Characterisation of Enterocins Produced by Antilisterial Enterococcus faecium BH04, BH12, BH84, and BH99 and In Vitro/In Situ Inhibition of Listeria monocytogenes
Haktan Aktaş

TL;DR
Researchers studied bacteriocins from Enterococcus faecium strains to inhibit Listeria monocytogenes, a dangerous foodborne pathogen.
Contribution
The study identifies and characterizes antilisterial enterocins from four E. faecium strains and evaluates their stability and effectiveness.
Findings
E. faecium strains BH04, BH12, BH84, and BH99 showed strong bacteriocin activity against Listeria monocytogenes.
Enterocins retained antilisterial activity under heat, pH changes, and chemical exposure but were reduced by UV light.
The strains could be used as co-cultures in fermented foods to control Listeria.
Abstract
Listeria monocytogenes is a food pathogen that can cause fatal infections, especially in newborns, elderly individuals, and immunocompromised individuals. In recent years, research on novel antibiotics and natural antimicrobial agents of plant or microbial origin has become increasingly important in the face of multiple antibiotic resistance. In this study, enterocins produced by Enterococcus faecium BH04, BH12, BH84, and BH99 were investigated in vitro and in situ as antilisterial agents. The results showed that all strains had bacteriocin activity of 0.4 kAU/mL (400 AU/mL) after 3 h and 25.6 kAU/mL (25600 AU/mL) after 6 h in M17 broth. The strains inhibited the growth of Listeria monocytogenes ATCC 7644 in BHI. Furthermore, E. faecium BH04, BH12, BH84, and BH99 had bacteriostatic potential, whereas enterocins BH04, BH12, BH84, and BH99 had bactericidal potential in a milk model…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
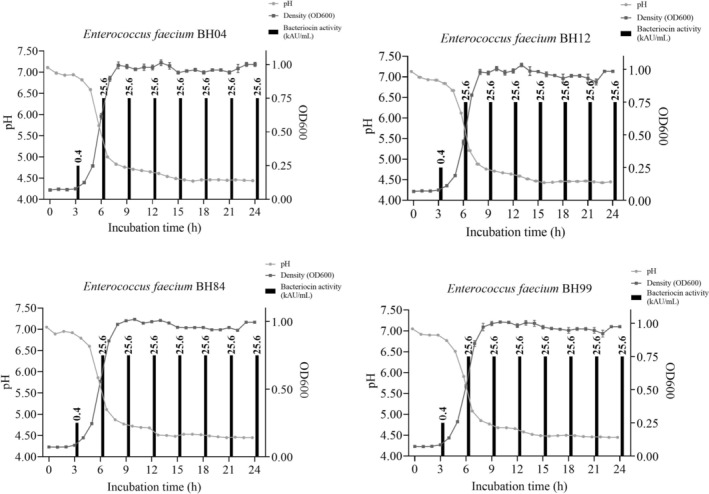 FIGURE 1
FIGURE 1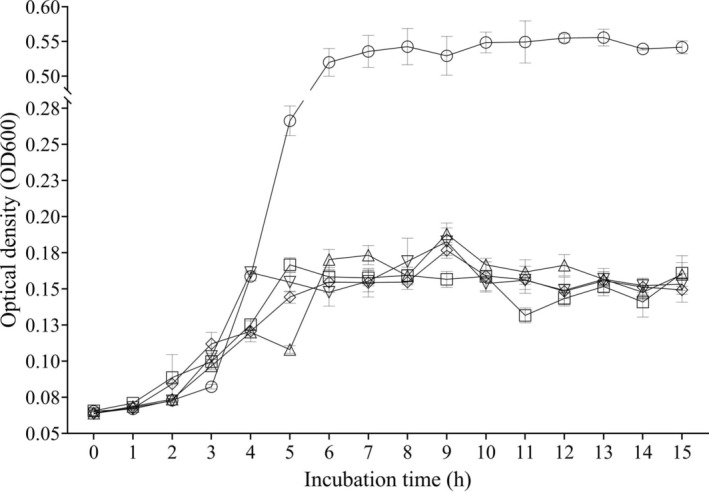 FIGURE 2
FIGURE 2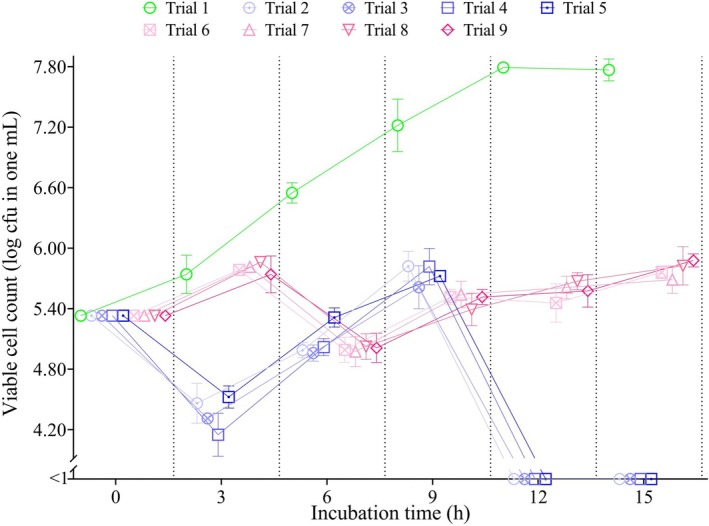 FIGURE 3
FIGURE 3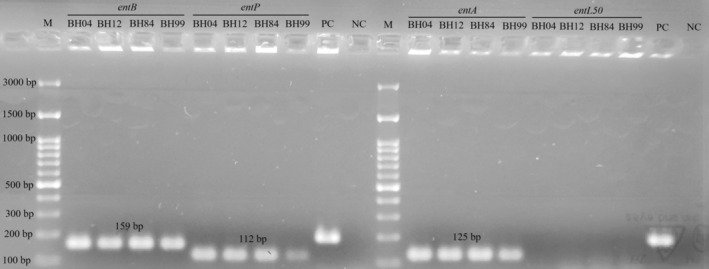 FIGURE 4
FIGURE 4 FIGURE 5
FIGURE 5| Isolates | Source | Growth conditions |
| |||
|---|---|---|---|---|---|---|
|
|
|
|
| |||
|
| Çetin and Aktaş ( | M17, 37°C | ND | — | — | — |
|
| Çetin and Aktaş ( | M17, 37°C | — | ND | — | — |
|
| Çetin and Aktaş ( | M17, 37°C | — | — | ND | — |
|
| Çetin and Aktaş ( | M17, 37°C | — | — | — | ND |
|
| ATCC 7644 | BHIA, 37°C | + (20.5 ± 0.7) | + (20.3 ± 0.4) | + (19.3 ± 0.4) | + (19.8 ± 0.4) |
|
| ATCC 29213 | NA, 37°C | — | — | — | — |
| Methicillin‐resistant | ATCC 43300 | NA, 37°C | — | — | — | — |
|
| ATCC 25922 | NA, 37°C | — | — | — | — |
|
| RSHMB 95091 | NA, 37°C | — | — | — | — |
|
| ATCC 6633 | BHIA, 30°C | — | — | — | — |
|
| ATCC 33019 | NA, 30°C | — | — | — | — |
|
| ATCC 7050 | NA, 30°C | + (12.5 ± 0.7) | + (10.0 ± 0.0) | + (11.5 ± 0.7) | + (12.3 ± 0.4) |
|
| ATCC 25175 | BHIA, 37°C | — | — | — | — |
|
| ATCC 29212 | M17, 37°C | — | — | — | — |
|
| Aktaş and Çetin ( | M17, 37°C | — | — | — | — |
|
