Analysis of a cancer-associated mutation in the budding yeast Nuf2 kinetochore protein
Angelica Andrade Latino, Sue Biggins

TL;DR
This study examines a cancer-related mutation in the NUF2 protein using yeast to understand its impact on cell division and cancer.
Contribution
The study uses a yeast model to investigate a human cancer-associated mutation in the NUF2 protein.
Findings
The nuf2 Q21A mutation in yeast does not cause growth defects or disrupt kinetochore composition.
The mutation does not affect interactions with the Dam1 complex or Mps1 kinase.
The results suggest the human HsNUF2 R19H mutation's role in cancer needs further investigation in human cells.
Abstract
The kinetochore is a highly conserved megadalton protein complex that ensures proper chromosome segregation via microtubule attachments. The NDC80 complex is one of the major conserved microtubule binding complexes in the kinetochore. NUF2, a protein within the NDC80 complex, has been identified as a cancer gene candidate because missense mutations, found across different tumor samples, cluster within NUF2’s calponin homology domain. In this study, we examined a NUF2 cancer-associated mutation in a simple and well-studied organism, Saccharomyces cerevisiae , to elucidate its effects on cell division. We studied the budding yeast nuf2 Q21A mutation with the intention of extrapolating our results to the homologous cancer associated mutation in Homo sapiens NUF2 R19H (HsNUF2 R19H ). Our studies demonstrate that the nuf2 Q21A mutant does not exhibit any growth defects or disrupt…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
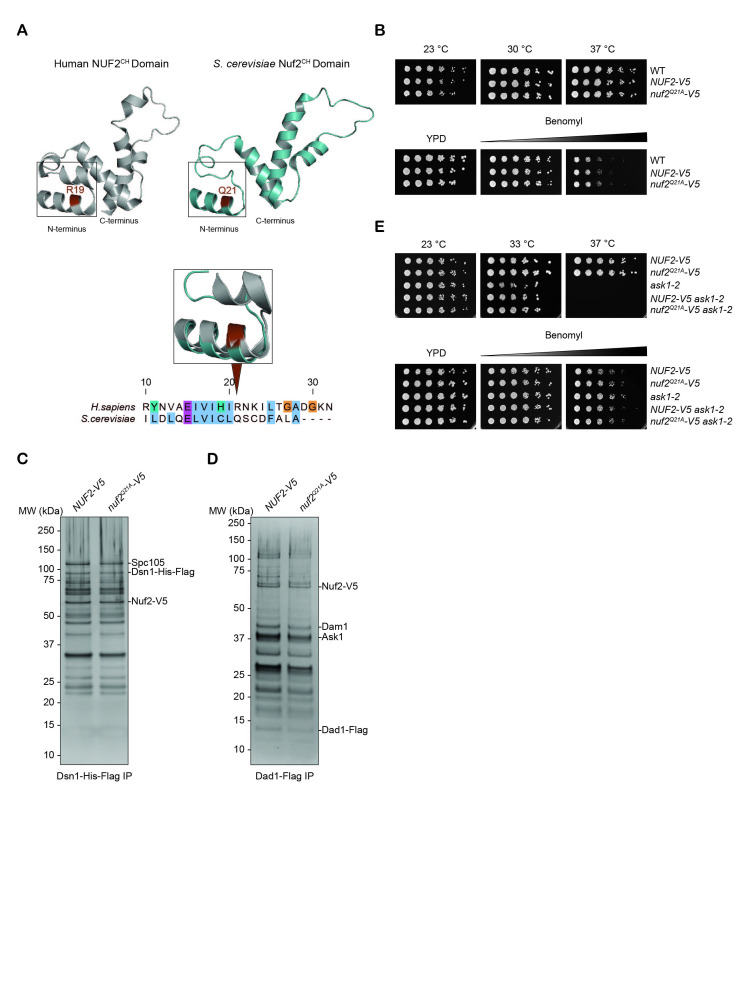 Figure 1
Figure 1|
Strains |
Genotype |
Reference |
|
SBY3 (W303) |
Mat
|
Biggins Lab |
|
SBY4 (W303) |
Matα
|
Biggins Lab |
|
|
Matα
|
De Regt et al., 2022 |
|
SBY1300 |
Mat
|
Gift from Elledge’s Laboratory, Harvard University, Cambridge, MA |
|
|
Matα
|
This study |
|
|
Matα
|
This study |
|
|
Mat
|
This study |
|
|
Matα
|
This study |
|
|
Mat
|
This study |
|
|
Matα
|
This study |
|
|
Mat
|
This study |
|
|
Mat
|
This study |
|
|
Mat
|
This study |
|
|
Mat
|
This study |
|
Plasmid number |
Purpose |
Reference |
|
pSB3158 |
Integration of the
|
Gift from Elçin Ünal’s Laboratory, University of California, Berkeley, CA Janke et al., 2004 |
|
pSB3218 |
Generate sgRNA used for CRISPR/Cas9 mutagenesis. |
Gift from Elçin Ünal’s Laboratory, University of California, Berkeley, CA |
