Phragmothecacentinelensis (Malvaceae, Malvoideae or Matisioideae), a newly-discovered, critically-endangered canopy tree species from a cloud forest in Pacific Ecuador

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5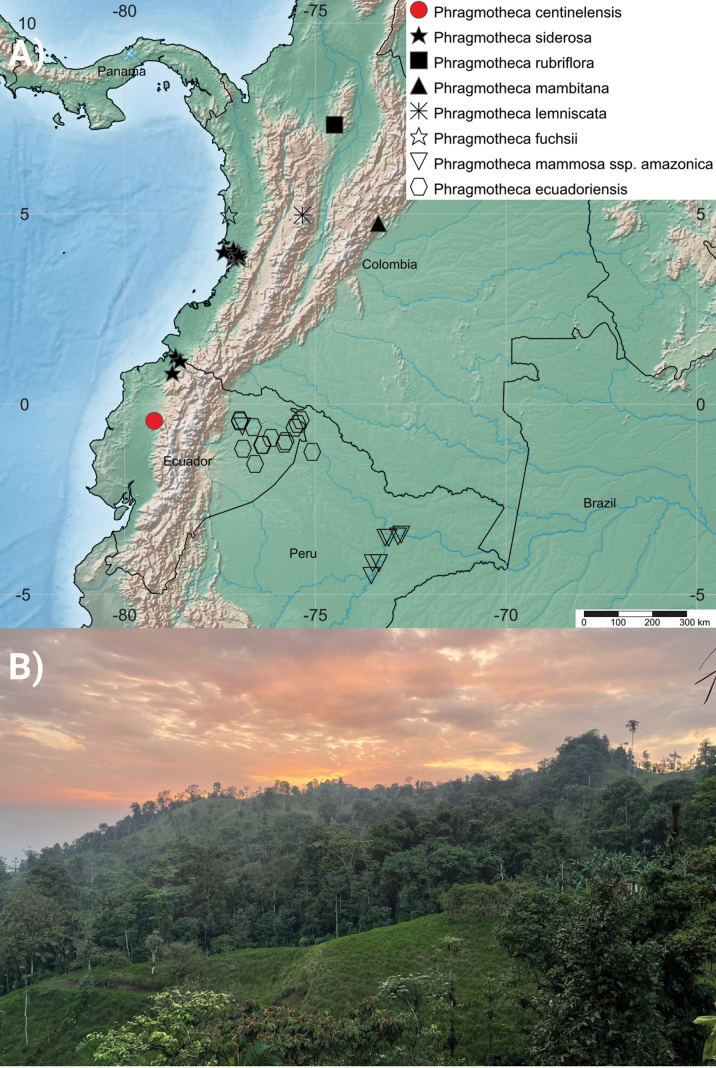 Figure 6
Figure 6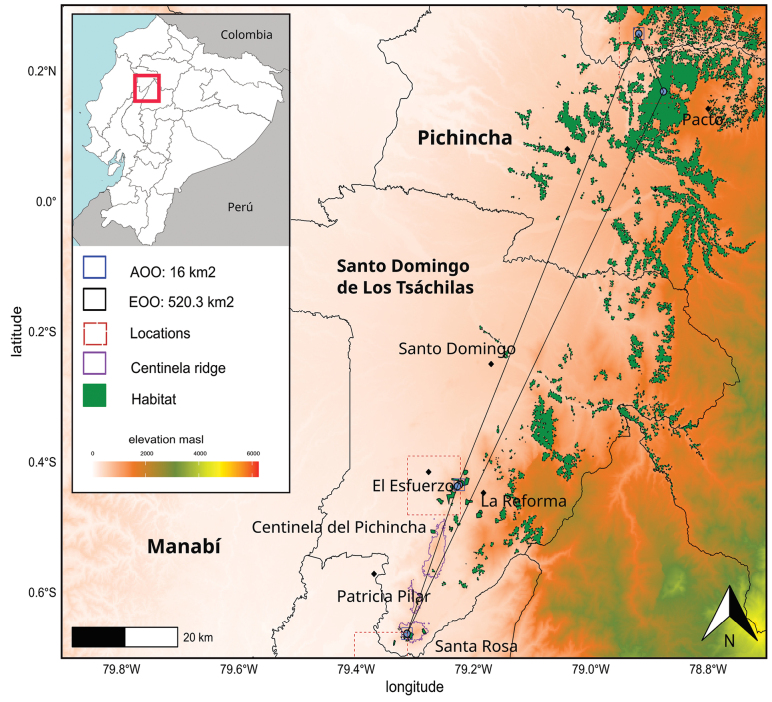 Figure 7
Figure 7| Species | Publication | Distribution | Country | Altitude (m) |
|---|---|---|---|---|
| 1946 | Chocoan | < 50 (–620) | ||
| 1971 | Chocoan |
| < 50 | |
| 1982 | Amazonian | 150–260 | ||
| 1991 | Amazonian | 115–150 | ||
| 1991 | Amazonian |
| 250–450 | |
| 1991 | Chocoan | 100–800 | ||
| 1996 | Chocoan |
| 50–100 | |
| 1996 | Chocoan |
| 180 | |
| 1996 | Andean |
| 750 | |
| 1996 | Amazonian |
| 320–700 | |
| 1999 | Cordilleran |
| 1700–1800 |
| Characters |
|
|
|
|
|
|---|---|---|---|---|---|
| Leaf blade shape | orbicular to oblong-elliptic | cordate-orbicular | broadly elliptic to suborbicular | lanceolate-oblong to narrowly obovate-oblong | ovate to elliptic or broadly elliptic |
| Abaxial leaf surface indument | sparsely distributed long-branched fasciculate hairs, lepidote-stellate scales and stellate-fasciculate hairs | densely covered by long-branched fasciculate hairs | densely covered by lepidote scales | stellate-lepidote | densely echinate-stellate |
| Primary leaf veins at base (not counting submarginals) | 5 | 5 | 3 | 2 | 5 |
| Secondary leaf veins (per side) | 5 | 4–5 | 4–5 | 6–8 | 4–5 |
| Calyx tube shape | narrowly campanulate | cylindrical | tubular-fusiform | tubular-cylindrical | tubular |
| Flower length, including pedicel (cm) | 3.3–5.2 | 2.8–2.9 | > 8 | 6.5–7.0 | ca. 9.0 |
| Petal colour | orange or pink | white or yellowish-white | white or yellowish-white | red | white or pale yellowish-brown |
| Staminal column length, with lobes (cm) | 2.9–3.6 | 1.7 | 7.0–7.2 | 6.5 | ca. 8.0 |
| Staminal column lobe length (cm) | 1.3–1.6 | 0.9–1.0 | 1.1–1.3 | 1.2–1.3 | 1.5 |
| Thecae per staminal column lobe | 6 (relatively uniform in size) | 3 (2 large plus 2 basal that are shared with adjacent lobes) | 4–6 (quite variable in size) | 7 (three pairs per lobe plus one basal that is shared) | – |
| Number of fovea (septa) per theca | 13–14 | 15–24 | few – many (> 50) | many | – |