| Aktaş ( | MRS, 37°C | — | — | + (21.0 + 1.4) | + (21.0 + 1.4) |
|
| ATCC 15009 | MRS, 37°C | — | — | — | — |
|
| ATCC 393 | MRS, 37°C | — | — | — | — |
| The primer name | Sequence (5′‐3′) | Target gene | Annealing temperature (°C) | Amplicon size (bp) | Reference |
|---|---|---|---|---|---|
| Enterocin A | F: AAATATTATGGAAATGGAGTGTAT |
| 55.5 | 125 | Du Toit et al. ( |
| R: GCACTTCCCTGGAATTGCTC | |||||
| Enterocin B | F: GAAAATGATCACAGAATGCCTA |
| 54.5 | 159 | |
| R: GTTGCATTTAGAGTATACATTTG | |||||
| Enterocin P | F: TATGGTAATGGTGTTTATTGTAAT |
| 54.5 | 112 | |
| R: ATGTCCCATACCTGCCAAAC | |||||
| Enterocin L50 | F: STGGGAGCAATCGCAAAATTAG |
| 56.5 | 135 | |
| R: ATTGCCCATCCTTCTCCAAT |
| Treatment | Time (min) | Residual activity (%) | |||
|---|---|---|---|---|---|
| Enterocin BH04 | Enterocin BH12 | Enterocin BH84 | Enterocin BH99 | ||
| Temperature | |||||
| −80°C | 10 | 100 | 100 | 100 | 100 |
| 30 | 100 | 100 | 100 | 100 | |
| 60 | 100 | 100 | 100 | 100 | |
| −20°C | 10 | 100 | 100 | 100 | 100 |
| 30 | 100 | 100 | 100 | 100 | |
| 60 | 100 | 100 | 100 | 100 | |
| 4°C | 10 | 100 | 100 | 100 | 100 |
| 30 | 100 | 100 | 100 | 100 | |
| 60 | 100 | 100 | 100 | 100 | |
| 37°C | 10 | 100 | 100 | 100 | 100 |
| 30 | 100 | 100 | 100 | 100 | |
| 60 | 100 | 100 | 100 | 100 | |
| 60°C | 10 | 100 | 100 | 100 | 100 |
| 30 | 100 | 100 | 100 | 100 | |
| 60 | 100 | 100 | 100 | 100 | |
| 80°C | 10 | 100 | 100 | 100 | 100 |
| 30 | 100 | 100 | 100 | 100 | |
| 60 | 100 | 100 | 100 | 100 | |
| 100°C | 10 | 100 | 100 | 100 | 100 |
| 30 | 50 | 50 | 50 | 50 | |
| 60 | 50 | 50 | 50 | 50 | |
| 121°C | 15 | 6.25 | 3.125 | 6.25 | 6.25 |
| pH | |||||
| 2 | 100 | 100 | 100 | 100 | |
| 4 | 100 | 100 | 100 | 100 | |
| 6 | 100 | 100 | 100 | 100 | |
| 8 | 100 | 100 | 100 | 100 | |
| 10 | 100 | 100 | 100 | 100 | |
| 12 | 12.5 | 12.5 | 6.25 | 6.25 | |
| Enzyme (1 mg/mL) | |||||
| Catalase | 100 | 100 | 100 | 100 | |
| Lysozyme | 100 | 100 | 100 | 100 | |
| Pepsin | 100 | 25 | 12.5 | 6.25 | |
| Proteinase K | 0 | 0 | 0 | 0 | |
| Trypsin | 0 | 0 | 0 | 0 | |
| Chemicals (1%, v/v) | |||||
| 2‐mercaptoethanol | 100 | 100 | 100 | 100 | |
| Acetone | 100 | 100 | 100 | 100 | |
| Chloroform | 100 | 100 | 100 | 100 | |
| Ethanol | 400 | 400 | 200 | 200 | |
| Ethylenediaminetetraacetic acid (EDTA) | 100 | 100 | 100 | 100 | |
| Glycerol | 100 | 100 | 100 | 100 | |
| Methanol | 200 | 200 | 200 | 400 | |
| Reconstitute skim milk | 100 | 100 | 100 | 100 | |
| Sodium chloride (NaCl) | 100 | 100 | 100 | 100 | |
| Sodium dodecyl sulfate (SDS) | 200 | 400 | 400 | 200 | |
| Triton X‐100 | 100 | 100 | 100 | 100 | |
| Tween‐20 | 100 | 100 | 100 | 100 | |
| Tween‐80 | 100 | 100 | 100 | 100 | |
| Urea | 100 | 100 | 100 | 100 | |
| UV (30 cm, 5 min) | |||||
| UV | 50 | 25 | 50 | 50 | |
- —Atatürk Üniversitesi 10.13039/501100004951
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProbiotics and Fermented Foods · Listeria monocytogenes in Food Safety · Microbial Inactivation Methods
