Structural basis for control of integrative and conjugative element excision and transfer by the oligomeric winged helix–turn–helix protein RdfS
Callum J Verdonk, Mark Agostino, Karina Yui Eto, Drew A Hall, Charles S Bond, Joshua P Ramsay

TL;DR
This study reveals how the RdfS protein controls the excision and transfer of a bacterial DNA element involved in nitrogen-fixing symbiosis.
Contribution
The study identifies a novel α1-helix in RdfS that enables oligomerization and indirect DNA recognition.
Findings
RdfS binds to DNA regions in the IntS attachment site and its own promoter to regulate ICE excision.
The α1-helix in RdfS is essential for oligomerization and DNA complex compaction.
Removing the α1-helix abolishes RdfS-mediated ICE excision and conjugative transfer.
Abstract
Winged helix–turn–helix (wHTH) proteins are diverse DNA-binding proteins that often oligomerize on DNA and participate in DNA recombination and transcriptional regulation. wHTH recombination directionality factors (RDFs) associated with tyrosine recombinases, stimulate excision of prophage and integrative and conjugative elements (ICEs). RdfS is required for excision and conjugation of the Mesorhizobium japonicum R7A ICE, ICEMlSymR7A, which carries genes for nitrogen-fixing symbiosis. We show RdfS binds to DNA regions within the IntS attachment site (attP) and within the rdfS promoter, enabling RdfS to coordinate rdfS/intS expression and stimulate RdfS/IntS-mediated ICEMlSymR7A excision. Several RdfS DNA-binding sites were identified. However, no consensus motif was apparent and no individual nucleotide substitutions in attP prevented RdfS binding. RdfS forms extensive helical filaments…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
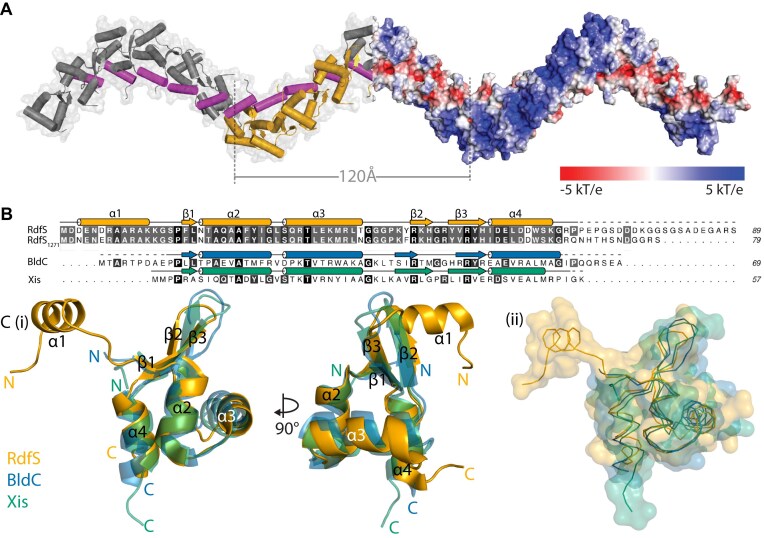 Figure 1
Figure 1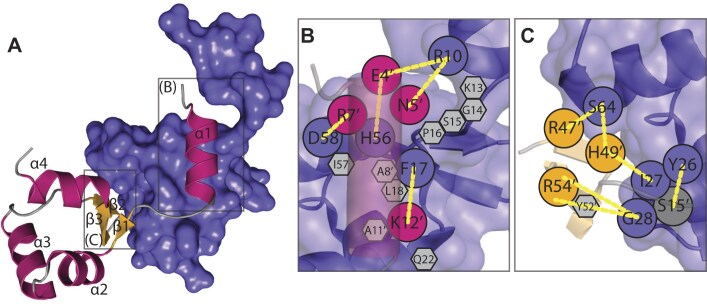 Figure 2
Figure 2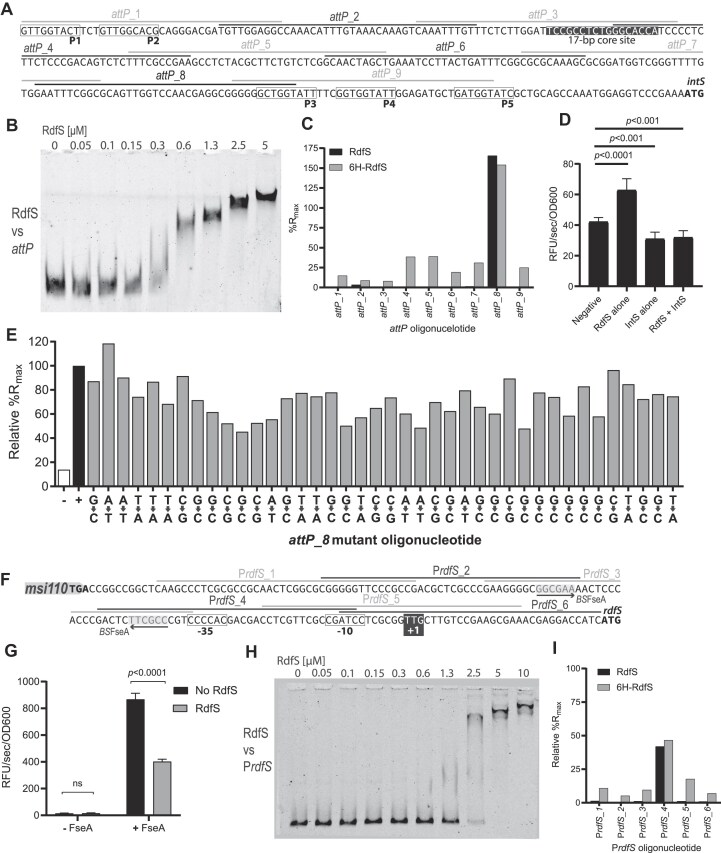 Figure 3
Figure 3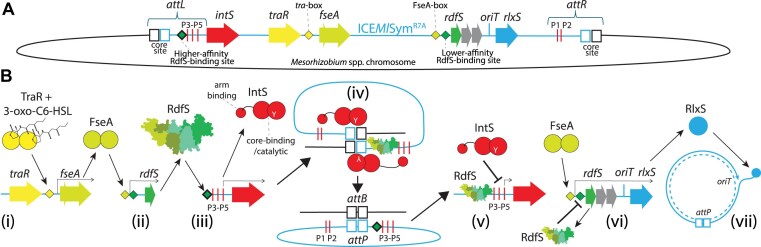 Figure 4
Figure 4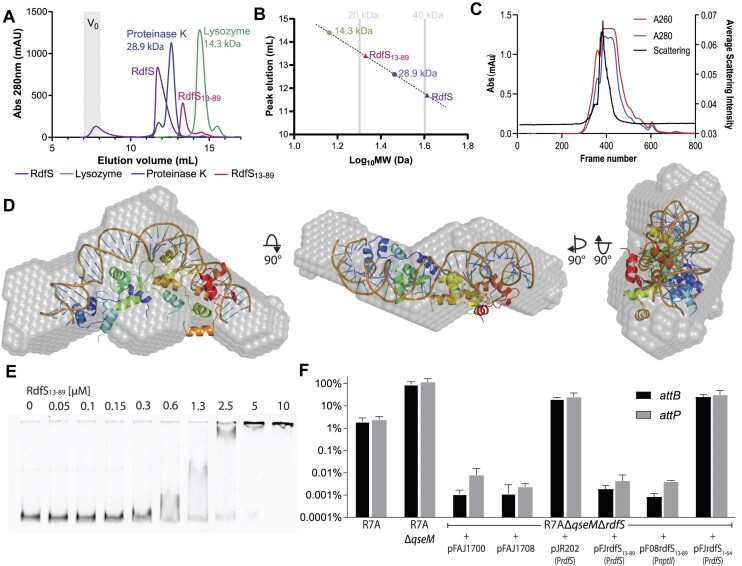 Figure 5
Figure 5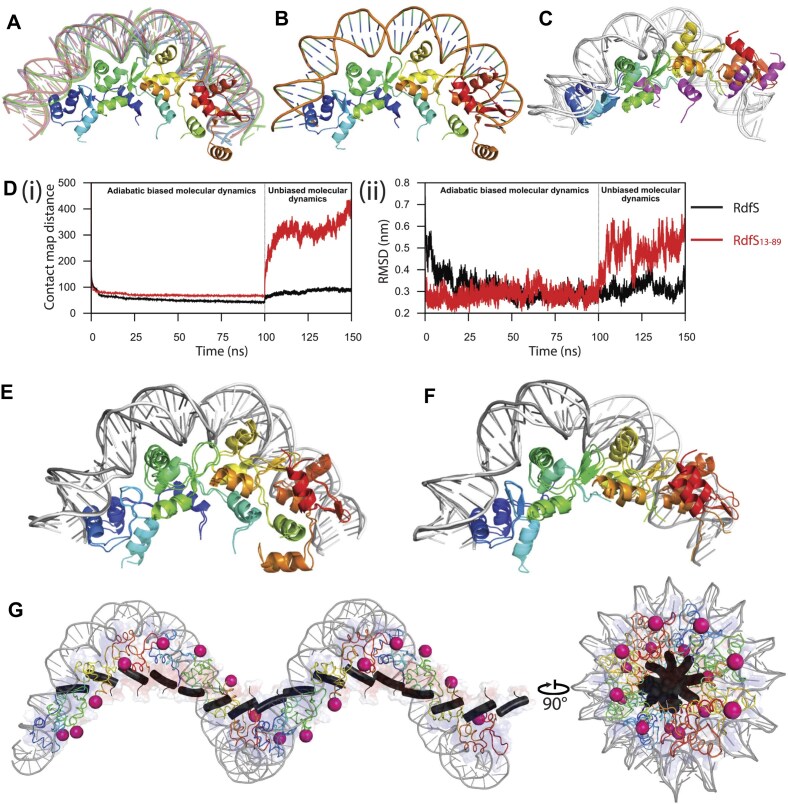 Figure 6
Figure 6- —Australian Research Council10.13039/501100000923
- —Australian Government10.13039/100015539
- —University of Western Australia10.13039/501100001801
- —Curtin University10.13039/100018548
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEnzyme Structure and Function · Metalloenzymes and iron-sulfur proteins · Neurological diseases and metabolism
Introduction
Integrative and conjugative elements (ICEs) are chromosomally integrating mobile genetic elements that often carry genes with selective advantages to their bacterial hosts, such as those necessary for biofilm formation, antibiotic and heavy metal resistances, virulence and pathogenicity, iron acquisition, and symbiotic nitrogen fixation [1–10]. Genome analyses indicate that ICEs are the most abundant type of conjugative element present in prokaryote genomes [11, 12]. ICEs reside integrated within the chromosomes of their hosts but can excise from the chromosome to form a transient circular element that is then able to transfer a single-stranded DNA copy through conjugation. Therefore, ICE excision and circularization is a prerequisite first step in ICE transfer. For most ICEs, it is unclear how the initiation of these distinct DNA recombination and conjugation processes are coordinated at a molecular level [13, 14].
