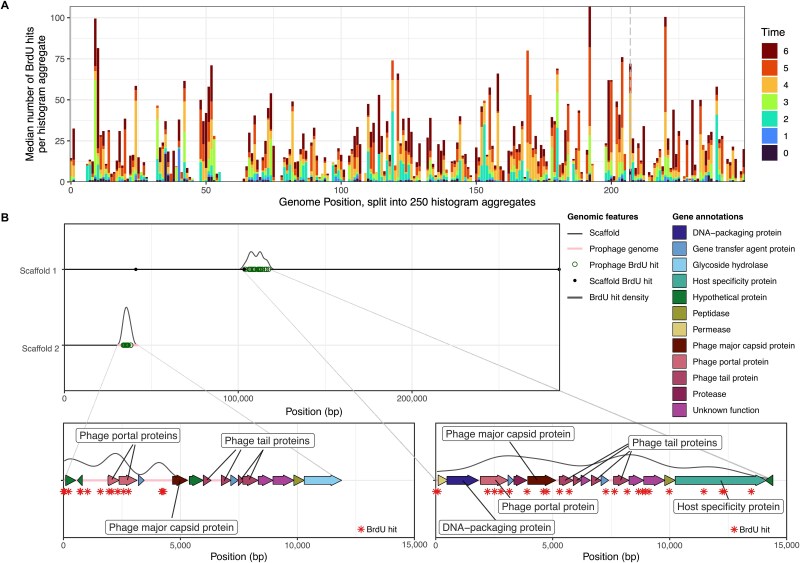 Figure 2
Figure 2- —ERC Synergy
- —German Research Foundation (DFG)
- —European Union’s Horizon 2020 Research and Innovation Programme
- —German Federal Ministry of Education and Research within the project “MultiKulti”
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCRISPR and Genetic Engineering · Bacteriophages and microbial interactions · Viral Infectious Diseases and Gene Expression in Insects
As most microbes are refractory to isolation, researchers often study them in ecosystems or enrichment cultures, the latter affording approaches to probe nutrient turnover and/or metabolic activity [1–3]. However, each of these methods has its limitations. To the best of our knowledge, no single technique can be used to simultaneously predict the replication states of microorganisms and mobile genetic elements (e.g. viruses, plasmids) as well as perform genome-resolved metagenomics of enrichment cultures. Methods predicated on genome coverage have proven to be inaccurate, most likely due to lacking consideration for strain heterogeneity [4, 5]. The thymidine base analog 5-bromo-2′-deoxyuridine (BrdU) incorporates into newly synthesized DNA and can be probed with antibodies [6]. While this technology gained popularity in the early 2000s, its application has remained limited to FISH-based microscopic analyses or 16S rRNA gene sequencing [7, 8]. Here, we exploit BrdU to monitor the replication of microorganisms and viruses in each of eight enrichment cultures from activated sludge (four with BrdU amendment and four as a negative control). We monitored these cultures across seven time points, resulting in 56 distinct ONT (Oxford Nanopore Technologies) metagenomes and 15 Illumina libraries (Fig. 1A).
We used DNAscent [9], which jointly evaluates mappings of Nanopore reads to a reference (metagenome-assembled genome (MAG) or assembly) and electric signal data of the mapped Nanopore reads, to estimate the probability that a predicted thymidine is more likely BrdU and applied it to complex enrichment cultures from activated sludge (BrdU confidence > = 95%; Fig. 1A). We verified this approach on (i) labeled and unlabeled 16S rRNA gene PCR products and (ii) four pure cultures of easy-to-cultivate model organisms (Escherichia coli, Bacillus subtilis, Corynebacterium glutamicum, and Salmonella enterica grown in pure cultures spiked with 20 μM BrdU) with well-characterized genomes (BrdU calling confidence > = 90%; see Methods in Supplementary Information and Supplementary Fig. S1). PCR products generated with either dTTP (unlabeled) or BrdUTP (labeled) were used as control to ensure the exclusive incorporation of BrdUTP in the positive control and the absence of DNA-modifications in the negative control. Long-read sequence metagenomes were then generated from activated sludge enrichments with and without the addition of BrdU, which were tracked over seven time points (Fig. 1A) in sets of four biological replicates each. This sequencing effort was complemented with Illumina short-read sequencing, generating a total of 118 dereplicated high-quality MAGs affiliated with Proteobacteria (38 MAGs), Actinobacteriota (25), Bacteroidota (20), and Chloroflexota (12). Of the 118 total MAGs generated, 114 incorporated BrdU, suggesting a broad applicability of BrdU for enrichment studies. Twenty-nine MAGs spanning nine phyla showed more than 0.001 normalized BrdU counts (see Methods in Supplementary Information). These included species of Microthrix (Actinobacteriota) and several Alpha- and Gammaproteobacterial genera, which were only moderately abundant in the enrichment culture (Fig. 1B). Highly abundant Nitrospirae MAGs showed low BrdU incorporation, which was consistent with their slow growth rate [10] and relative abundance change over time (Fig. 1B). Analyzing the BrdU incorporation in one distinct MAG revealed that BrdU can be incorporated randomly (Fig. 2A), as the origin of replication (oriC) did not exhibit significantly increased BrdU detection (Chi-square test, no significant P-values (<0.005) detected under the null hypothesis, gray dashed line) and certain regions showed no incorporation at all. MAG-level BrdU incorporation and coverage in samples augmented with BrdU yielded marginal positive correlations (Kendall’s τ = 0.205, P < 0.001, with timepoints 0 and 1 excluded as these did not contain BrdU), with minimal false positive BrdU hits detected in samples sans BrdU addition (see Supplementary Fig. S3), which are virtually eliminated with the use of R10.4.1 flowcells as demonstrated by BrdUTP PCR (see Supplementary Fig. S4). The yet-to-be improved BrdU calling confidence in combination with metagenomic relative abundance estimations do not yet allow for absolute quantitative insights. Improvement of the BrdU calling confidence with R10.4.1 and future chemistries will also increase the possibility to use the presented BrdU-Nanopore sequencing method in a more quantitative way.