Introduction
1
Foodborne pathogens such as Listeria monocytogenes ( L. monocytogenes ) have the potential to threaten food safety and adversely affect human health by causing many diseases (FAO/WHO 2021). L. monocytogenes , one of the most important foodborne pathogens, can cause some serious diseases, including septicaemia, meningitis, and foetal loss (Tauxe 2002). While the mortality rate of listeriosis was 25% at the beginning of the twenty‐first century (Tauxe 2002), recent reports unfortunately indicate that this rate has not decreased (Matsuda et al. 2023). This situation demonstrates the need for effective antibiotics and/or natural antimicrobials (Church and McKillip 2021). Furthermore, as antibiotic‐resistant bacterial infections are thought to cause too many deaths each year (e.g., 23,000 deaths in the United States alone), natural antimicrobial agents such as bacteriocins and plant extracts have attracted considerable interest in recent years to combat pathogens (Chen et al. 2024). Bacteriocins, such as enterocin, lactacin, brevicin, caseicin, lacticin, and reutericin, can be produced by many lactic acid bacteria (LAB) (Klaenhammer 1993) and offer important food safety advantages because they are safe, nontoxic to humans, generally heat and pH stable, and have a broad antimicrobial spectrum including many pathogens (Gálvez et al. 2007). For these reasons, it is essential to study and characterize antilisterial bacteriocins produced by LAB, especially to reduce the virulence potential of L. monocytogenes .
This study aimed to characterize the enterocins produced by antilisterial Enterococcus faecium ( E. faecium ) BH04, BH12, BH84, and BH99 in terms of several aspects, including antibacterial spectrum, detection of enterocin genes, sequencing, and determination of residual activity against many treatments such as temperature, pH, enzyme, chemical, and ultraviolet (UV) light. In addition, the in vitro and in situ inhibitory effects of the enterocins on L. monocytogenes ATCC 7644 were investigated.
Materials and Methods
2
Strains and Growth Conditions
2.1
In this study, the bacteriocinogenic E. faecium BH04, BH12, BH84, and BH99, which were isolated from raw cow's milk and were determined to have bacteriocin‐induced antimicrobial activity in a previous study by Çetin and Aktaş (2024), were used as enterocin‐producing strains. The 16S rRNA identification, probiotic characterization, and safety assessment results of these isolates are presented in Table S1. The incubation conditions of the strains in this study were on M17 agar or broth (Merck, Darmstadt, Germany) at 37°C for 24 h. In addition, all other isolates used in this study and their growth conditions are listed in Table 1.
TABLE 1: Antibacterial spectrum of the enterocins produced by the Enterococcus spp. on some pathogens and nonpathogen bacteria a .
Obtaining Enterocins by Antilisterial Enterococcus Spp.