|
pSB3608 |
Generated sgRNA (pSB3218 derivative) used for Nuf2-Q21 CRISPR/Cas9 mutagenesis. |
This study |
|
Oligo number |
Purpose |
Sequence |
Reference |
|
SB8342 |
Forward oligo for Gibson assembly to generate sgRNA for
|
5’- ggctgggcaacaccttcgggtggcgaatggGTTCCCCATTTTGGATCTAC -3’ |
This study |
|
SB8343 |
Reverse oligo for Gibson assembly to generate sgRNA for
|
5’- attttaacttgctatttctagctctaaaacGTAGATCCAAAATGGGGAAC -3’ |
This study |
|
SB8350 |
Forward primer to amplify
|
5’- TATTAATAAATACATGAATGAAATGCTCGAATATATGCAAgcggccgctctagaactagt -3’ |
This study |
|
SB8351 |
Reverse primer to amplify
|
5’- AGAAAACACAGAAGGGGGAGTAAAAATAAGTATACCGCTGccccctcgaggtcgacggta -3’ |
This study |
|
SB8371 |
Forward primer to amplify entire
|
5’- CTCCAGCATCCCTTGAGCAA -3’ |
This study |
|
SB8372 |
Reverse primer to amplify entire
|
5’- ACCTTCACATGTTCCGCAGA -3’ |
This study |
|
SB8373 |
Repair template for
|
5’- GAAAAAATACTGGTAAAAAGCATGTACTGAGGAGAAAGGCTCCAGCATCCCTTGAGCAAAATGAGTAGGAATCAAGATGTGTTCCCCATTTTGGATCTACAAGAACTAGTTATATGTTTGCATAGCTGTGATTTTGCGCTAGCCACACAGGAAAATATCTCTAGGCCCACCTCAGACTACATGGTAACCCTTTACAAACA -3’ |
This study |
|
SB8466 |
Amplify 5' end of
|
5’- TGCGGTTAAGTCGTCTAACG -3’ |
This study |
|
Name |
Type |
Species |
Brand |
Reference |
|
M3DK (α-Flag) |
Monoclonal |
Mouse |
GenScript |
(Barrero et al., 2024) |
- —National Institutes of Health (United States)https://ror.org/01cwqze88
- —National Institutes of Health (United States)https://ror.org/01cwqze88
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrotubule and mitosis dynamics · Ubiquitin and proteasome pathways · Endoplasmic Reticulum Stress and Disease
Description
All eukaryotic cells undergo mitosis, a key event required to produce two genetically identical daughter cells during cell division (Santaguida and Musacchio 2009). Defects in this crucial process can lead to chromosome mis-segregation events that result in aneuploid cells, a hallmark of cancer (Holland and Cleveland 2009). Proper chromosome segregation during mitosis requires the kinetochore, a highly conserved and tightly regulated protein network that assembles at centromeres and forms microtubule attachments (Santaguida and Musacchio 2009; Musacchio and Desai 2017; Ariyoshi and Fukagawa 2023). During anaphase, microtubules pull chromosomes to opposite poles of a dividing cell via kinetochore attachments to ensure proper inheritance of the genetic material.