| Fruit shape, length and width (cm) | ovoid to broadly ovate, 5.9–7.0 × 4.0–5.3 | broadly ovate, 5.2 × 6.0–6.2 | spherical, 7.0 × 4.0–4.5 | apparently spherical, ca. 4.0–3.8 | broadly ellipsoid to sub-obovoid, 6.5–8.0 × 6.3–6.8 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Diversity and Evolution · Ecology and Vegetation Dynamics Studies · Plant and animal studies
Introduction
The genus Phragmotheca Cuatrec. (Malvaceae, Malvoideae or Matisioideae) comprises 11 known species (Table 1): four in lowland Amazonian forests of Colombia, Ecuador and Peru; two in the Andean highlands of Colombia; and five in low- and mid-elevation Pacific-slope forests of Colombia and eastern Panama (Alverson 1991; Fernández-Alonso 1996; Fernández-Alonso & Jaramillo-Mejía 1999, Fernández-Alonso et al. 2017). Species are generally large canopy trees (to 50 m) with the exception of P.lemniscata Fern.Alonso, which is a treelet in the Chocó Region of Colombia. Phragmotheca is closely related to Matisia Bonpl. and Quararibea Aubl. The three genera consistently form a clade in phylogenetic analyses (/Matisieae in Baum et al. (1998, 2004)) and have been placed in the Matisieae K.Schum. (Schumann 1895; Reveal 2012) within the Malvoideae (sensu Baum et al. (1998, 2004); the /Malvoideae clade of Alverson et al. (1999)). A new revision of ranked names within Malvaceae (Colli-Silva et al. 2025) proposes that the /Matisieae clade be renamed Matisioideae, but one of us [WSA] considers this change to be unjustified at present (see Appendix 1). Phragmotheca differs from the other two genera by its conspicuous internal partitioning of staminal thecae into foveae or septa (cf. fig. 1 in Alverson (1991) and figs 4, 8, 9 and 12 in Fernández-Alonso (1996)), a trait that also is found in other “basal” genera of the /Malvatheca clade sensu Baum et al. (1998), i.e. the clade comprising the /Bombacoideae and /Malvoideae clades). Phragmotheca has long apical staminal lobes and five carpels, as does its presumably closest relative Matisia. (Quararibea has short apical staminal lobes and 2–4 carpels). However, unlike Matisia, Phragmotheca species also have thicker, prominently ribbed endocarps surrounding the five seeds (cf. fig. 1 in Fernández-Alonso (1996) and Fig. 5E, F).