2.2
The strains including E. faecium BH04, BH12, BH84, and BH99, stored at −80°C in 40% glycerol solution (Merck), were plated on M17 agar (Merck) and incubated at 37°C for 24 h for revival. The fresh cultures were then suspended in phosphate‐buffered saline (PBS, pH 7.2, Merck) at a concentration of 10^8^ cfu/mL. The suspension was added (2%, v/v) to M17 broth (Merck) containing 0.5% glucose (Merck) and incubated at 37°C for 15–18 h. Then, the pH of the broth was adjusted to 6.5–7.0 with 1 N NaOH (Merck) to eliminate acidity, and the mixture was centrifuged at 8000 × g for 10 min at 4°C using an Allegra X‐30R Centrifuge (Beckman Coulter Inc., Brea, California, USA). The cell‐free supernatant (CFS) containing enterocin was filtered through a 0.22 μm pore size cellulose acetate filter (Merck) and the catalase enzyme (Sigma‐Aldrich, St. Louis, Missouri, USA) was added (5 mg/mL) to the CFS to eliminate the antimicrobial activity of H_2_O_2_ (Sigma‐Aldrich) (Ogunbanwo et al. 2003).
Microbial Growth Curve and Bacteriocin Activity Against
Listeria monocytogenes ATCC 7644
2.3
M17 broth (Merck) containing 0.5% glucose (Merck) was used to create a microbial growth curve and to determine bacteriocin activity (against L. monocytogenes ATCC 7644) of E. faecium BH04, BH12, BH84, and BH99 strains. Briefly, an overnight culture was suspended in PBS (Merck) at a concentration of 10^8^ cfu/mL, and the suspension (2%) was added to M17 broth (Merck) containing 0.5% glucose (Merck). After pre‐incubation at 37°C for 18–24 h, the broth (2%) was inoculated again into M17 broth (Merck) containing 0.5% glucose (Merck). The broths were incubated at 37°C for 24 h, and the pH and optical density (OD_600_) of the broths were determined every hour to generate a microbial growth curve. In addition, CFS were harvested from M17 broth (Merck) every 3 h of incubation. The CFS were then serially diluted (two‐fold) with PBS (Merck) and 10 μL of each dilution was spotted onto Brain Heart Infusion Agar (BHIA, Sigma‐Aldrich) inoculated with L. monocytogenes ATCC 7644 using a sterile cotton swab. Using the last dilution that inhibited the growth of L. monocytogenes ATCC 7644, the enterocin activities of the strains were determined using the formula (two‐fold serial dilution method): Bacteriocin activity (AU/mL) = 2^n^ × 100, where n represents the last dilution that inhibited the growth of L. monocytogenes ATCC 7644 (Todorov and Dicks 2007; Murua et al. 2013).
Antibacterial Spectrum of Enterocins
2.4
The antibacterial spectrum of the enterocins produced by the Enterococcus spp. was determined using the well diffusion method on Gram‐positive and Gram‐negative bacteria listed in Table 1. Briefly, overnight cultures in Table 1 were suspended in PBS (Merck) at a concentration of 10^8^ cfu/mL and spread over the entire agar surface using a sterile cotton swab. Holes of 6 mm diameter were then punched aseptically using a sterile cork borer. One hundred microliters of the CFS were inoculated into the wells. The plates were incubated under the appropriate conditions listed in Table 1. At the end of the incubation, the diameter of the zone around the well was determined and expressed in millimeters (mm) (Balouiri et al. 2016).
In Vitro (Mode of Action) and In Situ (Milk Model) Inhibition of
Listeria monocytogenes
2.5
To determine in vitro inhibition activity of the enterocins, 20 mL of each CFS from the Enterococcus strains was added to BHI broth (150 mL) (Sigma‐Aldrich) containing L. monocytogenes ATCC 7644 in the early exponential phase (3 h). The medium was incubated at 37°C for 15 h, and the absorbance values (OD_600_) of the medium were determined every hour using a plate reader (Epoch, BioTek, Agilent Technologies Inc., Santa Clara, California, USA). Medium without the CFS was used as a control (Pinto et al. 2009).
Nine different treatments were used for in situ inhibition of L. monocytogenes , including L. monocytogenes alone (positive control) (trial 1), L. monocytogenes + Enterocin BH04 (trial 2), L. monocytogenes + Enterocin BH12 (trial 3), L. monocytogenes + Enterocin BH84 (trial 4), L. monocytogenes + Enterocin BH99 (trial 5), L. monocytogenes + E. faecium BH04 (trial 6), L. monocytogenes + E. faecium BH12 (trial 7), L. monocytogenes + E. faecium BH84 (trial 8), and L. monocytogenes + E. faecium BH99 (trial 9). Fresh cultures of L. monocytogenes and E. faecium were inoculated into reconstituted skim milk (RSM) at approximately 10^5^ cfu/mL, while the bacteriocins were inoculated at 10^3^ AU/mL. The RSM was then incubated at 37°C for 15 h and viable cell counts were determined every 3 h. For the enumeration of L. monocytogenes , PALCAM agar (Merck) with Listeria selective supplement (Merck) was used, and the plates were incubated at 37°C for 24 h (Benkerroum et al. 2002).
