A commonly used anti-SOX15 antibody fails to demonstrate specificity in mouse embryos
Farina Aziz, Bin Gu, Amy Ralston

TL;DR
A commonly used antibody for detecting SOX15 lacks specificity in mouse embryos, even when SOX15 is absent.
Contribution
The study reveals that a commercially available anti-SOX15 antibody is not specific in mouse embryos.
Findings
Nuclear signal was detected in both wild-type and Sox15-null embryos.
The antibody's signal persisted even when maternal or paternal Sox15 contributions were eliminated.
Abstract
SOX15 is a broadly conserved transcription factor involved in many critical processes, including mammalian cell fate specification and pluripotency. We investigated the specificity of a commercially available, polyclonal anti-SOX15 antibody advertised as knockout-validated. We generated a new mouse line carrying a null allele of Sox15 , and then evaluated anti-SOX15 activity in Sox15 null mouse embryos. Nuclear signal was detected in both wild type and null embryos, even when potential maternally or paternally-contributed Sox15 was eliminated. We conclude that this SOX15-detecting reagent may not be suitable for all applications, and caution the growing community of users accordingly.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
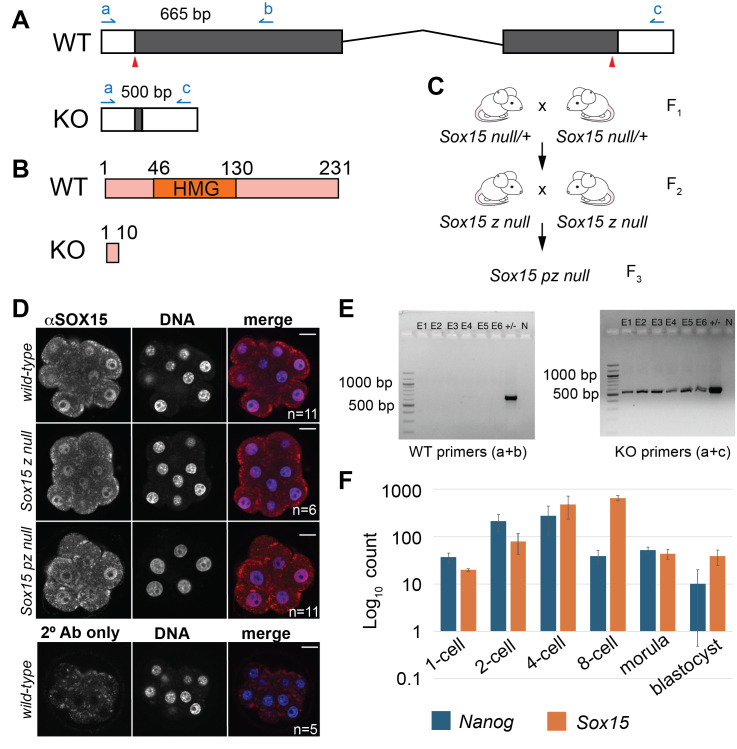 Figure 1
Figure 1- —Eunice Kennedy Shriver National Institute of Child Health and Human Development (United States)https://ror.org/04byxyr05
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAnimal Genetics and Reproduction · Cell Adhesion Molecules Research
Description
SOX proteins are SRY-related HMG-box transcription factors that play essential roles in many biological contexts (Kamachi & Kondoh, 2013; Sarkar & Hochedlinger, 2013). The Group G SOX encoded by * Sox15 * has been implicated in placental development (Ito, 2010; Yamada et al., 2008), neurogenesis (Choi et al., 2023), muscle regeneration (Lee et al., 2004; Savage et al., 2009) and cancer (Hu et al., 2024; Moradi et al., 2017). Additionally, SOX15 expression is enriched in mouse embryonic stem (ES) cells, where it helps promote pluripotency (Choi et al., 2023; Maruyama et al., 2005). ES cells are derived from the mouse embryo during preimplantation, when pluripotency is first established. Yet the SOX15 expression pattern in preimplantation mouse embryos has not been reported.