The two major budding yeast kinetochore subcomplexes responsible for forming microtubule attachments are the Ndc80 and Dam1 complexes (Ndc80c and Dam1c; Biggins 2013). The role of the Ndc80c is to form stable, load-bearing attachments to dynamic microtubules. The Dam1c facilitates these attachments by interacting with the Ndc80c and oligomerizing around microtubules enabling the Ndc80c to track the tip of dynamic microtubules with higher affinity (Westermann et al., 2005; Westermann et al., 2006; Lampert et al., 2010; Tien et al., 2010; Lampert et al., 2013). The highly conserved Ndc80c is a heterotetramer comprised of Ndc80p-Nuf2 and Spc24-Spc25 heterodimers (Wei et al., 2005). Each heterodimer is positioned on opposite ends of the dumbbell shaped complex with Ndc80p-Nuf2 oriented to interact with the microtubule while Spc24-Spc25 connects the complex to the rest of the kinetochore (Wei et al., 2005; Cheeseman et al., 2006). The N-terminus of each protein within the Ndc80p-Nuf2 heterodimer folds into independent globular calponin homology (CH) domains (Wei et al., 2007; Ciferri et al., 2008). The CH domain of Ndc80p interacts with microtubules directly, whereas the CH domain of Nuf2 does not (Alushin et al., 2010; Sundin et al., 2011). However, the Nuf2 ^CH^ domain plays a critical role in chromosome segregation. In S. cerevisiae , the Nuf2 ^CH ^ domain contains a conserved patch that is a docking site for the Dam1c and the kinase Mps1 that regulates kinetochore-microtubule attachments (Maure et al., 2007; Sarangapani et al., 2021; Parnell et al., 2024; Pleuger et al., 2024; Zahm et al., 2024). In HeLa cells, charge reversal mutations within the NUF2 ^CH^ domain resulted in cells arresting in mitosis (Sundin et al., 2011).
A previous study mapped somatic missense mutations, identified from different cancerous tumor samples, onto the available PDB (2VE7) structure of NUF2 and found that they significantly cluster within its CH domain (Kamburov et al., 2015). We speculated that the NUF2 cancer associated missense mutations could affect the structure of the CH domain and alter its protein function within cancerous cells. The aim of this study was to elucidate the effects that the human cancer associated missense mutations in NUF2 have on kinetochore composition and function by examining them in S. cerevisiae . Since the mutants are viable in cancer cells, we used budding yeast as a tractable system because it provides assays that allow for more in-depth analyses of these mutations than in human cells. For example, we can perform sensitive growth assays as well as study genetic and biochemical interactions between kinetochore proteins to uncover the functional consequences of the cancer-associated mutations.
The CH domain of NUF2 is conserved across *H. sapiens * and S. cerevisiae (Ciferri et al., 2008), so we performed a structural alignment of both proteins to identify the location of the cancer-associated residues in the yeast protein. Utilizing PyMOL, we aligned the NUF2 protein structures from the H. sapiens NDC80c ^Bonsai ^ and S. cerevisiae Ndc80c ^Dwarf^ PDBs ( Fig. 1A, top). Out of the 8 cancer-associated residues identified (Kamburov et al., 2015), 6 are structurally conserved in S. cerevisiae . Out of the 6 structurally conserved residues, we focused on the HsNUF2-R19 amino acid since it has the highest mutation frequency ( cBioPortal ) and is equivalent to Nuf2-Q21 in *S. cerevisiae * as depicted by the overlay of the two protein structures ( Fig. 1A, inset). A recent structural study revealed the Nuf2-Q21 residue to be nestled within an interacting pocket between the Ndc80c ^CH ^ domains and the Dam1c (Muir et al., 2023; Zahm et al., 2023). Residues of the Dam1c that are involved in this interaction are phosphoregulated by Aurora B/Ipl1 kinase when erroneous kinetochore-microtubule attachments are formed (Keating et al., 2009). The Nuf2-Q21 residue also lies near a highly conserved “interaction hub” in the Nuf2 ^CH ^ domain that binds to Mps1 and the Dam1c (Parnell et al., 2024; Pleuger et al., 2024; Zahm et al., 2024). Therefore, a mutation at the Nuf2-Q21 amino acid could alter these vital interactions.