Determining Genes Encoding Enterocin Production and Sequencing
2.6
Genes coding for enterocin production were detected by polymerase chain reaction (PCR) using primers specific for entA, entB, entP, and entL50 as presented in table 2 (Du Toit et al. 2000). The genomic DNA of the Enterococcus strains was obtained using the method of Singh and Ramesh (2009). The PCR was performed with 25 μL Taq OptiMix CLEAR Master Mix (AMPLIQON, Odense, Denmark), 1 μL of each primer (forward and reverse), 50 ng/μL template DNA, and nuclease‐free water (Thermo Fisher Scientific, Waltham, Massachusetts, USA). The thermal cycling was as follows: pre‐denaturation at 95°C for 5 min, denaturation at 95°C for 30 s, annealing at the correct temperature (see Table 2) for 40 s, and extension at 72°C for 30 s, for a total of 30 cycles, and post‐extension at 72°C for 5 min. The PCR products were then visualized using ClearBand Safe DNA Gel Stain Solution (Ecotech Biotechnology, Erzurum, Turkey) in 1.5% (w/v) agarose gel under UV light. Genes were sequenced using the ABI 3130 Xl genetic analyzer (Applied Biosystems, Waltham, Massachusetts, USA).
Characterization of Enterocins
2.7
The CFS containing enterocins produced by the Enterococcus spp. was obtained from M17 broth (Merck) containing 0.5% glucose (Merck) for characterization of the enterocins. The CFS was exposed to the following treatments. At the end of all treatments, the percent residual activity (%) of the enterocins was calculated by the two‐fold serial dilution method on BHIA (Sigma‐Aldrich) seeded with L. monocytogenes ATCC 7644.
Temperature Resistance
2.7.1
The CFS was exposed to different temperatures including −80°C, −20°C, 4°C, 37°C, 60°C, 80°C, and 100°C for 10, 30, and 60 min and 121°C for 15 min to determine the cooling and heating resistance of the enterocins (Ogunbanwo et al. 2003).
pH Sensitivity
2.7.2
The pH of the CFS was adjusted to 2, 4, 6, 8, 10, and 12 using 1 N NaOH (Merck) or HCl (Merck) and incubated at room temperature (25°C) for 3 h. The pH of the CFS was then neutralized to eliminate the antimicrobial activity of acidic or basic CFS (Meral‐Aktaş 2023).
Enzyme Treatment
2.7.3
The enterocins‐containing CFS was tested for sensitivity to various enzymes. For this purpose, enzymes including catalase, lysozyme, pepsin, proteinase K, and trypsin (Sigma‐Aldrich) were separately added to the CFS at a concentration of 1 mg/mL. The CFS was then incubated at 37°C for 2 h and heated at 80°C for 10 min to inactivate the enzymes (Todorov and Dicks 2005).
Effect of Chemicals on Enterocin Activity
2.7.4
To determine the effects of some chemicals on enterocin activity, 2‐mercaptoethanol, acetone, chloroform, ethanol, ethylenediaminetetraacetic acid (EDTA), glycerol, methanol, sodium chloride (NaCl), sodium dodecyl sulfate (SDS), triton X‐100, Tween‐20, Tween‐80, urea (Sigma‐Aldrich) and reconstituted skim milk were added to the CFS at a final concentration of 1% (v/v). The CFS was then incubated at 37°C for 5 h (Todorov and Dicks 2005).
Effect of UV Light on Enterocin Activity
2.7.5
The enterocins were exposed to UV light to determine their sensitivity. The CFS was held under UV light (distance: 30 cm) for 5 min (Ogunbanwo et al. 2003).
Statistical Analysis
2.8
The experiments were performed in triplicate, and all results are expressed as mean ± standard deviation (Kenward 1987). All means and standard deviations were calculated using the SPSS version 20.0 package program (SPSS Inc., Chicago, IL, USA).
Results and Discussion
3
Microbial Growth Curve and Bacteriocin Production
3.1
During microbial growth, the pH of the medium or food decreases and the microbial biomass increases. In addition, it is known that some microorganisms can produce antimicrobial peptides such as bacteriocin with their growth (Yang et al. 2018). In this study, microbial growth, including pH and optical density, and enterocin production by the Enterococcus spp. are shown in Figure 1. Considering the pH results, all strains decreased the pH of M17 broth (Merck) from 7.13 to 4.53 within 24 h. Vandera et al. (2020) reported that Enterococcus spp. isolates decreased the pH of M17 broth to 5.51 at the end of the incubation at 30°C for 24 h. On the other hand, Meral‐Aktaş, Erdoğan, and Çeti̇n (2024) found that E. faecium strains reduced the pH of M17 broth containing glucose to 4.50–5.00 (at 37°C for 24 h). Differences in results may be due to pre‐incubation treatment, differences in incubation temperature, and the addition of glucose. In this context, the OD_600_ value increased from 0.068 to 1.034 and the OD_600_ curve clearly showed the microbial growth phases of the strains, including the lag, exponential, and stationary phases. The results concluded that all strains started to produce enterocin with an activity of 0.4 kAU/mL (400 AU/mL) after 3 h. Furthermore, the bacteriocin activity increased to 25.6 kAU/mL (25,600 AU/mL) after 6 h and was maintained throughout the incubation period (Figure S1). Yang et al. (2018) found that the CFS of E. faecium JFR‐1 grown in the De Man, Rogosa, Sharpe (MRS) broth had bacteriocin activity against L. monocytogenes with an activity of 900 AU/mL after 11 h. In addition, Nascimento et al. (2010) found that the bacteriocin activity of E. faecium FAIR‐E 198 grown in MRS broth varied from 1600 to 19,200 AU/mL against different L. monocytogenes strains. These results suggest that the enterocin activity may be affected by differences in the medium, E. faecium strains, and L. monocytogenes strains. On the other hand, there are many studies reporting that low pH may reduce the activity of bacteriocins by microorganisms (Nascimento et al. 2010; Ben Belgacem et al. 2012; Yang et al. 2018). However, in this study, the activity of the enterocins produced by the E. faecium strains was not affected by decreasing pH during the incubation period. Furthermore, the strains rapidly produced enterocins with inhibitory activity against L. monocytogenes ATCC 7644 during the incubation. This study showed that the strains and/or their enterocins may have potential as antilisterial agents due to their rapid production and stability at low pH.