To investigate SOX15 expression in mouse embryos, we selected an anti-SOX15 antibody that had previously been knockout-validated in mouse ES cells (Choi et al., 2023). That is, a protein of the predicted size of SOX15 was detected in wild type mouse ES cells by Western blot using a polyclonal antibody (25415-1-AP) raised against amino acids 122-231 of SOX15, but not in ES cells carrying two alleles encoding the N-terminal 72 amino acids of SOX15. To ensure that we would also be able to validate this antibody in embryos, we generated a new mouse line carrying a * Sox15 * knockout allele, predicted to encode the first 10 amino acids of * Sox15 * ( Fig. 1A, B), by CRISPR/Cas9-mediated genome editing.
We then bred mice carrying the * Sox15 * knockout allele to generate preimplantation embryos that were lacking zygotically transcribed * Sox15 * ( Fig. 1C ). Because homozygous * Sox15 * knockout mice are viable and fertile (Lee et al., 2004; Maruyama et al., 2005), we also generated * Sox15 * null adults. These * Sox15 * null adults were subsequently intercrossed to produce embryos lacking * Sox15 * mRNA or protein potentially carried by gametes, as well as embryo-expressed * Sox15 * .
We then evaluated anti-SOX15 activity in preimplantation embryos by immunofluorescence and confocal microscopy ( Fig. 1D, E). During preimplantation, * Sox15 * mRNA is at least as abundant as * Nanog * ( Fig. 1F ), and NANOG is robustly detected by immunofluorescence during preimplantation (Strumpf et al., 2005). This provided confidence that SOX15 should be detectable by immunofluorescence during preimplantation. In fact, the anti-SOX15 antibody detected a nuclear signal in wild type embryos ( Fig. 1D ). However, this signal persisted with no apparent reduction in * Sox15 * null embryos, indicating that the antibody fails to accurately and specifically detect SOX15 in mouse embryos.
This finding underscores the need for characterization of antibody performance in specific applications (Kahn et al., 2024). We note that lot-to-lot variation could also impact results, but lot numbers have not been reported in publications that may have used this reagent for immunofluorescence or other application (Belair-Hickey et al., 2024; Choi et al., 2023; Hu et al., 2024; Liu et al., 2022). Increased transparency and reporting standards during publishing could positively impact the rigor of antibody-dependent studies in general (Taussig et al., 2018). Databases, technologies, and high throughput strategies to curtail the “antibody characterization crisis” have also been described (Ayoubi et al., 2024; Kahn et al., 2024). Ultimately, to ensure accurate characterization of SOX15, future studies should incorporate alternative well-characterized antibodies or orthogonal detection methods, such as knock-in reporters or proteomic approaches.
Methods
All animal research was conducted in accordance with the guidelines of the Michigan State University Institutional Animal Care and Use Committee. Mice were maintained on a 12 hr light/dark cycle.
The * Sox15 * null allele was generated by injecting sgRNAs and CAS9 enzyme sgRNAs into the pronucleus of zygotes and then transferring to recipient females following standard practices.
Mouse genotypes were determined by PCR using genomic DNA extracted from ear biopsy using the REDExtract-N-Amp kit. Embryo genomic DNA was extracted using the same kit, in 10 µl total volume. Genomic extracts (1-2 µl) were then subjected to PCR using allele-specific primers. PCR conditions were: 95 °C for 5 minutes, 30 cycles of 95 °C for 30 seconds, 57 °C for 1 minute, 68 °C (30 seconds KO and 40 seconds WT), and 68 °C for 5 minutes.
Embryos were collected by flushing the oviduct or uterus with M2 medium. Embryos were then incubated in 4% formaldehyde for 10 minutes at room temperature, 0.5% Triton X-100 in PBS for 30 minutes at room temperature, blocking solution (10% Fetal Bovine Serum, 0.1% Triton X-100 in PBS) overnight at 4 °C, primary antibody (anti-rabbit anti-SOX15, 1:400 in blocking solution) overnight at 4 ºC, blocking solution for 30 minutes at room temperature, 2º antibody (Donkey-anti-rabbit IgG Alexa 546, 1:400 in blocking solution) for 1 hour, blocking solution for 30 minutes at room temperature, and finally imaged in DRAQ5 1:400 in blocking solution.
Embryos were imaged using an Olympus FluoView FV1000 Confocal Laser Scanning Microscope system with 60× PlanApoN oil (NA 1.42) objective. For each embryo, z -stacks were collected, with 5 µm intervals between optical sections. All embryos were imaged before knowledge of their genotypes. Genotypes were determined by PCR after imaging as described above.