Phragmotheca is a conspicuous floristic element of the humid forests of the Chocó Region of Colombia and Ecuador and the western Amazon. It has been hypothesised that lowland humid forests of Panama, Colombia and Ecuador constitute its centre of diversification (Gentry 1986; Fernández-Alonso 1998). Forests of the Chocó Region of Ecuador are subject to one of the highest deforestation rates in the world. Few remnants persist, accounting for only 15% of the original vegetation cover (Finer and Mamani 2019). Since 2021, we have been carrying out floristic inventories in one of the most iconic and imperilled areas within the Chocó Region of Ecuador: the Centinela (or Montañas de Ila) area. Originally this area had ~ 500 km^2^ of forest, but less than 7 km^2^ remains. In the last 10 years, several new tree species have been described from material collected in Centinela highlighting the relevance of this area for botanical exploration (Torke and Perez 2013; Cornejo 2023; Fernández-Alonso and Cornejo 2024).
During one of our inventories, we collected a specimen of Phragmotheca that exhibits remarkable characteristics, clearly different from those of known species in the Chocó (Table 2). This is the first new species of Phragmotheca described in the 21^st^ century (Table 1). Thus, our results highlight the importance of botanical exploration and conservation not just in poorly-collected sites in the Neotropics, but specifically in Neotropical cloud-forest remnants.
Methods
After our initial fieldwork during the floristic inventories, we collected additional, fertile specimens of this unknown species of Phragmotheca. We also made an exhaustive search of Phragmotheca specimens deposited in Ecuador’s three largest herbaria: Herbario de la Universidad Católica del Ecuador (QCA), Herbario Alfredo Paredes de la Universidad Central del Ecuador (QAP) and Herbario Nacional del Ecuador (QCNE; abbreviations according to Thiers (2019)). To gather habit and morphological information, we consulted digitised specimen images of the genus Phragmotheca available at the Field Museum virtual herbaria (https://collections-botany.fieldmuseum.org) and 339 images from an archive maintained by one of us [WSA], with additional specimens seen at AMAZ, COL, F, GUAY, HUA, JAUM, MEDEL, MO, MOL, PMA, SCZ, USM and WIS.
To determine the range of the new species for a conservation status assessment using IUCN criteria, we estimated the Extent of Occurrence and Area of Occupancy using the package conR (Dauby et al. 2017). Habitat fragmentation was estimated using the most current deforestation data available for western Ecuador from the online platform Mapa Interactivo (MAATE 2024). We first downloaded ecosystem layers for western Ecuador, as well as deforestation maps for the period 1990–2022. Habitat reduction was then assessed by combining these layers with the layers of Extent of Occurrence (EOO) and Area of Occupancy (AOO) using the clip tool in the ArcGis software (ESRI 2011). The clip tool overlays a range size map of EOO (Extent of Occurrence) and AOO (Area of Occupancy) with boundary layers, which, in this case, include deforestation scenarios from 1990 to 2022 and ecosystem layers. This process refines the potential habitat by limiting it to forest fragments. Habitat loss was then calculated as the area outside the combined boundaries defined by the deforestation and ecosystem layers. Conservation assessment was done following IUCN Standards and Petitions Committee (2022).
Results
Taxonomic treatment
Phragmotheca
centinelensis
Taxon classificationPlantaeMalvalesMalvaceae
J.C.Cerón, A.Fernández & J.E.Guevara sp. nov.