Microbial growth curves and enterocin production abilities of the Enterococcus faecium strains. 1 kAU/mL indicates 1000 AU/mL.
Antibacterial Spectrum of Bacteriocins
3.2
Bacteriocins produced by LAB generally have many applications in food biotechnology due to their heat stability, inhibition effect on saprophytes and/or pathogens in products, and suppression of uncontrolled growth of starter cultures (Cotter et al. 2005). In addition, bacteriocins are known to have a broad inhibitory effect on Gram‐positive bacteria (De Kwaadsteniet et al. 2005). Although not as broad as the effect on Gram‐positive bacteria, some bacteriocins have also been reported to have an effect on Gram‐negative bacteria. For example, De Kwaadsteniet et al. (2005) reported that bacteriocins from Enterococcus mundtii ST15 inhibited Gram‐negative bacteria such as Acinetobacter baumannii, Klebsiella pneumoniae , and Pseudomonas aeruginosa , while Woraprayote et al. (2015) reported that bacteriocins from Weissella hellenica BCC 7293 inhibited Aeromonas hydrophila , Escherichia coli , Pseudomonas aeruginosa , and Salmonella Typhimurium . In addition, Wang et al. (2022) showed that the combination of bacteriocins by LAB with chelators such as lactic acid could increase the inhibitory effect on Gram‐negative bacteria. Therefore, the antibacterial activity of the bacteriocins should be investigated in a broad spectrum. In this study, the antibacterial spectrum of the enterocins obtained from E. faecium strains against some pathogens and LAB was determined, and the results are given in Table 1. All the strains had an inhibitory effect on L. monocytogenes ATCC 7644 ranging from 19.3 to 20.5 mm (Figure S2) as reported by García et al. (2004), Meral‐Aktaş (2023) and Azevedo et al. (2024). In addition, the strains also inhibited the growth of Heyndrickxia coagulans ATCC 7050 (previously named Bacillus coagulans, Narsing Rao et al. (2023)) between 10.0 and 12.5 mm. Similar results were reported by García et al. (2003) and Lucas et al. (2006). The results showed that E. faecium BH84 and BH99 had an inhibitory effect on Lactobacillus delbrueckii subsp. bulgaricus 239 M (21.0 mm), while E. faecium BH08 and BH12 did not. This was an expected result, as Lactobacillus delbrueckii subsp. bulgaricus is known to be susceptible to bacteriocins (Hegarty et al. 2017). On the other hand, other Lactobacillus spp. used for antibacterial spectrum, including Lactobacillus helveticus ATCC 15009 and Lactobacillus casei ATCC 393, were not affected by enterocins. Therefore, it can be concluded that the enterocins may be important for both food safety and food quality due to their inhibitory effect on L. monocytogenes ATCC 7644.
In Vitro (Mode of Action) and In Situ (Milk Model) Inhibition of
Listeria monocytogenes
3.3
In this study, the in vitro inhibition effect of enterocin BH04, BH12, BH84, and BH99 on L. monocytogenes ATCC 7644 was investigated, and the results obtained are shown in Figure 2 and Table S2. The OD_600_ value of BHI with L. monocytogenes ATCC 7644 alone varied from 0.07 to 0.54 over the incubation time. On the other hand, the OD_600_ value of BHI with L. monocytogenes ATCC 7644 and enterocin BH04, BH12, BH84, and BH99 increased up to 0.16 at the end of 15 h. It was concluded that all enterocins used in this study were able to inhibit the growth of L. monocytogenes ATCC 7644 in BHI compared to the control culture sample containing the pathogen growing alone. Similarly, there are many studies reporting that the enterocins obtained from E. faecium strains can exhibit a bacteriostatic mode of action on L. monocytogenes (Chakchouk‐Mtibaa et al. 2014; Yang and Moon 2021; Du et al. 2022).
The growth of L. monocytogenes ATCC 7644 in BHI broth containing enterocins. ○: L. monocytogenes ATCC 7644 alone, □: L. monocytogenes ATCC 7644 + enterocin BH04, ∆: L. monocytogenes ATCC 7644 + enterocin BH12, ▽: L. monocytogenes ATCC 7644 + enterocin BH84, ◊: L. monocytogenes ATCC 7644 + enterocin BH99.