Much like bacteriophage integration and excision, recombination between the ICE and bacterial chromosome is usually site-specific and catalyzed by a tyrosine or serine recombinase, also referred to as an integrase (Int). Integrases, together with specific DNA regions called attachment (att) sites, form nucleoprotein complexes that orchestrate DNA strand exchanges, facilitating both integrative and excisive recombination [15, 16]. Attachment sites for integrases of the tyrosine recombinase family contain a central core sequence that acts as the DNA substrate for strand cleavage and exchange. Recombination is catalyzed by the C-terminal recombinase domain of the integrase. During ICE integration, an attachment site attP present on the circular excised ICE is recombined with a corresponding chromosomal attB site. The attP region also contains specific ‘arm’ (also known as ‘P’) sequences flanking the recombined ‘core’ region. These arm sites interact with the N-terminal DNA-binding domain of the integrase and support the three-dimensional organization of the nucleoprotein complex during integration and excision reactions [17, 18]. Following ICE integration, recombined attP and attB sites produce chimeric attL and attR sites, each containing a copy of the core sequence and some of the arm sites derived from attP. The resulting recombined core sequences within attL and attR form direct DNA sequence repeats flanking the integrated ICE [15, 16, 19].
While integrase enzymes can catalyze both integrative and excisive recombination reactions, for most tyrosine recombinases, the formation of the integration products attL and attR is favoured. Excisive recombination often requires a recombination directionality factor (RDF) protein, also known as an excisionase. Tyrosine-recombinase RDFs are typically small positively charged DNA-binding proteins that bind DNA attachment sites and, in some cases, interact with integrase proteins directly [20–24]. While RDFs are extremely diverse at the primary amino-acid sequence level, they are frequently winged helix–turn–helix (wHTH)-domain proteins [20, 25]. wHTH proteins often also have roles as transcriptional regulators [26–33] and some are regarded to be nucleoid-associated proteins [34–38] that may perform roles analogous to eukaryotic histones; binding, bending, wrapping, and/or bridging along DNA to form large nucleoid complexes [39, 40].
The symbiosis ICE of Mesorhizobium japonicum R7A, ICEMlSym^R7A^, is a 502-kb ICE carrying genes necessary for nitrogen-fixation and symbiosis with Lotus spp. ICEMlSym^R7A^ encodes genes for root nodule formation, nitrogen fixation, metabolism, and protein secretion [6, 41, 42]. Integration and excision of ICEMlSym^R7A^ requires the tyrosine recombinase (integrase) IntS, which is encoded by the intS gene located immediately downstream of the attL site on the integrated ICEMlSym^R7A^ element. Experiments using a constructed ‘mini-ICE’ carrying only the attP region or the attP-intS region (on a plasmid unable to replicate in Mesorhizobium) reveal attP and intS are the only ICEMlSym^R7A^-encoded factors required for efficient integration. The 283-bp attP sequence contains five IntS P/arm-type sequence motifs (consensus TGKTGGTATC), and all five P/arm-type sites are required for efficient integration [43].
Most RDF genes are positioned nearby their cognate integrase genes. However, the ICEMlSym^R7A^ RDF gene rdfS is unique in that it is positioned as the first gene in an operon containing conjugation genes traF-msi107 and is upstream of the origin of transfer (oriT) site and the conjugative relaxase gene rlxS [19]. This genetic organization of rdfS is conserved on numerous ICEs and plasmids found throughout the proteobacteria [14], suggesting the stimulation of ICEMlSym^R7A^ excision has become intertwined with the activation of conjugation. Interestingly, deletion of rdfS not only abolishes ICEMlSym^R7A^ excision, but also abolishes the ability of ICEMlSym^R7A^ to mobilize a plasmid carrying a cloned copy of the ICEMlSym^R7A^oriT [43], suggesting RdfS is also required for activation of conjugation.
Introduction of a plasmid overexpressing rdfS is lethal in M. japonicum R7A and only cells that have lost ICEMlSym^R7A^, named R7ANS, survive [19]. The R7ANS strain contains a repaired attB site resembling a naïve host capable of receiving ICEMlSym^R7A^. Introduction of the rdfS gene expressed on a plasmid stimulated complete excision and loss of the attP-intS mini-ICE from the cell population. Introduction of a similar plasmid over-expressing intS only moderately increases excision frequency (measured by attP abundance) and loss of the mini-ICE (attB abundance), suggesting both RdfS and IntS together are required to strongly stimulate IntS-mediated excision [44, 45].
Several ICEs related to ICEMlSym^R7A^ (referred to as the ICESym family [14]) exist in a tripartite configuration. These tripartite ICEs are present as three separate DNA sections in their host chromosomes. Tripartite ICEs assemble into a single circular element prior to conjugative transfer and then separate back into three parts integrated within the chromosomes of recipients [46]. The archetypical tripartite ICE, ICEMcSym^1271^ from Mesorhizobium ciceri WSM1271, encodes three site-specific tyrosine recombinases—IntS, IntG, and IntM. During ICEMcSym^1271^ integration, the circularized tripartite ICE, which carries three distinct attP sites, integrates into the chromosome at one of the three different attB sites and then recombines twice more with the chromosome to separate the ICE into three sections [46, 47]. Tripartite ICE excision is the reverse of this process and requires an orthologue of RdfS, RdfS_1271_, which shares 85% amino-acid identity with RdfS. ICEMcSym^1271^ encodes two additional RDF genes rdfG and rdfM, located adjacent to their cognate integrase genes intG and intM. Expression of RdfS_1271_ activates transcription from both the rdfG and rdfM promoters [44], thus coordinating all three excision reactions and formation of a single circular excised ICEMcSym^1271^ prior to transfer.