B7C40E4A-0646-5C97-95AF-9DF1B6BD342C
urn:lsid:ipni.org:names:77359001-1
Diagnosis.
The new species is morphologically similar to Phragmothecahydra Fern.Alonso, but differs by its orbicular to oblong-elliptic (vs. orbicular-cordate) leaves with a mixture of long-branched fasciculate hairs and lepidote-stellate scales in the axils of the mid-vein and secondary veins on the abaxial leaf surface (vs. fasciculate hairs only); larger flowers (3.5–5.0 [including the pedicel] × 2.9–3.5 vs. 2.8–2.9 × 2.8–3.0 cm) with glabrous, concave-spoon-like (vs. linear spathulate) petals; longer, glabrous staminal column (2.9–3.6 vs. 1.7 cm and densely covered by stellate hairs); staminal lobes each bearing 6 thecae (vs. 3–4 thecae); glabrous (vs. sparsely covered with long-branched, fasciculate hairs) style; subcapitate (vs. subacute) stigma; narrower, patelliform fruiting calyx (3.3–3.9 vs. 4–4.5 cm in diameter); and ovoid (vs. globose) fruits that are proportionately more slender (5.0–7.0 × 4.0–5.3 vs. 5.2 × 6.0–6.2 cm in diameter).
The semi-cordate leaf bases of this new species resemble those of Phragmothecalemniscata Fern.Alonso, but P.centinelensis differs by its larger size (canopy trees to 35 m vs. small treelets); indument of abaxial leaf surfaces (sparse fasciculate hairs, lepidote-stellate scales and stellate-fasciculate hairs vs. dense lepidote scales); floral calyces (narrowly campanulate vs. tubular-fusiform); petals (orange or pink, glabrous, glossy, distally concave, non-reflexed, with slightly convolute margins vs. white-cream, linear-spathulate, distally reflexed, internally covered by a mixture of stellate and lepidote scales); shorter staminal column (2.9–3.6 vs. 7.0–7.2 cm long, including the apical lobes) with apical lobes differing in indument (densely covered by septate simple hairs vs. glabrous); fruiting calyces (patelliform and covered by lenticels vs. discoid sub-cupular); and fruits (broadly ovoid vs. globose).
Phragmothecacentinelensis J.C.Cerón, A.Fernández & J.E.Guevara, sp. nov. Image of the holotype at QCNE (J.C. Cerón, A. Fernández, D. Navas, & L. Narváez 4643).
Type.
Ecuador – Santo Domingo de los Tsáchilas • (fl, fr, J.C. Cerón, A. Fernández, D. Navas, & L. Narváez 4643 (holotype: QCNE!, isotypes: F!, MO!, QCA!, WIS!)); Parroquia El Esfuerzo, Recinto Milton Murillo, Finca del Sr. Marlon Sarango, fragmento de bosque intervenido rodeado de pastizales; 0°35'54.53"S, 79°13'27.23"W; alt. 824 m; 01 Nov 2023.
Description.
Canopy trees to 25–35 m tall; trunk cylindrical, longitudinally fissured and forming thin plates, buttressed to 7 m height (Fig. 2A), outer bark reddish with dispersed, irregular grey spots (Fig. 2B), internal bark fibrous (outer half reddish-brown, inner half yellow-cream; Fig. 2C); branches: main branches off trunk verticillate, smaller branches glabrous, terete, longitudinally fissured, with granular lenticels. Stipules 2–3 mm long, broadly triangular, caducous. Leaves alternate, clustered at the tips of the branchlets; petioles 3.6–12.2 cm long, 1–3.5 mm in diameter, terete, finely striate, densely pubescent, pulvinate at both ends. Leaf blades (Fig. 3A, B) glaucous when young, pale green abaxially when mature, coriaceous, entire, orbicular to oblong-elliptic, (6–)6.7–31.1(32) × (5–)7.6–29.3(–30) cm, semi-cordate to deeply cordate at base with slightly asymmetric lobes 0.5–4.3 cm depth, the apex obtuse to shortly apiculate; the abaxial surface densely covered by a mixture of lepidote-stellate scales, stellate-fasciculate trichome and fasciculate trichomes; venation conspicuous on the abaxial surface; primary veins palmate near leaf base (Fig. 3D) with 5 basal nerves, 3–4 submarginal nerves slightly ascending to the leaf margin and with 5 pairs of