Reagents
CD-1 mice (Jackson Labs)
Fetal Bovine Serum (Hyclone)
M2 Medium (Millipore)
Formaldehyde (Polysciences)
Triton X-100 (Sigma Aldrich)
PBS (Sigma Aldrich)
DRAQ5 (Cell Signaling Technologies)
Rabbit anti-SOX15 (25415-1-AP, ProteinTech, lot 00082448)
Donkey anti-rabbit IgG Alexa 546 (Invitrogen, A10040)
REDExtract-N-Amp Kit (Sigma-Aldrich)
sgRNAs (Synthego): GCTCCTCACAAGCAGAGACT, AAGGGAAGTATTATATGGAG
Allele-specific primers (IDT): a) GCAGCTGTTGGGACTTTGTG, b) TCGGTACTTGTAGTCGGGATAG, c) GTTGCTGCTGTAGGGAGAGAATAC (see Fig. 1).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ayoubi R Ryan J Gonzalez Bolivar S Alende C Ruiz Moleon V Fotouhi M Alqazzaz M Southern K Alshafie W Baker MR Ball AR Jr Callahan D Cooper JA Crosby K Harvey KJ Houston DW Kumaran R Rego M Schofield C Wu H Biddle MS Brown CM Kahn RA Bandrowski A Virk HS Edwards AM Mc Pherson PS Laflamme C 20241217 A consensus platform for antibody characterization.Nat Protoc 10.1038/s 41596-024-01095-839690206 PMC 13054593 · doi ↗ · pubmed ↗
- 2Belair-Hickey Justin J. Khalili Saeed Coles Brenda L.K. Ballios Brian G. Liu Jeff C. Bader Gary D. van der Kooy Derek 20241216 Sox 15 is a novel retinal developmental gene that promotes cone photoreceptor differentiation through inhibition of alternative rod photoreceptor fates 10.1101/2024.12.10.627788 · doi ↗
- 3Choi EB Vodnala M Saini P Anugula S Zerbato M Ho JJ Wang J Ho Sui SJ Yoon J Roels M Inouye C Fong YW 202329 Transcription factor SOX 15 regulates stem cell pluripotency and promotes neural fate during differentiation by activating the neurogenic gene Hes 5.J Biol Chem 29930021-925810299610299610.1016/j.jbc.2023.10299636764520 PMC 10023989 · doi ↗ · pubmed ↗
- 4Hu J Li X Xu K Chen J Zong S Zhang H Li H Zhang G Guo Z Zhao X Jiang Y Jing Z 202484 Circ VPS 8 promotes the malignant phenotype and inhibits ferroptosis of glioma stem cells by acting as a scaffold for MKRN 1, SOX 15 and HNF 4A.Oncogene 43360950-92322679269510.1038/s 41388-024-03116-y 39098847 · doi ↗ · pubmed ↗
- 5Ito Michihiko 201031 Function and molecular evolution of mammalian Sox 15, a singleton in the Sox G group of transcription factors The International Journal of Biochemistry & Cell Biology 4231357-272544945210.1016/j.biocel.2009.10.02319909824 · doi ↗ · pubmed ↗
- 6Kahn RA Virk H Laflamme C Houston DW Polinski NK Meijers R Levey AI Saper CB Errington TM Turn RE Bandrowski A Trimmer JS Rego M Freedman LP Ferrara F Bradbury ARM Cable H Longworth S 2024814 Antibody characterization is critical to enhance reproducibility in biomedical research.Elife 1310.7554/e Life.10021139140332 PMC 11324233 · doi ↗ · pubmed ↗
- 7Kamachi Yusuke Kondoh Hisato 20131015 Sox proteins: regulators of cell fate specification and differentiation Development 140201477-91294129414410.1242/dev.09179324086078 · doi ↗ · pubmed ↗
- 8Lee HJ Göring W Ochs M Mühlfeld C Steding G Paprotta I Engel W Adham IM 2004101 Sox 15 is required for skeletal muscle regeneration.Mol Cell Biol 24190270-73068428843610.1128/MCB.24.19.8428-8436.200415367664 PMC 516755 · doi ↗ · pubmed ↗