The results of the in situ inhibitory activity of enterocin BH04, BH12, BH84, and BH99 on L. monocytogenes ATCC 7644 in the RSM are presented in Figure 3 and Table S3. The count of L. monocytogenes ATCC 7644 alone increased from 5.33 to 7.77 log cfu/mL at the end of 15 h. However, the viable cell count of the pathogen in the RSM combined with E. faecium BH04, BH12, BH84, and BH99 was between 5.59 and 5.96 log cfu/mL at the end of the incubation time. The E. faecium strains were bacteriostatic against L. monocytogenes ATCC 7644 in the RSM model. In addition, the count of L. monocytogenes ATCC 7644 in RSM containing enterocins BH04, BH12, BH84, and BH99 fell below the countable limit by the twelfth hour of incubation. It was concluded that all enterocins inhibited L. monocytogenes ATCC 7644 and had bactericidal activity against the pathogen. Similarly, Nascimento et al. (2010) reported that E. faecium FAIR‐E 198 strain had bactericidal activity against L. monocytogenes Scott A in the skimmed milk. Furthermore, in the study by Vandera et al. (2017), in skimmed milk (pH decreased from 6.42 to 6.04 during incubation) containing enterocin‐producing E. faecium KE82, it was found that the count of L. monocytogenes was suppressed (4.23 log cfu/mL) after 6 h of incubation compared to the control (5.21 log cfu/mL), and this difference was statistically significant.
In situ inhibition of L. monocytogenes ATCC 7644 in RSM. Cfu: Colony‐forming unit, h: Hour. Trial 1: L. monocytogenes alone (positive control), trial 2: L. monocytogenes + Enterocin BH04, trial 3: L. monocytogenes + Enterocin BH12, trial 4: L. monocytogenes + Enterocin BH84, trial 5: L. monocytogenes + Enterocin BH99, trial 6: L. monocytogenes + E. faecium BH04, trial 7: L. monocytogenes + E. faecium BH12, trial 8: L. monocytogenes + E. faecium BH84, trial 9: L. monocytogenes + E. faecium BH99.
The results of the in vitro and in situ inhibition experiments indicated that enterocin BH04, BH12, BH84, and BH99 could be used as antilisterial agents. Moreover, * E. faecium* BH04, BH12, BH84, and BH99 strains were also able to suppress the growth of L. monocytogenes ATCC 7644. Therefore, it is thought that these strains can be used as a co‐culture to control the pathogens.
Determining Genes Encoding Enterocin Production and Sequencing
3.4
In this study, entA, entB, entP, and entL50 genes encoding enterocin production were screened by PCR. The resulting gel images are shown in Figure 4. The results showed that all the strains used in this study had entA, entB, and entP genes as in the study by Meral‐Aktaş, Erdoğan, and Çeti̇n (2024). On the contrary, the entL50 gene was not detected in any of the strains. In addition, the nucleotide sequencing of entA, entB, and entP genes of the strains is shown in Figure 5. entA and entB genes in all strains had 100% identity with each other and with entA (OR566524.1 and OR566518.1) and entB (MF457399.1) genes in the National Center for Biotechnology Information (NCBI). On the other hand, entP genes in all strains had up to four nucleotide gaps. entP genes were 98.68% identity with FJ416486.1 and 100% identity with DQ867125.1 when these gaps were ignored.
The gel image of genes encoding enterocin production, including entA, entB, entP, and entL50, in Enterococcus faecium BH04, BH12, BH84, and BH99 strains. M: DNA ladder, bp: Base pair, PC: Positive control, NC: Negative control.
Nucleotide sequencing of entA (A), entB (B), entP (C) genes of the strains.
Characterization of Enterocins
3.5
In this study, enterocin BH04, BH12, BH84, and BH99 were characterized with respect to different treatments including temperature, pH, enzyme, chemical, and UV. The characterization results are shown in Table 3. Cold storage (−80°C, −20°C, and +4°C) and hot storage (37°C, 60°C, and 80°C) did not affect the antilisterial activity of enterocins. On the other hand, the residual activity decreased by 50% after 30 and 60 min at 100°C, whereas it did not change after 10 min at 100°C. In addition, the inhibitory effect of all enterocins on L. monocytogenes ATCC 7644 was significantly reduced after sterilization (at 121°C for 15 min). Bacteriocins are known to have a protein structure and to be heat‐stable (Gálvez et al. 2007). The present study revealed that the enterocins were stable against cold storage and heating up to 100°C for 10 min. Similar results were also reported by Mareková et al. (2007) and Meral‐Aktaş (2023).
The antilisterial activity of enterocin BH04, BH12, BH84, and BH99 was not affected by pH 2, 4, 6, 8, and 10 treatments as reported by Qiao et al. (2020). However, in the study by Du et al. (2022), the residual activity of enterocin HDX‐2 obtained from E. faecium HDX‐2 at pH 10 was found to be reduced to 80.28%. Considering these results, it can be reported that the stability of enterocins BH04, BH12, BH84, and BH99 is higher in alkaline medium. However, the enterocins used in this study lost about 90% of their antilisterial activity at pH 12. Similarly, Yang and Moon (2021) reported that the activity of enterocin from E. faecium CJNU 2524 decreased by 50% in pH 11. All these results revealed that the enterocins may lose their antilisterial activity in environments above pH 10.