secondary veins arising from mid-rib in distal 2/3 of blade, these with barbate tufts of fasciculate trichomes and sessile lepidote-stellate scales in their axils (Fig. 3G); tertiary venation also prominent on the abaxial surface (Fig. 3E), forming a conspicuous reticulum (inconspicuous on the adaxial surface), with golden lepidote-stellate scales on the lamina between nerves (Fig. 3F). Flowers slightly zygomorphic, solitary (Fig. 4A, B), borne opposite to leaves on short branches (brachyblasts), 3.3–5.2 cm long (including the pedicels), the pedicels 1.2–1.4 cm long, 0.3–0.4 cm in diameter, finely striate, covered with sessile stellate-lepidote scales and fasciculate hairs; floral bracts absent below the calyx. Calyx narrowly campanulate (Fig. 4C), 2.3–3.3 × 0.9–1.2 cm, densely covered by a brownish, floccose indument (imparting a granular appearance), broadly acuminate and 4-lobed at summit, internally covered with adpressed sericeous hairs. Petals spathulate, distinctly bilobed at apex, strongly concave distally (Fig. 4B, D), the apex obtuse or rounded, the margins slightly convolute, 3.5–4.1 × 1.5–2.0 cm, orange or pink, glabrous, glossy, 2–4 inner corolla lobes covering the staminal tube and non-reflexed corolla tube lobes. Staminal column 2.9–3.6 cm long, the tube (16.5–)16.8–20.6(–21.0) × 0.4–0.6 mm long, orange, with 5 linear terminal lobes (Fig. 4G), each lobe 12.9–15.8 × 2.5–2.8 mm, orange or pink and bearing 6 thecae in two parallel lines on the adaxial surface, these surrounded by long, unbranched, septate hairs (Fig. 4G–I), with 14–16 foveae per theca. Ovary elongate, 1.5–1.7 mm long, 5-carpellate, glabrous, the style not exceeding staminal filaments in length, longitudinally 6-sulcate, the stigma capitate, spongy, glandular. Fruiting pedicel short, terete, 7.3–8.9 × 5.8–6.1 mm. Fruiting calyx patelliform and slightly dentate (Fig. 5A, B), surrounding only the basal 1/5 of the fruit and beset with cream-coloured lenticels. Fruit ovoid to broadly ovoid, 5.9–7 × 4–5.3 cm, the exocarp pale, smooth, finely fissured longitudinally (Fig. 5C) cream-coloured, sparsely granular with short, fasciculate hairs; the mesocarp fibrous-pulpy, white or cream-coloured and exuding yellowish mucilage (Fig. 5D, E), the endocarp densely fibrous, forming a woody pyrene with each of the five seeds (Fig. 5F). Mature seeds elliptic, 4–4.8 × 1.7–2 cm.
Phragmothecacentinelensis J.C.Cerón, A.Fernández & J.E.Guevara A trunk B outer bark C inner bark. Photos of the type individual (Cerón et al. 4643) by Andrea Fernández and Juan Carlos Cerón.
Phragmothecacentinelensis J.C.Cerón, A.Fernández & J.E.Guevara A leaves, adaxial surface B leaves, abaxial surface C terminal branch showing the terminal bud and lenticels D basal leaf veins, abaxial surface (8×) E tertiary and quaternary venation, abaxial surface (8×) F stellate-lepidote scales, abaxial surface of leaf (100×) G barbate axil of mid-vein and secondary vein, with tufts of long-branched fasciculate hairs and stellate-lepidote scales, abaxial leaf surface. Photos of the type individual (Cerón et al. 4643) by Andrea Fernández and Juan Carlos Cerón.
Phragmothecacentinelensis J.C.Cerón, A.Fernández & J.E.Guevara A flowering branch B flower with reflexed, concave-spoon-like petals C campanulate flowering calyx D distal part of petal showing the diagnostic concave form that is characteristic of this new species E flower bud (8×) F close-up of flower bud indument, a mix of stellate-lepidote scales and fasciculate hairs G apical lobes of staminal column (note sterile tips), each with two parallel rows of thecae and the shorter style in centre H close-up of apical lobes with elongate, foveate thecae (15×); I elongate, septate hairs on the thecae (100×).