Applying BrdU to a pure culture of S. enterica serovar Typhimurium LT2 showed increased BrdU in one of the prophages, making induction by BrdU a possibility (Supplementary Fig. S2). Consequently, focusing on prophages, and using the surrounding scaffold of BrdU incorporation from the host as a baseline, we examined BrdU induction of prophages across the enrichment culture sample sets. Since the same prophages were not induced across replicates (see prophage clustering relative to BrdU incorporation; Supplementary Fig. S7) we conclude that induction observed in some cases was random and not prompted by the addition of BrdU. In these instances, we detected a significant increase in BrdU incorporation in prophage regions encoding for prophage capsid and tail proteins, indicative of phage induction and replication in the hosts’ cytoplasm (Fig. 2B). As such, we recommend the use of BrdU alongside ONT for monitoring prophage induction in respective assays.
Despite the challenges and shortcomings involved in reproducing enrichment cultures, assaying a broad spectrum of substrates and monitoring the growth responses of microorganisms of interest via the herein introduced BrdU-ONT assay will empower efforts toward cultivating not-yet-cultivated microbes. Identifying the induction of prophages that might hinder the growth of uncultivated organisms of interest will factor largely in future strategies to optimize cultivation conditions. In such cultivation efforts, it will be essential to account for the potential cytotoxicity of BrdU which could unintentionally impact microbial growth, yet the concentration of BrdU used in within this study (20 μM) has reportedly shown low toxicity [7]. We believe that as ONTs continue to improve chemistry and flow cell technology (e.g. DNAscent was recently adapted to work with R10 flow cells), activity-based metagenomics will no longer require short-read data to infer growth/replication from high-quality ONT-based MAGs [11]. Taken together, BrdU coupled to ONT base calling is a valuable tool for studying the activity of microbes in enrichment cultures along with deciphering their genomic content.
Supplementary Material
Simon_et_al_BrdU_SuppText_ISMEComms_final_ycaf041(1)
Simon_et_al_BrdU_ISMEComms_SupplementaryTables_ycaf041
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Singer E, Wagner M, Woyke T. Capturing the genetic makeup of the active microbiome in situ. ISME J 2017;11:1949–63. 10.1038/ismej.2017.5928574490 PMC 5563950 · doi ↗ · pubmed ↗
- 2Ünal B, Perry VR, Sheth M et al. Trace elements affect methanogenic activity and diversity in enrichments from subsurface coal bed produced water. Front Microbiol 2012;3. 10.3389/fmicb.2012.00175 PMC 334927122590465 · doi ↗ · pubmed ↗
- 3Nakamura K, Iizuka R, Nishi S et al. Culture-independent method for identification of microbial enzyme-encoding genes by activity-based single-cell sequencing using a water-in-oil microdroplet platform. Sci Rep 2016;6:22259. 10.1038/srep 2225926915788 PMC 4768102 · doi ↗ · pubmed ↗
- 4Long AM, Hou S, Ignacio-Espinoza JC et al. Benchmarking microbial growth rate predictions from metagenomes. ISME J 2021;15:183–95. 10.1038/s 41396-020-00773-132939027 PMC 7852909 · doi ↗ · pubmed ↗
- 5Brown CT, Olm MR, Thomas BC et al. Measurement of bacterial replication rates in microbial communities. Nat Biotechnol 2016;34:1256–63. 10.1038/nbt.370427819664 PMC 5538567 · doi ↗ · pubmed ↗
- 6Urbach E, Vergin KL, Giovannoni SJ. Immunochemical detection and isolation of DNA from metabolically active bacteria. Appl Environ Microbiol 1999;65:1207–13. 10.1128/AEM.65.3.1207-1213.199910049885 PMC 91166 · doi ↗ · pubmed ↗
- 7Pernthaler A, Pernthaler J, Schattenhofer M et al. Identification of DNA-synthesizing bacterial cells in coastal North Sea plankton. Appl Environ Microbiol 2002;68:5728–36. 10.1128/AEM.68.11.5728-5736.200212406771 PMC 129917 · doi ↗ · pubmed ↗
- 8Roux-Michollet DD, Schimel JP, Holden PA. Pushing the limits for amplifying Brd U-labeled DNA encoding 16S r RNA: DNA polymerase as the determining factor. J Microbiol Methods 2010;83:312–6. 10.1016/j.mimet.2010.09.01620883730 · doi ↗ · pubmed ↗