Other structurally characterized wHTH proteins have been shown to oligomerize in a head-to-tail fashion along DNA, forming variable-length nucleoprotein structures with pleotropic roles on transcription and recombination. In this work, we interrogate the previously solved RdfS crystal structure [45] to gain insight into the structural basis for the various functions of RdfS. RdfS oligomerized in solution even in the absence of DNA, and in crystals, RdfS oligomers have been shown to form an infinite superhelical polymer [45]. Compared to other wHTH proteins, RdfS carries an additional N-terminal α helix positioned at the core of the superhelical axis of RdfS polymers within crystals. DNA-binding assays confirmed that RdfS specifically bound DNA sites within attP and within the promoter regions of rdfS and rdfG. However, no obvious DNA-binding sequence motif was identified within these regions and no individual nucleotides within the attP site abolished binding. Removal of the unique N-terminal α-helix of RdfS diminished but was not essential for DNA binding in vitro; however, its removal abolished the ability of RdfS activate ICEMlSym^R7A^ excision and conjugative transfer. We propose that the unique N-terminal α-helix enhances RdfS oligomerization in vivo and that variable length RdfS oligomers specifically recognize diverse DNA sites on ICEs through an indirect readout DNA recognition mechanism.
Materials and methods
Microbiological techniques
Bacteria were cultured as previously described [19, 43, 48, 49] and further details can be found in the supplementary data. A list of strains and plasmids used in this study can be found in Supplementary Table S1. Oligonucleotides and synthetic DNA are listed in Supplementary Table S2. DNA extraction, plasmid construction, and β-galactosidase assays were carried out as previously described [49–52], and further information can be found in the supplementary data. ICEMlSym^R7A^ conjugation assays were carried out as described previously [46], with the exception that the R7ANS recipient strain carrying pPR3G was used in conjugation experiments and the selection medium contained gentamicin at 25 μg ml^−1^.
DNA-binding assays
Electrophoretic mobility shift assays (EMSAs) were performed as described previously [49, 53]. DNA containing the ICEMlSym^R7A^attP and PrdfS were amplified by polymerase chain reaction (PCR) using IRDye800-labelled DNA oligonucleotides. Binding reactions were carried out using labelled DNA concentrations of 10 nM as described previously [49], and detailed in the supplementary data. Surface plasmon resonance (SPR)-based DNA-footprinting assays were performed using the Re-Useable DNA-Capture Technique (ReDCaT) [54] on the Biacore T200 (GE Healthcare) as described previously [49, 53] with protein concentrations at 1 μM. The oligonucleotide array for the attP, PrdfS, and oriT DNA regions were constructed using the Perl script poop.pl [54].
Expression and protein purification
Purification of RdfS is described elsewhere [45]. 6HRdfS_1271_ and RdfS_13–89_ were purified using the same methods. All protein purifications were carried out on the ÄKTA pure chromatography system (GE Healthcare), using a 5 ml HisTrap column (Cytiva) for immobilized metal affinity chromatography and a HiLoad 16/600 Superdex 200 (GE Healthcare) for size-exclusion chromatography.
Gel-filtration chromatography
Analytical gel-filtration was performed with a Superdex 75 10/300 GL (Cytiva) column equilibrated with size exclusion chromatography (SEC) buffer (50 mM Tris–HCl, 300 mM NaCl, 5% (v/v) glycerol; pH 7.4) on an ÄKTA pure chromatography system (GE Healthcare) at 4°C with a flow rate of 0.5 ml min^−1^. Each protein was concentrated to ∼7 mg ml^−1^ and injected in aliquots of 50 μl. Wavelengths of 230, 260, and 280 nm were used to visualize the interactions. Further details are available in the supplementary data.
Small-angle X-ray scattering
Small-angle X-ray scattering (SAXS) data were collected on the SAXS/WAXS beamline at the Australian Synchrotron [55] with continuous data collection using a PILATUS 1M detector [56], using methods similar to those previously described [45]. Collection and processing parameters are shown in Supplementary Table S3. Further methodology can be found in the supplementary data.
Molecular modelling and simulation
A model of RdfS binding to DNA was generated by adapting homology modelling methods previously described for generating DNA–protein complexes [57]. The model was generated through aligning monomers of RdfS (PDB 8DGL) to the complex of the Streptomyces protein BldC with DNA (PDB 6AMA) [36]. Molecular dynamics simulations were performed using GROMACS 2020.3 [58] patched with PLUMED 2.6.1 [59]. Full details of molecular modelling and simulations are described in the supplementary data.
Quantitative PCR
Quantitative PCR assays to determine ICEMlSym^R7A^ excision rates were performed as previously described [19, 49]. Details of culture growth and primer efficiencies can be found in the supplementary data.
Results
RdfS is an oligomeric wHTH-domain with a unique N-terminal α-helix
We recently solved the X-ray crystal structure for RdfS, revealing it forms a head-to-tail oligomer containing four RdfS molecules within the asymmetric unit (ASU), arranged with eight-fold helical symmetry within the crystal [45] (PDB entry 8DGL; Fig. 1A). The infinite head-to-tail oligomerization of RdfS protomers results in a superhelical protein filament extending throughout the crystal. Each of the four independent molecules observed share an effectively identical structure of four α-helices and a single three-stranded β-sheet: α1 (RdfS residues 3–12), β1 (17–18), α2 (19–28), α3 (30–40), β2 (47–49), β3 (52–56), and α4 (57–65) (Fig. 1B). In these crystals the C-terminal 21 residues (24%) of RdfS are completely disordered. The primary amino-acid sequence of RdfS (residues 17–66) is most similar to the group 17 HTH domain, which includes several other excisionase proteins (InterPro IPR041657). Alignments of the RdfS structure with Mycobacterium virus Pukovnik excisionase (RDF) Xis or the MerR-family Streptomyces BldC protein (Fig. 1C) exhibit Cα root mean square deviations (RMSD) of <2 Å.
X-ray crystal structure of RdfS and comparisons to other wHTH-containing proteins. (A) Extended RdfS crystal structure revealing superhelical filaments. Electrostatic surface representation of 24 RdfS molecules is shown in red and blue at the right half of the helix, representing electronegative and electropositive regions, respectively (shown as a −5 to 5 kBT/ec gradient). The RdfS asymmetric unit of four molecules shown is depicted in cartoon format in yellow. The central α1 helix is shown in in magenta throughout the length of one full turn of the helix (eight RdfS molecules) spanning ∼120 Å. (B) Amino acid alignment of ICEMlSymR7A RdfS and closely related Mesorhizobium RdfS homologue from M. ciceri WSM1271 (RdfS1271). Secondary structural representation shown above based on the X-ray crystal of R7A RdfS. Other HTH 17 grouped proteins included are Streptomyces venezuelae BldC [36] and Mycobacterium virus Pukovnik Xis [60] with their relative secondary structure features shown above each sequence. (C) (i) Ribbon model of RdfS (PDB 8DGL), BldC (PDB 6AMK), and Xis (PDB 4J2N) aligned to show similar features with each N- and C-terminus marked, and each location of the α-helices and β-strands for RdfS labelled to match panel (B). (ii) Surface representation of RdfS, BldC, and Xis, with ribbon-backbone shown to highlight overall similarity between each structure [in same orientation as panel (C) (i)]. The RMSD between the wHTH domains of RdfS and BldC, and RdfS and Xis, are 1.66 Å (46 atoms) and 1.12 Å (39 atoms), respectively.
The superhelical RdfS filament is stabilized by two protein–protein interfaces. The first interface involves the unique RdfS N-terminal α-helix (α1) from Asp3-Lys12, which sits in a hydrophobic pocket within the adjacent RdfS protomer (Fig. 2A). As far as the authors are aware, the α1 helix is absent from all other structurally characterized wHTH-domain proteins. The α1 charged sidechains from Glu4 and Arg7 form salt-bridges with the adjacent protomers’ charged side-chains His56 and Asp58, respectively. Hydrogen bonds are also formed between the α1 Asn5 and Lys13 sidechains interacting with the backbones of Phe17 and Arg10 from the neighbouring protomer. The α1-mediated interaction between molecules is primarily stabilized by the nine hydrophobic residues which span the surface of α1 and the neighbouring RdfS protomer (Fig. 2B). The overall surface area of α1 is 1286 Å^2^ and with an interface area of 463 Å^2^ with the adjacent RdfS molecule it is the major source of interaction area between RdfS molecules in the crystal structure. The second interface is between the wing region (β2–β3) of one RdfS molecule and the helices α2 and α4 of the neighbour; and with fewer hydrophobic residues at this interface, hydrogen bonding is the primary mediator of interface stability. The positively charged side-chains from strands β2 (Arg47 and His49) and β3 (Arg54) of RdfS interact with the backbone oxygens of α2 (Gly28 and Ile27) and α4 (Ser64) of the neighbouring molecule (Fig. 2C). This wing–α2/α4 interaction has a lower interface area of ∼400 Å^2^ compared with the α1’–RdfS interactions, suggesting that it may contribute more weakly to association.