Phragmothecacentinelensis J.C.Cerón, A.Fernández & J.E.Guevara A fruiting branch B patelliform fruiting calyx C apex of the fruit showing the fine longitudinal fissures D longitudinal view of the fruit showing yellow exudate from the mesocarp E transverse section of the fruit showing the exocarp, fibrous mesocarp and very thick endocarp tissue surrounding the seeds F two pyrenes (seeds with some residual fibrous-woody endocarp and mesocarp tissue).
Phenology.
Flowering from November to December; fruiting between February and March.
Distribution.
Phragmothecacentinelensis is a large tree of non-flooded habitats in the Chocó cloud forests of Ecuador occupying a narrow altitudinal band between 600 and 1000 m elevation. It occupies mostly highly dissected terrain in the ridges of the Centinela area (Figs 6, 7). According to our tree species inventory, P.centinelensis appears to be relatively rare in the region and possibly endemic to the Ecuadorian Chocó; the only two known populations are in cloud forest remnants in the western Andean foothills of Centinela. Although we know of no record of P.centinelensis from Colombia, it may occur in similar habitats of the adjacent Chocó Department. Despite its rarity, distinctive features make it conspicuous in the field, notably the large buttresses up to 7 m height, its reddish outer bark and the longitudinal fissures on the trunk.
A Distribution of Phragmothecacentinelensis J.C.Cerón, A.Fernández & J.E.Guevara, its relatives in the Chocó Region of Ecuador and Colombia and two morphologically similar Amazonian species B cloud-forest habitat of the new species in Ecuador.
Map showing the Extent of Occurrence (EOO), Area of Occupancy (AOO) and available habitat of Phragmothecacentinelensis J.C.Cerón, A.Fernández & J.E.Guevara in western Ecuador. Inset: Location of Santo Domingo de las Tsáchilas Province within Ecuador.
Habitat and ecology.
Some of the most conspicuous floristic elements of the Centinela cloud forests include the families Fabaceae, Lecythidaceae and Malvaceae with dominant tree species including Ocoteainsularis (Meisn.) Mez, Ruageaglabra Triana & Planch., Guatteria sp., Aegiphilaalba Moldenke, Trichiliasurinamensis (Miq.) C.DC., Socrateaexorrhiza (Mart.) H.Wendl. and Iriarteadeltoidea Ruiz & Pav. The genera Eschweilera Mart. ex DC., Inga Mill., Gustavia L., Matisia, and Quararibea are also both dominant and species-rich groups in this area. It is remarkable that Phragmothecacentinelensis co-exists locally with several other species within the /Matisieae clade, including M.palenquiana (A.Robyns) W.S.Alverson, M.coloradorum Benoist, M.giacomettoi Romero, M.castano H.Karst. & Triana and Q.casasecae Fern.Alonso & Castrov., Q.grandifolia (Little) Cuatrec. perhaps due to similar dispersal mechanisms or pollination syndromes. Natural regeneration of P.centinelensis seems to be rare in the forest fragments of the Centinela area. Gravity seems to be the prominent mechanism of primary dispersion. We found a high density of fruits and seeds underneath parent trees with abundant signs of predation by rodents or other small mammals, nearly all fruits with bite marks. There may be secondary dispersal due to scatter-hoarding by these mammals. The heavily armoured endocarps may be an adaptation to facilitate this, but more study is needed. Our current observations suggest a high mortality of seeds and seedlings due to a negative density dependence process via Janzen-Connell phenomena (Forrister et al. 2019).
Conservation status.
Phragmothecacentinelensis is known from only three confirmed populations in the western foothills of the Ecuadorian Andes in the few remnants of cloud forests of the Montañas de Ila, more specifically in areas outside of Centinela Ridge (Fig. 7). This small population, within a 150-ha patch of forest, represents the type locality. In this area, only three adult individuals have been observed, all of them with a diameter at breast high > 30 cm. Another subpopulation of four individuals has been reported at the southern end of the Centinela Ridge, ca. 15 km from the confirmed population. Based on field observations by other botanists, we believe there to be a second population occurring in a large fragment of forest (2500 ha) in the private Mashpi Reserve, 85 km north of Centinela (A. Pérez and G. Toasa, pers. comm.). We reviewed a sterile Phragmotheca herbarium specimen from the Mashpi Reserve and confirmed it belongs to the new species described herein. A third population is located in small patch of cloud forest at 1100–1200 m elevation, near the town of Cielo Verde in Imbabura Province, approximately 15 km north of Mashpi Reserve. This forest is part of the buffer zone from Los Cedros Reserve with approximately 4500 ha protecting the last remnants of Chocó cloud forests in western Ecuador.