The Nuf2-Q21 residue is positioned on the outer surface of an alpha helix, most likely engaging in intermolecular interactions with other nearby proteins. We therefore generated a mutant that would most effectively disrupt interactions by substituting the glutamine with an alanine to introduce a cavity. The * nuf2 ^Q21A^ * mutant was made in a *NUF2-V5 * background strain at the endogenous locus via CRISPR/Cas9 mutagenesis.
To test for growth defects, we performed a serial dilution growth assay. Strains were plated at various temperatures or onto various concentrations of benomyl, a microtubule depolymerizing agent ( Fig. 1B ). The assays demonstrated that the presence of the V5 epitope tag does not affect cell viability and that the * nuf2 ^Q21A^ * mutant did not exhibit growth defects under any of the tested conditions.
We next examined kinetochore composition by purifying the Dsn1 protein because it isolates most of the budding yeast native kinetochore (Akiyoshi et al., 2010; Gupta et al., 2018). We purified Dsn1 from WT and * nuf2 ^Q21A^ * mutant cells and analyzed kinetochore composition through silver-stained SDS-PAGE analysis. The * nuf2 ^Q21A^ * mutant had no detectable changes in kinetochore composition compared to WT ( Fig. 1C ). Some kinetochore proteins that might be affected by the presence of the * nuf2 ^Q21A^ * , such as the Mps1 kinase, are not easily detected via silver-stained analysis. As such, we performed mass spectrometry (MS) on the Dsn1-His-Flag immunoprecipitation samples. MS analysis revealed Mps1 to be among the most enriched proteins detected; however, there were no significant changes in its abundance between WT and the * nuf2 ^Q21A^ * mutant (Extended data files 1 and 2). Additionally, the MS analysis further corroborated the findings of the silver-stained analysis in that there were no significant changes in peptide counts detected across the co-precipitated kinetochore proteins (Extended data files 1 and 2). Together, these findings indicate the * nuf2 ^Q21A^ * mutant does not perturb cell viability, or the Ndc80c’s interaction with Mps1 kinase and other kinetochore proteins isolated in the Dsn1-His-Flag purification.
Next, we tested whether the * nuf2 ^Q21A^ * mutant affected physical or genetic interactions between the Ndc80c and the Dam1c. Since the Dam1c is substoichiometric in the Dsn1-His-Flag purifications, we directly purified the Dam1c component Dad1 via a Dad1-Flag co-immunoprecipitation experiment to assay its interaction with Nuf2 ( Fig. 1D ). In both WT and the mutant, similar amounts of Nuf2 co-purified with Dad1, indicating that the * nuf2 ^Q21A^ * mutant does not disrupt interactions between the Ndc80c and the Dam1c. We also asked whether the * nuf2 ^Q21A^ * mutant exhibited genetic interactions with a mutant in the Dam1c. We introduced the * nuf2 ^Q21A ^ * mutant into a background strain that cripples the Dam1c using the temperature sensitive allele *ask1-2 * (Li et al., 2002). The double mutant was viable even when its microtubule attachments were challenged and did not exhibit temperature sensitivity at the ask1-2 semi-permissive temperature of 33 °C ( Fig. 1E ). Taken together, these data suggest that the * nuf2 ^Q21A^ * mutant does not have a major effect on Ndc80c function or affect the Ndc80c’s interaction with the Dam1c.