The interface between adjacent RdfS molecules in the crystal. (A) Ribbon diagram of RdfS molecule interacting with adjacent molecule as surface representation (purple). Ribbon colour scheme shown is strand; yellow, helix; magenta, and loop; grey. Schematic showing: (B) Close-up of RdfS N-terminus α-helix (α1) (transparent magenta cylinder) interacting residues. (C) RdfS β1–β2–β3 ‘wing’ region interacting with an adjacent RdfS protomer. Diagram residues coloured as in panel (A), hydrogen bonds which stabilize the interface shown as yellow. Hydrophobic contact residues shown as grey hexagons.
Typically, wHTH proteins contact both the DNA major groove through their recognition helix and the DNA minor groove with their β-sheet ‘wing’. The positions of the expected major and minor DNA groove binding regions of the wHTH motif and electrostatic potential along the RdfS polymer are consistent with RdfS being capable of binding long stretches of double-stranded DNA (dsDNA) as an oligomer. The tentative recognition helix α3 contains exposed positively charged basic residues Arg32, Lys36, and Arg38 and exposed acidic residues Glu35 and Gln31, which may stabilize the protein–DNA interface. The arrangement of basic amino acids on the external surface results in a strongly positive electrostatic potential (Fig. 1A) and a continuous DNA-binding interface running the length of the superhelical protein filament. Superimposing RdfS monomers with a selection of crystal structures of wHTH:DNA complexes (Supplementary Fig. S1) revealed the orientation of the RdfS α3 and β2-β3 wing in the crystal structure resembles the orientation of DNA-binding components of other members of the wHTH family.
RdfS binds the attP site upstream of intS and upstream of promoters PrdfS and PrdfG
Characterized RDFs have been shown to bind specific DNA regions within their cognate att sites and promote excisive recombination catalyzed by their cognate integrase. Consistent with RdfS playing a similar role in ICEMlSym^R7A^ excision, overexpression of RdfS stimulates excision and loss of ICEMlSym^R7A^ from cells. Likewise, overexpression of RdfS in the presence of a mini-ICE plasmid carrying only the attP-intS region also results in loss of the mini-ICE, indicating that RdfS likely binds the att regions of ICEMlSym^R7A^ to stimulate IntS-mediated excision. To test whether RdfS could bind the attP region present on the circularized ICEMlSym^R7A^, RdfS was purified as an N-terminal hexahistidine fusion protein (6H-RdfS) and the 6H tag was removed following purification. EMSAs were carried out with the 283-bp attP region (Fig. 3A). EMSAs revealed RdfS retarded the migration of 10 nM attP when supplied at concentrations of 0.3–0.6 μM. The attP fragment was increasingly retarded with increased RdfS concentrations, suggesting that additional RdfS molecules were potentially oligomerizing on the RdfS–DNA complex (Fig. 3B). While the ratio of RdfS:DNA required to retard attP was relatively high, RdfS was unable to shift a 100-bp control DNA region (100-bp of the downstream intS gene) even with 10 μM of RdfS (Supplementary Fig. S2), confirming that RdfS binding to the attP region was specific. To refine the location of RdfS binding to attP, SPR-based DNA-footprinting assays were carried out. A tiled array of 40-bp double-stranded oligonucleotides spanning the attP region was designed, with each dsDNA oligonucleotide overlapping the sequence of the adjacent oligonucleotide by 10 bp (Fig. 3A). SPR assays with these oligonucleotides revealed that purified RdfS (1 μM) induced a strong binding response with a single dsDNA oligonucleotide, attP8, the sequence of which overlaps the putative IntS arm site ‘P3’ towards the 3′ end of the attP region. The SPR binding response (%R_max) values were 150%–250% of the theoretical R_max_ (across multiple experiments), suggesting more than one RdfS molecule was bound to attP_8. Due to the apparent complexity of RdfS–DNA interactions we were unable to determine association/dissociation kinetics from SPR data. Interestingly, when 6H-RdfS was used in SPR experiments, binding responses with attP_8 were unchanged; however, binding to each of the other oligonucleotides increased, suggesting that the 6H tag slightly increased non-specific DNA binding (Fig. 3C).
RdfS DNA-binding to the ICEMlSymR7AattP and PrdfS. (A) Sequence of the minimal ICEMlSymR7AattP DNA region required for integration, highlighting the arrangement of putative IntS-binding P sites P1–P5 (labelled boxes) and central recombination region (including the putative IntS ‘core’ site, white text, black background). Oligonucleotides (40 bp) used in SPR-based DNA-footprinting experiments are indicated by alternating grey and black lines above the attP sequence, labelled attP_1-attP_9. (B) EMSA using fluorescently labelled attP DNA (10 nM) and two-fold increasing concentrations of RdfS. (C) SPR DNA-footprinting for RdfS and 6H-RdfS using individual 40-bp oligonucleotides from the attP region. Oligonucleotide attP_8 consistently exhibited the largest %Rmax response and was therefore used as a positive control in all subsequent SPR footprinting experiments. (D) β-Galactosidase activity in strains carrying a chromosomally integrated PintS::lacZ fusion (R7ANS::pFUS2P1-P5). Cultures were tested in the presence of RdfS (pSRKrdfS) and/or IntS (pJJ611), or with empty vectors (pSRKKm and pFAJ1708). (E) Analysis of SPR responses for dsDNA attP_8 oligonucleotides containing single base-pair substitutions. For each nucleotide position a substitution is indicated, where each base was switched for its complementary base on both strands of the dsDNA oligonucleotide. Each substitution was sequentially tested in SPR DNA-footprinting in a single run and the positive control attP_8 (+) and negative control attP_2 (−) oligonucleotides (shown as black and white bars) were run every 10th cycle and used to normalize drifting data over the duration of the experiment. (F) Sequence of the ICEMlSymR7A PrdfS region. Each oligonucleotide tested in SPR DNA-footprinting are shown as alternating grey and black bars above the sequence (labelled PrdfS_1-PrdfS_6). The PrdfS transcriptional activator FseA binding site (BSFseA) [61] is shaded with a grey background and the inverted repeat are shown by arrows. The −35, −10 regions and +1 transcriptional start site for PrdfS estimated from previous 5′RACE experiments are indicated. (G) β-Galactosidase activity of stationary-phase R7ANS cultures containing PrdfS-lacZ (pSDrdfS-lacZ) plasmid constructs in the absence of FseA (‘− FseA’) or containing PrdfS-lacZ with the presence of FseA (pFseArdfS-lacZ: ‘+ FseA’). Cells additionally contained vectors pSRKKm (No RdfS: black) or pSRKrdfS (RdfS: grey). (H) EMSA with RdfS and the PrdfS region shown in panel (F). (I) SPR DNA-footprinting of RdfS and 6H-RdfS against PrdfS 40-bp oligonucleotides. Values are shown as relative %Rmax to the attP_8 control oligonucleotide (i.e. where attP_8 has a %Rmax of 100%).
We attempted to further delineate the RdfS binding region within the attP8 sequence by carrying out SPR experiments with a 20-bp oligonucleotide array covering the *attP*8 sequence, with each oligonucleotide overlapping the adjacent oligonucleotides by 15 bp. However, none of these 20-bp dsDNA regions achieved >45% R_max_ observed for attP_8 (Supplementary Fig. S2), suggesting that either the optimal RdfS binding region in attP_8 was larger than 20-bp and/or that the positioning of the oligonucleotide did not capture the entire binding motif. To determine whether there were any critical nucleotides targeted by RdfS within the 40-bp attP_8 oligonucleotide, we created 40 new variants of attP8 dsDNA oligonucleotide, each with a single-base-pair substitution to its complement nucleotide. Strikingly, no single nucleotide substitution abolished RdfS binding specificity, although some nucleotide changes reduced the relative %R_max to ∼50%–60% that of the native sequence (Fig. 3E). Together, these data suggest that RdfS oligomers may bind the attP sequence over a relatively large DNA-binding region, and that while the binding to this region is specific, no individual nucleotides in the region are absolutely critical for RdfS binding.
Due to the position of attP_8 sequence directly upstream of intS, we wondered whether RdfS and/or IntS might regulate transcription of the intS gene. In previous work, a plasmid pFUS2P1-P5 was constructed carrying the attP region with the intS gene replaced by lacZ, thus forming an attL-integrated transcriptional fusion carrying the PintS promoter [43]. Therefore, we used this strain to report on the expression of intS in the presence and absence of rdfS. Measured β-galactosidase expression from the PintS-lacZ fusion was increased ∼53% above background levels when rdfS was expressed from plasmid pSRKrdfS, indicating RdfS activates transcription of intS (Fig. 3D). In contrast, β-galactosidase expression was decreased ∼25% below background levels in the presence of the intS-expressing plasmid pJJ611, revealing that IntS negatively regulates its own expression. When both pSRKrdfS and pJJ611 were introduced, β-galactosidase expression was similar to the strain carrying the intS-plasmid pJJ611 alone, indicating that the negative autoregulation by IntS was dominant over the observed transcriptional activation by RdfS. In summary, the results of these experiments are consistent with a model where RdfS activates transcription from PintS and once IntS is produced, IntS negatively autoregulates its own expression.