The inhibitory effect of the enterocins used in this study on L. monocytogenes ATCC 7644 was not altered by the addition of catalase and lysozyme. Similarly, Franz et al. (1996) found that enterocin 900 from E. faecium BFE 900 was stable to catalase and lysozyme. On the other hand, enterocins are known to be sensitive to some proteolytic enzymes such as pepsin, proteinase K, and trypsin (Franz et al. 1996; Kumar et al. 2010; Qiao et al. 2020). The results of this study showed that the enterocins were sensitive to pepsin, proteinase K, and trypsin (except the enterocin BH04 against pepsin).
All enterocins were stable to all chemicals used in this study. In addition, ethanol, methanol, and SDS increased the antilisterial activity of the enterocins by between 200% and 400%. These results were expected because ethanol (Oh and Marshall 1993) and methanol (Sidorenko and Buzoleva 2012) have antimicrobial activity against L. monocytogenes . In addition, SDS is known to have a potentiating or antimicrobial effect (Kaktcham et al. 2019). Therefore, similar results were found in the present study.
In this study, UV treatment reduced antilisterial activity of enterocins by 50% for BH04, BH84, and BH99 and by 75% for BH12. Ogunbanwo et al. (2003) reported that bacteriocins produced by Lactobacillus plantarum F1 and Lactobacillus brevis OG1 were not affected by UV light. It is suggested that these different results may be due to the type of bacteriocin.
Conclusion
4
This study demonstrated that E. faecium BH04, BH12, BH84, and BH99 have the potential to produce enterocins effective against L. monocytogenes ATCC 7644, an important foodborne pathogen. The bactericidal and bacteriostatic activity of enterocin BH04, BH12, BH84, and BH99 against L. monocytogenes ATCC 7644 by strains carrying entA, entB, and entP genes encoding enterocin production was demonstrated using in vitro mode of action and in situ milk model. In addition, the enterocins retained their antilisterial activity under certain conditions such as heating and acidic environment. Finally, because of all this and because the enterocins were effective against L. monocytogenes ATCC 7644 but had no inhibitory effect on LAB such as Streptococcus thermophilus 212S, Lactobacillus helveticus ATCC 15009, and Lactobacillus casei ATCC 393, it was concluded that these strains or their bacteriocins could be used as antilisterial agents in fermented products.
Conflicts of Interest
The author declares no conflicts of interest.
Supporting information
Data S1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aktaş, H. 2023. “Isolation of Probiotic and Starter Yoghurt Bacteria and Determination of Microbiological, Physicochemical and Technological Properties of Yoghurts Produced With Selected Strains.” Ph D thesis, Atatürk University.
- 2Aktaş, H. , and B. Çetin . 2024. “Multidimensional Evaluation of Techno‐Functional Properties of Yoghurt Bacteria.” International Dairy Journal 148: 105795. 10.1016/j.idairyj.2023.105795. · doi ↗
- 3Azevedo, I. , J. Barbosa , H. Albano , T. Nogueira , and P. Teixeira . 2024. “Lactic Acid Bacteria Isolated From Traditional and Innovative Alheiras as Potential Biocontrol Agents.” Food Microbiology 119: 104450. 10.1016/j.fm.2023.104450.38225051 · doi ↗ · pubmed ↗
- 4Balouiri, M. , M. Sadiki , and S. K. Ibnsouda . 2016. “Methods for In Vitro Evaluating Antimicrobial Activity: A Review.” Journal of Pharmaceutical Analysis 6: 71–79. 10.1016/j.jpha.2015.11.005.29403965 PMC 5762448 · doi ↗ · pubmed ↗
- 5Ben Belgacem, Z. , A. Rehaiem , P. Fajardo Bernárdez , M. Manai , and L. Pastrana Castro . 2012. “Interactive Effects of p H and Temperature on the Bacteriocin Stability by Response Surface Analysis.” Microbiology 81: 195–200. 10.1134/S 002626171201002 X.22693830 · doi ↗ · pubmed ↗
- 6Benkerroum, N. , Y. Ghouati , H. Ghalfi , et al. 2002. “Biocontrol of Listeria monocytogenes in a Model Cultured Milk (Lben) by In Situ Bacteriocin Production From Lactococcus lactis Ssp. Lactis.” International Journal of Dairy Technology 55, no. 3: 145–151. 10.1046/j.1471-0307.2002.00053.x. · doi ↗
- 7Çetin, B. , and H. Aktaş . 2024. “Monitoring Probiotic Properties and Safety Evaluation of Antilisterial Enterococcus faecium Strains With Cholesterol‐Lowering Potential From Raw Cow's Milk.” Food Bioscience 61: 104532. 10.1016/j.fbio.2024.104532. · doi ↗
- 8Chakchouk‐Mtibaa, A. , L. Elleuch , S. Smaoui , et al. 2014. “An Antilisterial Bacteriocin Bac FL 31 Produced by Enterococcus faecium FL 31 With a Novel Structure Containing Hydroxyproline Residues.” Anaerobe 27: 1–6. 10.1016/j.anaerobe.2014.02.002.24583094 · doi ↗ · pubmed ↗