In sum, we demonstrated that the * nuf2 ^Q21A^ * mutant does not have detectable effects on kinetochore composition or exhibit any growth defects, suggesting it does not influence kinetochore-microtubule attachments. It is therefore unclear whether the homologous HsNUF2 ^R19H^ missense mutation contributes a significant role in the tumorigenesis of the cancers it was found in. It is important to note that the HsNUF2 ^R19H ^ is not a hotspot mutation and was obtained from an outdated dataset. With the emergence of new sequencing profiles of more tumor samples, new mutations should be considered for future studies. For example, the HsNUF2 ^S340L^ now has a higher mutation frequency than the HsNUF2 ^R19H^ ( cBioPortal ). Ultimately, the HsNUF2 ^R19H^ mutant should be studied in human cells as the region this residue occupies is not highly conserved at the primary sequence ( Fig. 1A ). Consistent with this limitation, we were unable to generate a chemically similar HsNUF2 ^R19H ^ mutation in yeast since the chemical properties of the arginine and glutamine side chains are distinctly different from one another. Additionally, the Dam1c complex is not conserved but has a functional ortholog called the SKA complex in metazoan cells (Hanisch et al., 2006; Welburn et al., 2009). Cross-linking mass spectrometry demonstrates a potential interaction between the CH domains of the mammalian NDC80c with the C-terminus of SKA3, a protein within the SKA complex (Helgeson et al., 2018). It will therefore be interesting to explore whether HsNUF2 ^R19H^ mutation affects the NUF2-SKA3 interaction in human cells. Similarly, although the interaction between MPS1 and NUF2 ^CH^ domain is conserved in metazoan cells, the region at which the interaction occurs differs. In human cells, the NUF2 ^CH ^ domain contacts the middle region of the MPS1 kinase (Ji et al., 2015) whereas in S. cerevisiae the interaction occurs at the N-terminal tail of Mps1 (Parnell et al., 2024; Pleuger et al., 2024; Zahm et al., 2024). Hence the different modes of interaction may account for the lack of phenotype observed in the * nuf2 ^Q21A^ * mutant. In conclusion, future studies in human cells may shed light on the function of HsNUF2-R19 residue and its potential role in cancer.
Methods
Culture conditions
All S. cerevisiae strains used in this study were grown in yeast extract peptone dextrose (YPD) liquid cultures, containing 2% D-glucose (Sigma-Aldrich) and 0.02% adenine (MP Biomedicals). Cultures were grown at 23-30 ^°^ C and were harvested at late-log phase OD 600nm = ~3.
Spotting dilution assay
Strains were grown overnight in 5 ml of YPD liquid. Utilizing the SmartSpec Plus spectrometer (Bio-Rad), the concentration of cells was measured at OD 600nm and were diluted to OD 600nm = 1. Each strain underwent a five-fold serial dilution in a 96-well plate. The first column of the plate contained fully saturated OD 600nm = 1 cells and subsequent wells had a 1:5 dilution in water. Strains were spotted onto YPD and YPD benomyl coated plates (5 μg/ml and 15 μg/ml). YPD plates containing benomyl were incubated at 23 ^°^ C while the plates containing just YPD were incubated at the indicated temperatures. Cells were grown for 3-5 days.