Artificial overexpression of RdfS from a plasmid is lethal in R7A cells and leads to curing of ICEMlSym^R7A^ from cells [19]. However, we have never observed natural curing of ICEMlSym^R7A^ from cells during culture, suggesting that RdfS might negatively regulate its own transcription from PrdfS and prevent rdfS overexpression. To explore this, a previously constructed PrdfS-lacZ fusion was used to report on PrdfS transcription using β-galactosidase assays [50]. Since there is negligible expression from PrdfS in the absence of the transcriptional activator FseA, a plasmid containing both an IPTG-inducible copy of fseA and a PrdfS-lacZ transcriptional fusion was used to estimate transcription from PrdfS (Fig. 3G). A second plasmid carrying rdfS also under control of the lac promoter (pSRKrdfS) was used to express RdfS. As previously demonstrated, strong PrdfS-lacZ expression was observed from the pFseArdfS-lacZ plasmid because FseA activates transcription from PrdfS. However, when RdfS was additionally expressed from plasmid pSRKrdfS in the same cells, β-galactosidase activity was reduced ∼54%, indicating RdfS also binds PrdfS and attenuates the transcriptional activation by FseA.
EMSAs were performed with purified RdfS and a 162-bp region containing PrdfS. RdfS began to reduce migration of this DNA region when supplied at 1.3 μM. This concentration is approximately double the concentration of RdfS required to shift the attP region, indicating RdfS has an apparent weaker affinity for its own promoter than attP (Fig. 3H). We next again used SPR-footprinting with 40-bp tiled arrays of the PrdfS region to identify RdfS binding region(s). A single 40-bp dsDNA oligonucleotide PrdfS4 was bound by RdfS. In comparison to the 40-bp attP region identified previously, the 40-bp rdfS binding response was 52% of the theoretical R_max, which is ∼43% that of the binding response observed for the attP_8 oligonucleotide (Fig. 3I). The sequence of PrdfS_4 overlapped the −35 and −10 region of PrdfS, immediately downstream of the FseA binding site, consistent with RdfS repressing transcription by potentially occluding RNA polymerase from PrdfS (Fig. 3A). Shorter 20-bp dsDNA oligonucleotides derived from the PrdfS_4 sequence did not respond to RdfS in SPR experiments, again suggesting the optimal RdfS DNA binding site may be larger than 20 bp (Supplementary Fig. S2). Overall, the results of these experiments support a model of ICEMlSym^R7A^ excision in which RdfS first binds and stimulates intS transcription and stimulates IntS-mediated excision, after which IntS represses its own transcription from PintS. Once RdfS concentration increases above a certain threshold, RdfS binds PrdfS and represses its own expression, tentatively explaining why RdfS-dependent growth inhibition and loss of ICEMlSym^R7A^ has not been observed without artificial overexpression of rdfS (Fig. 4).
Stepwise induction of ICEMlSymR7A excision and transfer coordinated by IntS and RdfS. (A) A simplified depiction of integrated ICEMlSymR7A (light blue) including the attL and attR regions, the putative arm-type IntS-binding sites P1–P5 and other genes and binding sites involved in coordinating ICEMlSymR7A excision (as labelled). (B) A stepwise model of ICEMlSymR7A excision and transfer. (i) In transfer-active cells, the quorum sensing transcriptional activator TraR together with 3-oxo-C6-HSL binds a tra-box upstream of fseA and stimulates transcription (fseA is present as a two-part open-reading frame msi172-msi171 and the FseA protein is translated following a programmed ribosomal frameshift). (ii) An FseA dimer binds the FseA box upstream of PrdfS and activates transcription of rdfS and downstream genes involved in transfer (traF, msi107, and rlxS). (iii) RdfS (shown as tetramer in shades of green), either as monomers or potentially as preformed oligomers bind the attL region of ICEMlSymR7A upstream of the P3 arm-type IntS binding site. RdfS stimulates transcription of intS and production of IntS proteins. (iv) IntS proteins bind the core regions and arm-type sites present within attL and attR and together with RdfS, IntS catalyzes the excision of ICEMlSymR7A through recombination of attL and attR, producing attP and attB. (v) Once excised, attP-bound IntS protein prevents further transcription of intS, likely keeping the ratio of IntS:RdfS low, preventing reintegration of ICEMlSymR7A. (vi) Once RdfS concentration increases above a certain threshold, RdfS binds the apparent lower-affinity binding site within PrdfS, repressing transcription from this promoter and thus preventing RdfS-mediated growth inhibition. (vii) Finally, following some unknown signal indicating a mating-pair has formed with a recipient cell, conjugation commences with RlxS nicking the oriT and piloting an ssDNA copy of ICEMlSymR7A through the type-IV secretion system to the recipient cell.
RdfS and RdfS_1271_ of the tripartite-ICE ICEMcSym^1271^ share 85% amino-acid identity over their aligned length and otherwise only differ in the length and sequence of their C termini (Fig. 1B). Previously we have demonstrated RdfS_1271_ activates transcription from the PrdfG and PrdfM promoters of ICEMcSym^1271^ [44], stimulated tripartite ICE excision through the sequential activities of integrases IntS_1271_, IntG, and IntM. To test if RdfS_1271_ exhibited similar DNA-binding characteristics to RdfS, hexahistidine-tagged RdfS_1271_ (6HRdfS_1271_) was expressed and purified from Escherichia coli. 6HRdfS_1271_ was tested in SPR experiments using the tiled oligonucleotide arrays designed for ICEMlSym^R7A^attP and PrdfS regions as shown in Fig. 3A and F. Similar to R7A RdfS, 6HRdfS_1271_ bound the ICEMlSym^R7A^attP8 and PrdfS4 oligonucleotides with the highest relative %R_max (Supplementary Fig. S3A), indicating an identical DNA-binding specificity on ICEMlSym^R7A^, despite RdfS_1271 originating from ICEMcSym^1271^. Next, tiled oligonucleotide arrays were designed spanning the sequences of ICEMcSym^1271^ promoters PrdfS, PrdfG, and PrdfM. For the ICEMcSym^1271^ PrdfS, 6HRdfS_1271_ bound a dsDNA oligonucleotide in similar relative position within the PrdfS1271 promoter region as RdfS did with its cognate promoter (offset 5′ by 8 bp: Supplementary Fig. S3B) confirming both RdfS and RdfS_1271_ bind in similar positions overlapping their own gene promoters. 6HRdfS_1271_ bound two distinct regions within the PrdfG promoter region (Supplementary Fig. S3C). One of these was centred 170 bp upstream of the rdfG start codon, while the other was positioned directly upstream of the rdfG start codon, overlapping the putative ribosome-binding site. Only very weak binding was observed for any of the oligonucleotides in the PrdfM array (Supplementary Fig. S3D); however, this is consistent with very weak transcriptional activation of the PrdfM promoter by RdfS_1271_ compared to much stronger activation of the PrdfG promoter [44].