Our estimates of extent of occurrence (EOO) and area of occupancy (AOO) are 520 km^2^ and 16 km^2^, respectively (Bachman et al. 2011) (Fig. 7). Two of the three known confirmed populations of Phragmothecacentinelensis are not formally protected in the Ecuadorian Protected Area National System (SNAP). Our analysis demonstrates that, from 1990 to 2022, deforestation across the range of this species has reduced its EOO and AOO in 21% approximately. Furthermore, based on estimated EOO and AOO (Fig. 7) and historic deforestation over the last 34 years, we found that 34% of its potential habitat has disappeared. Thus, on the basis of current deforestation rates in the Chocó Region (Finer and Mamani 2019) and habitat loss, our assessment of P.centinelensis’s global threat status is Endangered [EN] under IUCN Criterion EN B1ab(i,ii,iii)+2ab(i,ii,iii). It is important to note that historical deforestation of the Centinela Ridge predates the 1970s, but several large forest fragments remained interspersed in a matrix of cacao, balsa and Gmelina L. plantations (Dodson and Gentry 1991). However, at present, deforestation was occurring even during our visits: at the time of our floristic inventories in 2021, one of the three adult individuals which we recorded was being removed by loggers.
Etymology.
We named the new species in honour of the iconic site that the renowned botanists Alwyn Gentry and Calaway Dodson visited more than 40 years ago in the Centinela area, close to Santo Domingo de los Colorados in the western foothills of the Ecuadorian Andes. At the time they visited an area known as Centinela del Pichincha, they observed a severely fragmented landscape, which led them to conclude that ongoing deforestation had wiped out almost all the remaining forests in the region. After the publication of their seminal paper (Dodson and Gentry 1991), the term “Centinelan extinction” was coined and popularised by Wilson (1992). The term aimed to reflect the global extinction of a high number of endemic plant species, many of them undescribed, following high levels of forest fragmentation.
Additional specimens examined.
Ecuador – Santo Domingo de los Tsáchilas • J.C. Cerón et al. 4684 (QCA, QCNE, F); Parroquia El Esfuerzo, Recinto Milton Murillo, Finca del sr. Marlon Sarango. bosque siempreverde piemontano de la cordillera occidental de los Andes (BsPn01), Fragmento de bosque intervenido rodeado de pastizales; 0°26'15.35"S, 79°13'38.57"W; alt. 840 m; 01 Nov 2023; fl • Santo Domingo de los Tsáchilas: J.C. Cerón et al. 4688 (QCA, QCNE, F); Parroquia Centinela del Pichincha, Comunidad Santa Rosa, bosque siempreverde piemontano de la cordillera occidental de los Andes (BsPn01), bosque maduro intervenido; 0°39'51.15"S, 79°18'48.37"W; alt. 682 m; 01 Nov 2023; fr. • Pichincha: Reserva Masphi • PDRBA 43 (QCA); Colecciones en la parcela 7 de 60 × 60 m del transecto altitudinal para el monitoreo de la dinámica forestal. Bosque Montano Bajo. Árbol, placa 7294; 0°10'6.63"N, 78°52'34.2402"W; alt. 800–900 m; 2–25 Aug 2019. • Imbabura: Cielo Verde • PDRBA 285 (QCA); Colecciones en la parcela 7 de 60 × 60 m del transecto altitudinal para el monitoreo de la dinámica forestal. Bosque Montano Bajo. Árbol, placa 586; 0°15'26.2074"N, 78°55'6.096"W; alt. 1100–1200 m; 10–22 Jul 2021.
Discussion
Based on morphology and geographic distribution, Phragmothecacentinelensis is a new species distinct from other morphologically similar congeneric species in the Chocó and Amazon Regions.