One-step purifications & silver-staining
Flag purifications were performed as previously described (Gupta et al., 2018) except lysates were treated with benzonase as described below. Briefly, cells were harvested during late-log phase (OD 600nm = ~3) and then flash frozen in Buffer H/0.15 (25 mM HEPES pH 8.0, 2 mM MgCl 2 , 0.1 mM EDTA pH 8.0, 0.5 mM EGTA-KOH pH 8.0, 15% Glycerol, 0.1% NP-40, and 150 mM KCl) containing phosphatase inhibitors (1 mM sodium-pyrophosphate, 2 mM sodium-β-glycerophosphate, 0.1 mM sodium orthovanadate, and 5 mM NaF), 0.1 mM microcystin-LR, 0.2 mM PMSF, and protease inhibitors (10 μg/mL for each of the following leupeptin, pepstatin A, and chymostatin). Pellets were lysed via a Freezer Mill (SPEX SamplePrep) and were then treated with 50 units/ml of benzonase (EMD Milipore Corp) for 30 minutes on ice. Treated lysates were then ultracentrifuged at 24,000 RPM for 90 minutes at 4 ^°^ C and protein layer was extracted. The Pierce BCA Protein Assay Kit (Thermo Scientific) was used to measure the protein concentration within each extract which were subsequently normalized for each immunoprecipitation experiment. Conjugated Flag Protein G Dynabeads were made via crosslinking the α-M3DK antibody (Barrero et al., 2024) [GenScript] to Protein G Dynabeads (Invitrogen). These beads were incubated with the normalized protein extracts for 3 hours at 4 °C. After the incubation, beads were washed three times with Buffer H/0.15 containing phosphatase inhibitors, protease inhibitors, 0.1 mM microcystin-LR, 2 mM DTT and then two times with Buffer H/0.15 containing protease inhibitors, and LPC. Proteins were eluted off beads into Buffer H/0.15 containing 0.2 mM PMSF, protease inhibitor stock and 0.5mg/mL 3 x M3DK Peptide (Sigma-Aldrich) via gentle agitation for 30 minutes at room temperature.
5-10 μl of the immunoprecipitated samples were boiled in 1x sample buffer (Invitrogen 1x LDS sample buffer, 5% β-mercaptoethanol, and sterile water) for 5 mins at 95 °C. Each immunoprecipitated sample was run on precast 4-12% Bis-Tris gels (Thermo Fisher Scientific). Gels were stained with silver-staining agents following the Silver Quest Staining Kit (Invitrogen) instructions. The ChemiDoc MP system (Bio-Rad) was used for imaging.
Protein sequence alignment
Nuf2 protein sequences were obtained from UniProt ( *Homo sapiens, * Q9BZD4) and the *Saccharomyces * Genome Database (W303 background). Entire protein sequence alignment was generated utilizing the default conditions of the Clustal Omega algorithm in Jalview (version 2.11.4.1). Clustal color scheme was used to denote the conservation and the biological properties of the side chains. Image was exported as an EPS file.
Protein structure alignment
Utilizing PyMOL (version 3.1.3), Nuf2 ^CH^ domains from the following PDB structures were isolated: PDB: 2VE7 (NDC80c ^Bonsai^ ) and PDB: 5TCS (Ndc80c ^Dwarf^ ). Alignment was performed of only amino acids 10 or 12 to 100 of each Nuf2 ^CH^ domain. Conserved residues were identified through the alignment. Protein structures were then individually exported as PNGs in the aligned orientation. A similar procedure was followed for the inset, except only residues 10-30 of HsNUF2 and residues 12-32 of Saccharomyces cerevisiae Nuf2 were aligned.
Mass spectrometry preparation and analysis
Immunoprecipitated samples were prepared as indicated under the one-step purification method until the last wash step. After the last wash, samples were rinsed two times with pre-elution rinse buffer (50 mM Tris pH 8.3, 75 mM KCl, and 1 mM EGTA). The samples were eluted via gentle agitation on a Vortex-Genie 2 (Scientific Industries) set to the lowest setting for 30 minutes at room temperature. Each sample was eluted in 70 μl of 0.2% RapiGest (Waters Corporation) in 50 mM ammonium bicarbonate. 10 μl of each elution sample was used for silver-stained analysis as indicated under silver-staining methods while the remaining 60 μl was sent for mass spectrometry processing.
Samples were prepared for LC-MS/MS analysis by reducing disulfide bonds, capping free cysteines, and digesting the proteins with trypsin. The resulting peptide samples were analyzed by LC-MS/MS on an Thermo Eclipse mass spectrometer. The mass spectrometry data collected in this study is available through Mass Spectrometry Interactive Virtual Environment (MassIVE, University of California San Diego; ftp://massive.ucsd.edu/v09/MSV000097121/ ).