RdfS oligomerizes alone in solution and together with DNA
Most wHTH protein sthat form head-to-tail oligomers do so only in the presence of their cognate DNA sites [34]. The crystal structure of RdfS indicates that it can, at least in the crystal environment, form extensive oligomers in the absence of any DNA. We carried out analytical gel-filtration experiments to determine the oligomerization states of RdfS in solution. In the absence of DNA, RdfS eluted as an asymmetric peak with a leading edge corresponding to a maximum of ∼40 kDa (the expected mass of a tetramer) and a long trailing edge suggesting the presence of conformational or compositional heterogeneity (a flexible oligomer, and the presence of smaller oligomers such as trimers or dimers) (Fig. 5A and B). To validate the potential oligomeric properties of RdfS when binding DNA, we performed size-exclusion chromatography-coupled synchrotron SAXS (SEC-SY-SAXS) of 6H-RdfS bound to attP8. Although there were multiple long trailing peaks for the 6H-RdfS-attP8 sample in the SEC elution absorbance profiles, there was only one clear peak for the average X-ray scattering intensity profile (Fig. 5C), suggesting a single stable complex was being formed for 6H-RdfS-*attP*8. Analysis of the 6H-RdfS-*attP8 complex indicated it shows properties of a monodisperse sample. The normalized Kratky plot indicates a flexible extended particle, with a peak above ∼1.1 and qRg* beyond √3 returning towards zero at high qRg, indicating elongation [62–64], commensurate with the likely shape of a nucleoprotein complex (Supplementary Fig. S4). SAXS-derived molecular weight estimates using size and shape [65] indicated a 6H-RdfS-attP_8 mass of 85 655 Da, while concentration-independent estimation with Consensus Bayesian Assessment [66] suggested a complex mass of 83 125 Da (21.55% probable; credibility interval of 75 300–86 950 Da at 91.27% probability). These molecular weight estimates indicate a tetramer of 6H-RdfS bound to the dsDNA 40-mer attP_8; however, they predict a mass slightly larger than the expected molecular weight of 76.5 kDa, corresponding to four copies of 6H-RdfS and attP_8. The slight over-estimation for the molecular mass of the 6H-RdfS-attP8 nucleoprotein complex is likely due to the increased scattering intensities of more electron-dense nucleic acids [67]. Generally adjusting for the difference in electron intensities for nucleic acids [68], the molecular weight estimates from the scattering approximates 71.5 kDa—within 7% of the expected molecular weights (Supplementary Table S3). Ab initio bead models from the scattering approximates an envelope shape which superficially matches a generated RdfS–DNA molecular dynamics model (Fig. 5D). While fits into these bead models are poor, this is unsurprising given the absence of the 6H tag and disordered C-termini in the molecular dynamics-generated RdfS–DNA model—which is reflected in the residuals at low and mid-q (Supplementary Fig. S4). We concluded from these data that RdfS is binding as a tetramer to the 40-bp *attP*8 DNA target, suggesting 10-bp DNA per molecule of RdfS, comparable with previously characterized wHTH proteins which form multimers on DNA [36, 69–71].
RdfS oligomerization, DNA binding and in vivo functions are impacted by deletion of α1. (A) Comparison of analytical size exclusion elution profiles of RdfS and RdfS13–89. RdfS elutes larger than 28.9 kDa, indicating likely oligomeric states of tetramers >40 kDa, with some soluble aggregation/large multimer complexes appearing at the void volume (V0; shown as grey). α1-truncated RdfS (RdfS13–89) elutes between 14.3 and 28.9 kDa in size, indicating dimers or trimers in solution (chromatography peak slightly larger than 20 kDa). (B) Log10 plot of SEC data from (A) for estimation of RdfS/RdfS13–89 complex sizes. (C) Ultraviolet elution profile showing absorbance at 260 (red) and 280 (blue) nm, and average intensity of scattering of the 6H-RdfS-attP_8 (black). Each frame represents 1 s of exposure. (D) Most representative (as determined by DAMAVER) DAMMIF ab initio envelope bead model of the 6H-RdfS-attP_8 complex, with a representative atomic structure of the RdfS–DNA complex from molecular dynamics simulations unbiased adiabatic model shown within. This atomized model excludes the extended N- and C-termini present in the scattering data and is only shown as a representative of a superficial visualization of a potential nucleoprotein arrangement fit to the ab initio envelope. (E) EMSA showing RdfS13–89 retarding the migration of the fluorescently labelled attP region. The N-terminal truncated RdfS13–89 remained capable of binding attP DNA with ∼50% the affinity of RdfS. Much like the EMSAs with RdfS, migration of attP was increasingly retarded with increasing concentrations of RdfS13–89, suggesting RdfS13–89 was still likely capable of oligomerizing on DNA with increasing concentrations. (F) The relative abundance (relative to the R7A chromosome copy number) of attP and attB (present only when ICEMlSymR7A is excised) were measured by quantitative PCR to determine the frequency of ICEMlSymR7A excision. Plasmids expressing RdfS or truncated RdfS variants RdfS13–89 and RdfS1–64 were introduced to determine if each variant was functional in activating ICEMlSymR7A excision. The R7AΔqseMΔrdfS background was used as it is derepressed at a regulatory level for ICEMlSymR7A excision and conjugation but lacks rdfS gene required to excise and transfer [52]. The promoter used to drive expression of each of the RdfS variants is shown in brackets under the name of each plasmid.
Molecular dynamics simulations suggest that the N-terminal helix stabilizes RdfS oligomers and compacts the RdfS quaternary structure
Despite exhaustive attempts, we were unable to produce crystals of RdfS complexed with DNA, so we used in silico methods to assess how RdfS tetramers might bind with DNA targets and to probe whether the additional N-terminal α1 helix of RdfS might be important for these interactions. Extracting the DNA regions from wHTH-DNA nucleoprotein crystal structures (Supplementary Fig. S1) and superimposing them on the finalized RdfS ASU tetramer (Fig. 6A), we observed a strongly curved DNA bend path. To ensure this DNA bending was physically realistic, we first compared it to other structures with curved DNA and noted the bend to have similar parameters to nucleosomal DNA (Fig. 6B). We then performed molecular dynamics simulations on protein-only tetramers of both full length and N-terminally truncated RdfS_13–89_ (Supplementary Fig. S5A and B). The RMSDs (Supplementary Fig. S5C and D) over the simulation time courses for each tetramer suggest that RdfS_13–89_ yields a more flexible tetramer than the full length RdfS; this is further supported by cluster analysis, which indicates a vastly greater number of clusters for RdfS_13–89_ versus RdfS (Supplementary Table S4). The distance between the centres of mass of the capping molecules over the simulation time courses for each model, as well as within the middle structure of the largest cluster (Supplementary Fig. S5E and F), indicate that RdfS_13–89_ yields a slightly more compact tetramer compared to the full-length RdfS tetramer.
RdfS modelled with DNA. In all panels, the RdfS ASU tetramer is coloured in a blue-to-red rainbow from N- to C-terminal. (A) Visual representation of RdfS tetramer with an ensemble of fragments from crystallographically determined DNA for wHTH proteins: MtaN (red; PDB 1R8D), and filament forming wHTH proteins: BldC (blue; PDB 6AMK) and Xis (purple and green; 2IEF and 1RH6, respectively). Accompanying alignment to each structure individually can be found in Supplementary Fig. S1. (B) RdfS ASU tetramer modelled with nucleosome DNA (extracted from PDB 5GSE [72]) closely resembles the fragment ensemble model generated in panel (A). (C) Overlayed models of DNA-bound RdfS and RdfS13–89 initially prepared via molecular modelling based on BldC-DNA (PDB 6AMA). The additional α1 (not present in RdfS13–89) shown in magenta. (D) (i) Distance in the contact map from the target state with respect to the RdfS crystal structure and (ii) RMSD over the adiabatic biased molecular dynamics (first 100 ns) and the subsequent unbiased simulation (final 50 ns) for DNA-bound full length RdfS (black) and RdfS13–89 (red). (E) Overlay of the representative structure of DNA-bound RdfS from adiabatic biased molecular dynamics (with DNA shown in white) with the representative structure following unbiased molecular dynamics simulation (with DNA shown in grey). See in-text and supplementary data for further molecular dynamics analysis. (F) Overlay of the representative structure of DNA-bound RdfS13–89 from adiabatic-biased molecular dynamics (ABMD) (with DNA shown in white) with the representative structure following unbiased molecular dynamics simulation (with DNA shown in grey). (G) Minimized model of extended RdfS filament (16 total molecules) coloured with electrostatic surface, and DNA derived from unbiased molecular dynamics simulation shown in panel (E). RdfS α1 helix coloured in black and a magenta sphere marks the Cα of Pro66, showing the relative arrangement of the N- and C-termini of the RdfS superhelical structure.
RdfS–DNA complexes for RdfS and RdfS_13–89_ generated by homology modelling (Fig. 6C) were subjected to adiabatic-biased molecular dynamics (ABMD), during which RdfS monomer-monomer interactions were targeted to replicate those observed in RdfS ASU tetramer (see supplementary data for full details). RdfS–DNA complexes showed structural similarity (RMSD ∼0.3 nm) to the protein-only RdfS crystal structure, indicating successful targeting of this protein assembly using ABMD (Fig. 6D). Subsequent unbiased simulations following the ABMD simulations suggested the general stability of full-length RdfS in this complex, maintaining a shape with similar curvature to nucleosomal DNA (Fig. 6E). In contrast, the complex with RdfS_13–89_ was conformationally unstable in unbiased simulations, with the distal two RdfS_13–89_ molecules dislodging from the DNA (Fig. 6F), suggesting that the α1 helix is required for stable protein–protein and/or protein–DNA interactions. Extension of this 40-bp DNA model to a continuous RdfS superhelix highlights the possibility that RdfS and DNA can form an extensive polymeric complex (Fig. 6G), where the N-termini assemble along the superhelical axis of the elongated nucleoprotein and the C-termini face outward toward the external surface, with an overall left-handed twist consistent with negative supercoiling.