Phragmothecacentinelensis is one of many new species described from remnant forests in western Ecuador over the last 15 years (Cornejo 2009a, 2009b,; Palacios 2012; Torke and Perez 2013; Guevara-Andino and Fernández-Fernández 2020; Couvreur et al. 2022; Fernández-Alonso and Cornejo 2024; Clark et al. 2024). Its discovery adds valuable new insight into the region’s conservation value and floristic richness, as well as providing another piece of the puzzle to understand the historical biogeography and evolution of members of the /Matisieae clade (the > 100 species included in the genera Matisia, Phragmotheca and Quararibea). For example, M.palenquiana (A.Robyns) W.S.Alverson was described (as a Quararibea) from specimens collected in 1974 by Gentry and Dodson at the Río Palenque Biological Station, very close (60 km) to the type locality of P.centinelensis. The diagnosis in the original article (Robyns 1976) notes the similarity of the new species to M.bicolor Ducke, an Amazonian species. A third, very closely-related species, with similarly distinctive flowers and fruits, was recently discovered in Costa Rica notwithstanding many decades of dedicated botanical collecting efforts there: M.tinamastiana A. Estrada & Cascante (Estrada and Cascante (1998), with an updated distribution in Fernández-Alonso and Campos-Pineda (2023)). Triplets of closely-related species distributed east of the Andes in the Amazon, west of the Andes in Colombia and Ecuador and northwest into isthmian Costa Rica and Panama are evident in several genera of the /Bombacoideae and /Malvoideae clades (sensu Baum et al. (1998, 2004)), which invites further investigation once additional molecular phylogenies for these genera are available.
Conclusion
Phragmothecacentinelensis is a distinct and endangered new species of canopy tree found in forest remnants of western Ecuador. Discovery and documentation efforts for rare species such as this are critical to their conservation and to the value they add to our understanding of floristics, historical biogeography and the evolutionary diversification of Neotropical cloud forests.
Supplementary Material
XML Treatment for Phragmotheca centinelensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alverson WS (1991) A synopsis of Phragmotheca (Bombacaceae), with two new species and a new subspecies.Brittonia 43(2): 73–87. 10.2307/2807297 · doi ↗
- 2Alverson WS Whitlock BA Nyffeler R Bayer C Baum DA (1999) Phylogeny of the core Malvales: Evidence from ndh F sequence data.American Journal of Botany 86(10): 1474–1486. 10.2307/265692810523287 · doi ↗ · pubmed ↗
- 3Bachman S Moat J Hill Ade la Torre J Scott B (2011) Supporting Red List threat assessments with Geo CAT: Geospatial conservation assessment tool.Zoo Keys 150: 117–126. 10.3897/zookeys.150.2109 PMC 323443422207809 · doi ↗ · pubmed ↗
- 4Baum DA Alverson WS Nyffler R (1998) A durian by any other name: Taxonomy and nomenclature of the Core Malvales.Harvard Papers in Botany 3(2): 315–330.
- 5Baum DA De Witt Smith S Yen A Alverson WS Nyffeler R Whitlock BA Oldham RL (2004) Phylogenetic relationships of Malvatheca (Bombacoideae and Malvoideae; Malvaceae sensu lato) as inferred from plastid DNA sequences.American Journal of Botany 91(11): 1863–1871. 10.3732/ajb.91.11.186321652333 · doi ↗ · pubmed ↗
- 6Carvalho-Sobrinho JG Alverson WS Alcantara S Queiroz LP Mota AC Baum DA (2016) Revisiting the phylogeny of Bombacoideae (Malvaceae): Novel relationships, morphologically cohesive clades, and a new tribal classification base on multilocus phylogenetic analyses.Molecular Phylogenetics and Evolution 101(2016): 56–74. 10.1016/j.ympev.2016.05.00627154210 · doi ↗ · pubmed ↗
- 7Clark JL Fernández A Zapata JN Restrepo-Villarroel C White DM Pitman N (2024) Amalophyllonmiraculum (Gesneriaceae), an exceptionally small lithophilous new species from the western Andean slopes of Ecuador.Phyto Keys 242: 307–316. 10.3897/phytokeys.242.11806938903848 PMC 11188077 · doi ↗ · pubmed ↗
- 8Colli-Silva MPérez-Escobar OA Ferreira CDM Costa MTR Gerace S Coutinho TS Yoshikawa VN Antonio-Domingues H Hernández-Gutiérrez R Bovini MG Duarte MC Cheek M Chase MW Fay MF Christenhusz MJM Dorr LJ Schoeplein C Cocoran M Roy S Cable S Mc Lay T Maurin O Forest F Baker WJ Antonelli A (2025) Taxonomy in the light of incongruence: An updated classification of Malvales and Malvaceae based on phylogenomic data. Taxon, Early View [published 23 Jan 2025]. 10.1002/tax.13300 · doi ↗