Reagents
Yeast strains and plasmid methods
To generate the NUF2-3V5:HIS3Mx6 strain, a *3V5:HIS3Mx6 * cassette was amplified from pSB3158 using PCR. This cassette was then inserted into the endogenous locus of *NUF2 * at the 3’ of the coding region replacing the stop codon via homologous recombination (Petracek and Longtine 2002). To generate the * nuf2 ^Q21A^ * missense mutation in this strain, a single-step CRISPR/Cas9 edit was performed (Akhmetov et al., 2018). *DAD1-3Flag * was made as described (De Regt et al., 2022). DSN1-6His-Flag was generated as described (Akiyoshi et al., 2010). The *ask1-2 * yeast strain was a gift from Stephen Elledge’s laboratory (Harvard University, Cambridge, MA) and plasmids pSB3158 and pSB3218 were gifts from Elçin Ünal’s Laboratory (University of California, Berkeley, CA).
Saccharomyces cerevisiae strains used throughout this study are derivatives of SBY3 and SBY4 (W303).
**: **
Plasmids used in this study.
**: **
Oligonucleotides used in this study.
**: **
Antibody used in this study:
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Akhmetov A Laurent JM Gollihar J Gardner EC Garge RK Ellington AD Kachroo AH Marcotte EM 2018320 Single-step Precision Genome Editing in Yeast Using CRISPR-Cas 9.Bio Protoc 862331-832510.21769/Bio Protoc.276529770349 PMC 5951413 · doi ↗ · pubmed ↗
- 2Akiyoshi B Sarangapani KK Powers AF Nelson CR Reichow SL Arellano-Santoyo H Gonen T Ranish JA Asbury CL Biggins S 20101125 Tension directly stabilizes reconstituted kinetochore-microtubule attachments.Nature 46873230028-083657657910.1038/nature 0959421107429 PMC 3108429 · doi ↗ · pubmed ↗
- 3Alushin Gregory M. Ramey Vincent H. Pasqualato Sebastiano Ball David A. Grigorieff Nikolaus Musacchio Andrea Nogales Eva 2010101 The Ndc 80 kinetochore complex forms oligomeric arrays along microtubules Nature 46773170028-083680581010.1038/nature 0942320944740 PMC 2957311 · doi ↗ · pubmed ↗
- 4Barrero DJ Wijeratne SS Zhao X Cunningham GF Yan R Nelson CR Arimura Y Funabiki H Asbury CL Yu Z Subramanian R Biggins S 202489 Architecture of native kinetochores revealed by structural studies utilizing a thermophilic yeast.Curr Biol 34170960-982238813893.e 510.1016/j.cub.2024.07.03639127048 PMC 11387133 · doi ↗ · pubmed ↗
- 5Biggins Sue 201381 The Composition, Functions, and Regulation of the Budding Yeast Kinetochore Genetics 19441943-263181784610.1534/genetics.112.14527623908374 PMC 3730914 · doi ↗ · pubmed ↗
- 6Cheeseman IM Chappie JS Wilson-Kubalek EM Desai A 2006121 The conserved KMN network constitutes the core microtubule-binding site of the kinetochore.Cell 12750092-867498399710.1016/j.cell.2006.09.03917129783 · doi ↗ · pubmed ↗
- 7Ciferri C Pasqualato S Screpanti E Varetti G Santaguida S Dos Reis G Maiolica A Polka J De Luca JG De Wulf P Salek M Rappsilber J Moores CA Salmon ED Musacchio A 200852 Implications for kinetochore-microtubule attachment from the structure of an engineered Ndc 80 complex.Cell 13330092-867442743910.1016/j.cell.2008.03.02018455984 PMC 4754795 · doi ↗ · pubmed ↗
- 8de Regt Anna K. Clark Cordell J. Asbury Charles L. Biggins Sue 2022420 Tension can directly suppress Aurora B kinase-triggered release of kinetochore-microtubule attachments Nature Communications 1312041-172310.1038/s 41467-022-29542-8PMC 902126835443757 · doi ↗ · pubmed ↗