The N-terminal helix of RdfS is not essential for DNA-binding in vitro but is essential for in vivo activation of ICEMlSymR7A excision and conjugative transfer
Noting the importance of contacts involving the N-terminal α1 helix from RdfS, and its influence on stable DNA binding revealed by both the crystal structure and the molecular dynamics simulations, we decided to test experimentally if α1 was essential for oligomerization or DNA binding. A construct expressing a truncation of RdfS with residues 2–12 removed (RdfS_13–89_) was generated and the resulting protein purified. Analytical gel filtration experiments revealed RdfS_13–89_ was present mostly as dimers with an approximate molecular mass at ∼20 kDa (Fig. 5A). EMSAs were then carried out with RdfS_13–89_ against the attP region. A concentration of 1.3 μM RdfS_13–89_ was required to shift attP DNA, which is approximately twice the concentration required for the wild-type RdfS to shift the same DNA. Interestingly, we observed no significant reduction of the multi-levelled migration effect (Fig. 5E), suggesting that additional RdfS_13–89_ molecules can oligomerize independently of α1 when binding DNA. Therefore, it seems that removal of the α1 helix reduces protein oligomerization, but DNA binding propensity is reduced, and further oligomerization likely still occurs in conjunction with DNA. This was not surprising given other wHTH domains do not have the unique RdfS α1 helix and yet are capable of oligomerizing on DNA, and RdfS maintains the same, albeit smaller, oligomerization interface as these other proteins.
As RdfS_13–89_ was still capable of binding DNA in vitro, we wondered if an allele encoding this truncated version would be capable of activating ICEMlSym^R7A^ excision and conjugative transfer. Versions of the rdfS gene encoding N- and C-terminal truncations in RdfS, each positioned downstream of the native rdfS ribosome-binding site and PrdfS promoter, were constructed and cloned into plasmid pFAJ1700. To test the ability of these truncated alleles to stimulate ICEMlSym^R7A^ excision, we introduced these plasmids into M. japonicum R7A and used quantitative PCR assays to measure ICEMlSym^R7A^ excision. The R7AΔqseMΔrdfS background was used as it is derepressed for activation of ICEMlSym^R7A^ excision and transfer through deletion of the antiactivator gene qseM, but incapable of ICEMlSym^R7A^ excision or transfer due to the deletion of rdfS. R7AΔqseMΔrdfS cells containing empty-vector controls exhibited population excision frequencies <0.01% (Fig. 5F). When R7AΔqseMΔrdfS was used as a donor in conjugation experiments, transfer of ICEMlSym^R7A^ was not detected (detection limit of 1.3 × 10^−9^) (Supplementary Table S5). In the presence of a plasmid carrying the wild-type RdfS allele expressed from PrdfS (pJR202), the proportion of chromosomes with an excised ICEMlSym^R7A^ increased to ∼20% and conjugative transfer of ICEMlSym^R7A^ was restored to an average of 1.2 × 10^−4^ exconjugants per donor. In contrast, no restoration of excision or conjugative transfer was observed in the presence of α1-truncated RdfS_13–89_ expressed from pFJrdfS_13–89_. We wondered if overexpression of this rdfS13–89 allele might be able to compensate for the reduced DNA-binding exhibited by this protein; however, even when rdfS_13–89_ was expressed from the strong constitutive nptII promoter on pF08rdfS_13–89_, no restoration of excision or conjugative transfer of ICEMlSym^R7A^ was observed suggesting RdfS-mediated excision of ICEMlSym^R7A^ requires the RdfS α1-helix. We also constructed a separate plasmid coding for a version of RdfS deleted for the C-terminal 25 amino acids, RdfS_1–64_ (residues 64–89 are random coil or unresolved in the RdfS crystal structure). In contrast to the N-terminal truncation, C-terminally truncated version of RdfS restored excision and conjugation frequences to a similar level as full-length RdfS, suggesting that the disordered C-terminus of RdfS is not critical for excision and conjugation functions.
Discussion
The diverse molecular roles of the RdfS homologues encoded by Mesorhizobium spp. ICEs have been previously reported [19, 43, 44]; however, their structural basis has remained uncharacterized. In this work, we analysed the X-ray crystal structure of apo RdfS and discover that the left-handed superhelical filaments formed in crystals are also represented in solution where, at the concentrations tested, RdfS forms tetramers both in the presence and absence of DNA. The head-to-tail oligomerization of RdfS molecules is likely enhanced by the unique N-terminal α-helix not present in other characterized wHTH proteins. DNA-binding assays confirmed RdfS bound the ICEMlSym^R7A^attP site within a 40-bp region directly upstream of the putative IntS P3 arm-site, supporting its role as an RDF/excisionase for IntS-mediated ICEMlSym^R7A^ excision. RdfS binding to attP also activated transcription from the PintS promoter, which is likely silenced immediately following ICEMlSym^R7A^ excision through negative autoregulation by the IntS protein. DNA binding assays revealed RdfS also binds its own promoter PrdfS and thus negatively autoregulates its own expression. The apparent weaker affinity of RdfS for PrdfS likely facilitates the establishment of an equilibrium concentration of RdfS in transfer-active cells, which ensures ICEMlSym^R7A^ excision and conjugation functions are activated without causing the growth inhibition and ICEMlSym^R7A^-curing observed following artificial rdfS overexpression. Molecular dynamics simulations predicted the unique N-terminal α1-helix of RdfS was likely required for optimal oligomerization and compaction of higher-order RdfS–DNA complexes. Interestingly, removal of the α1-helix reduced DNA binding and oligomerization in vitro, but this same modification abolished all detectable activity of RdfS in vivo. In summary, the RdfS protein is a unique member of the wHTH family with a novel oligomer-stabilizing α1-helix. We propose that RdfS oligomerizes in a concentration-dependent manner and such oligomerization facilitates the stepwise binding to various target sites required for ICEMlSym^R7A^ excision and conjugation functions.
The identified RdfS target region attP_8, overlaps the presumed promoter region of intS. Our PintS-lacZ fusion experiments revealed RdfS activates intS transcription. In isolation it could be interpreted that the sole role of RdfS therefore is to transcriptionally activate IntS expression rather than participate as RDF. However, overexpression of IntS alone does not stimulate the observed ∼100% ICEMlSym^R7A^ excision when both RdfS and IntS are expressed together [19], confirming RdfS is required to stimulate excision directly as observed for other RDFs [15, 20, 73, 74]. Our experiments also showed IntS negatively regulates its own expression. This presumably prevents RdfS from overinducing PintS, which could shift the ratio of RdfS:IntS in favour of ICEMlSym^R7A^ integration. Therefore, our data are consistent with a model in which RdfS expression transiently activates IntS expression and then IntS and RdfS together excise ICEMlSym^R7A^, after which IntS expression is inhibited by IntS, which a ratio of RdfS:IntS that maintains ICEMlSym^R7A^ in an excised state in preparation for conjugative transfer.
Other RDFs such P2 Cox [75], phage 186 Apl [76], P4 Vis [77], WΦ Cox [70], and P22 Cox [78] form higher-order complexes on att DNA. The increasing DNA retardation with RdfS concentration observed in EMSAs here supports RdfS forming variable-length multimers along the attP DNA region. Our SPR experiments revealed RdfS exhibits specificity for a 40-bp region attP_8 and smaller portions of this DNA region were not bound as strongly, suggesting the natural footprint for RdfS binding may be close to this size (at least at this site). SEC-SY-SAXS analysis of 6H-RdfS indicated a stable complex with attP_8 as a tetramer (4 RdfS: 1 attP_8 dsDNA), with one RdfS molecule per 10-bp of DNA. Preparations of purified RdfS tended to precipitate at higher concentrations, suggesting larger oligomers may form in solution but are unstable in the conditions tested. It seems possible that in more suitable conditions RdfS may form larger multimers together with longer DNA regions.
Much like RdfS, the P2 bacteriophage RDF Cox_P2_ is a wHTH domain protein that oligomerizes in vitro and its oligomerization is essential for its roles in P2 recombination and regulation [75]. However, Cox_P2_ encodes an additional C-terminal helix beyond the corresponding α4 helix in RdfS (Supplementary Fig. S6). The two C-terminal α-helices of Cox_P2_ enable it to form a continuous left-handed superhelical structure in crystals. This arrangement of Cox_P2_ enables the formation of oligomeric filaments which compact on DNA, mimicking histone architecture [79]. It seems likely that the C-terminal helices of Cox_P2_ and the N-terminal α1 helix of RdfS function in an analogous manner, enabling protein–protein contacts along DNA to compact and transform overall morphology of the nucleoprotein filament. Much like the removal of the α1 helix of RdfS, mutations within the C-terminal helices of Cox_P2_ abolish biological activity [24]. It seems possible that like the Cox_P2_ C-terminal helices, the RdfS α1 helix may facilitate nucleoprotein contraction and that this contraction may be critical to its functions as an RDF and a transcription factor. Such DNA compaction is supported by our molecular dynamic simulations (Fig. 6), although remains experimentally untested.
Despite identifying several DNA-binding targets for RdfS, we were unable to identify consensus binding motifs. Our SPR experiments also indicated no individual nucleotide changes abolished RdfS binding to attP_8, although most changes reduced the binding response. The Streptomyces BldC protein is a wHTH-domain regulator with structural similarity to ICEMlSym^R7A^ RdfS (Fig. 1). BldC binds multiple DNA targets and oligomerizes in a head-to-tail manner. Whilst BldC can bind a 22-bp DNA-target with 9-bp direct repeats (whiI) as a dimer, the mechanism of binding to the larger pseudo-continuous smeA-sffA promoter differs. BldC cooperatively forms extended filaments along the smeA-sffA region with one DNA-turn between each bound BldC molecule [36]. This resembles our proposed model of DNA binding by RdfS (Supplementary Fig. S6). Interestingly, binding of BldC to the smeA-sffA region appears not to be strictly dependent on DNA sequence-consensus, suggesting the target DNA structure could be more crucial than sequence—a phenomenon referred to as indirect readout [80, 81]. A similar indirect readout type of DNA-binding mechanism is described for another BldC orthologue from Actinoplanes missouriensis, which binds AT-rich regions [82], and for phage λ Xis which forms cooperative filaments along DNA [69]. Unlike RdfS however, Streptomyces BldC remains a monomer in solution and the multimeric superhelical behaviour is dependent on the presence of DNA—indicating BldC must be anchored to its cognate DNA to assemble into higher-order oligomers [36]. BldC primarily forms its head-to-tail oligomers due to its hydrophobic core and cross-contacts between Glu and Arg between protomers at the β wing-HTH region. Although the relative position of these residues differs in RdfS, equivalent contacts are made at the β wing-HTH, although many more RdfS intermolecular contacts are made with the α1 (Fig. 2B and C). Our analytical SEC and molecular dynamics suggest that RdfS oligomerization is mediated independent of DNA and RdfS–DNA binding stability is enhanced by the N-terminal α1 helix. Our data here indicate that RdfS can oligomerize in solution, so it seems possible that RdfS may oligomerize in situ prior to DNA binding and as such, have the capability to recognize larger and more dispersed DNA recognition motifs.
Evolving a capacity to recognize larger DNA recognition sites would presumably afford RdfS with a larger DNA footprint within which binding specificity could evolve. Such a capability may have been critical for the evolutionary success of ICEMlSym^R7A^ in competition with the multitude of related ICEs present in the genus Mesorhizobium. Proteins closely related to RdfS are encoded by conjugative elements throughout the genus Mesorhizobium. Indeed, all identified ICEs (average 1.2 ICEs per genome [14]) and most identified conjugative plasmids in Mesorhizobium carry a homologue of RdfS encoded in a similar genetic context upstream of the transfer genes traF and rlxS. Phylogenetic comparisons of the core conjugation/transfer genes of all Mesorhizobium ICEs places all symbiosis-gene carrying ICEs (both monopartite and tripartite) into a monophyletic group [14], which we have named the ICESym family. Mesorhizobium ICEs outside this clade exhibit a more diverse evolutionary history but nonetheless carry the same set of core conjugation/transfer genes, including rdfS. Strikingly, we observed here that only RdfS homologues encoded by the ICESym family contain the α1 helix (Supplementary Fig. S7). All other RdfS homologues encoded by Mesorhizobium ICEs and plasmids, while exhibiting ∼61% similarity over the wHTH domain region with ICEMlSym^R7A^ RdfS, do not encode the α1 helix. We speculate therefore that evolution of this helix coincided with the expansion and dissemination of the ICESym family throughout the genus Mesorhizobium and that the additional RdfS helix may have reduced regulatory interference from other RdfS homologues present in Mesorhizobium genomes. This model is also consistent with the broader evolutionary distribution of ICESym family ICEs compared to other ICE families in the genus [14, 83].
Several lines of evidence point to RdfS having a role in the regulation of conjugation. The position of the rdfS gene upstream of traF and rlxS conjugation genes is conserved across related ICEs and plasmids throughout the proteobacteria [14, 19]. This contrasts with nearly all other identified RDF genes, which are typically located adjacent to their cognate integrase gene [20]. Deletion of rdfS also abolishes ICEMlSym^R7A^ conjugation; however, this is not surprising given the requirement for ICEMlSym^R7A^ excision. More surprising is that the rdfS deletion also abolishes the ability of the ICEMlSym^R7A^ conjugation system to mobilize a plasmid carrying the ICEMlSym^R7A^oriT, a process which does not physically require ICEMlSym^R7A^ to excise. This suggests RdfS more directly activates conjugation functions. Furthermore, in these experiments, low-level expression of rdfS from the lac promoter restored ICEMlSym^R7A^ excision but did not restore conjugation [43], suggesting conjugation might require a greater level of rdfS expression. In conjugation experiments here, we introduced rdfS expressed from its native promoter and in contrast to our previous work, this restored both ICEMlSym^R7A^ excision and conjugation. These observations suggest that a greater concentration of RdfS is required to activate conjugation than stimulate excision. We speculate that the RdfS regulon extends beyond the attP and PrdfS/PrdfG regions identified in this work and that at higher concentrations RdfS activates or derepresses additional ICE genes more directly involved in conjugative transfer. The concentration-dependent binding and oligomerization of RdfS with various DNA targets described in this work fits well with such a model. In future experiments, genome-wide protein–DNA interaction assays such as ChIP-Seq in conditions of transient RdfS overexpression may unveil additional targets of the multifunctional RdfS.
Supplementary Material
gkaf249_Supplemental_File
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Beaber JW, Hochhut B, Waldor MK Genomic and functional analyses of SXT, an integrating antibiotic resistance gene transfer element derived from Vibrio cholerae. J Bacteriol. 2002; 184:4259–69.10.1128/JB.184.15.4259-4269.2002.12107144 PMC 135215 · doi ↗ · pubmed ↗
- 2Drenkard E, Ausubel FM Pseudomonas biofilm formation and antibiotic resistance are linked to phenotypic variation. Nature. 2002; 416:740–3.10.1038/416740 a.11961556 · doi ↗ · pubmed ↗
- 3Peters SE, Hobman JL, Strike P et al. Novel mercury resistance determinants carried by Inc J plasmids p MERPH and R 391. Molec Gen Genet. 1991; 228:294–9.10.1007/BF 00282479.1886614 · doi ↗ · pubmed ↗
- 4Ravatn R, Studer S, Springael D et al. Chromosomal integration, tandem amplification, and deamplification in Pseudomonas putida F 1 of a 105-kilobase genetic element containing the chlorocatechol degradative genes from Pseudomonas sp. Strain B 13. J Bacteriol. 1998; 180:4360–9.10.1128/JB.180.17.4360-4369.1998.9721270 PMC 107442 · doi ↗ · pubmed ↗
- 5Finan TM Evolving insights: symbiosis islands and horizontal gene transfer. J Bacteriol. 2002; 184:2855–6.10.1128/JB.184.11.2855-2856.2002.12003923 PMC 135049 · doi ↗ · pubmed ↗
- 6Sullivan J, Ronson C Evolution of rhizobia by acquisition of a 500-kb symbiosis island that integrates into a phe-t RNA gene. Proc Natl Acad Sci USA. 1998; 95:5145–9.10.1073/pnas.95.9.5145.9560243 PMC 20228 · doi ↗ · pubmed ↗
- 7Burrus V, Marrero J, Waldor MK The current ICE age: biology and evolution of SXT-related integrating conjugative elements. Plasmid. 2006; 55:173–83.10.1016/j.plasmid.2006.01.001.16530834 · doi ↗ · pubmed ↗
- 8Dimopoulou ID, Kartali SI, Harding RM et al. Diversity of antibiotic resistance integrative and conjugative elements among haemophili. J Med Microbiol. 2007; 56:838–46.10.1099/jmm.0.47125-0.17510272 · doi ↗ · pubmed ↗