Morphometric synthesis of Pollimyrus (Teleostei, Mormyridae) with the description of four new species
Katrien Dierickx, Soleil Wamuini Lunkayilakio, Roger Bills, Emmanuel Vreven

TL;DR
This study identifies four new species of Pollimyrus fish in Africa and suggests some species may need to be grouped together.
Contribution
The paper describes four new Pollimyrus species and re-evaluates species boundaries using morphometric analysis.
Findings
Four new Pollimyrus species were identified and described from different African rivers.
Some species showed little morphological difference, suggesting possible synonymization.
Twenty morphologically distinct Pollimyrus species are currently recognized.
Abstract
Mormyridae, a species‐rich family endemic to Africa, remains taxonomically understudied. This has been the case for the genus Pollimyrus Taverne, 1971, which hinders further understanding of the distribution, ecology, and conservation of its species. Therefore, an in‐depth morphometric comparison of all currently valid species is carried out using most of the available type specimens. Species delineations were re‐evaluated, and four species new to science described: Pollimyrus ibalazambai sp. nov. (the Luki River, the Democratic Republic of the Congo), Pollimyrus krameri sp. nov. (the Lugenda River, Mozambique), Pollimyrus vanneeri sp. nov. (the Kouilou‐Niari River, the Republic of the Congo), and Pollimyrus weyli sp. nov. (the Buzi River, Mozambique). In this study, Pollimyrus guttatus is confirmed to belong to Pollimyrus, whereas Pollimyrus eburneensis and Cyphomyrus plagiostoma seem…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
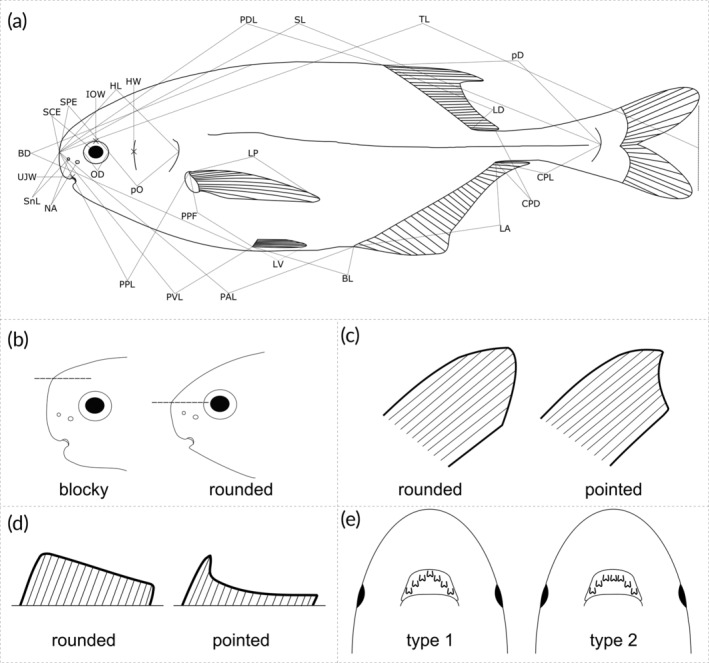 FIGURE 1
FIGURE 1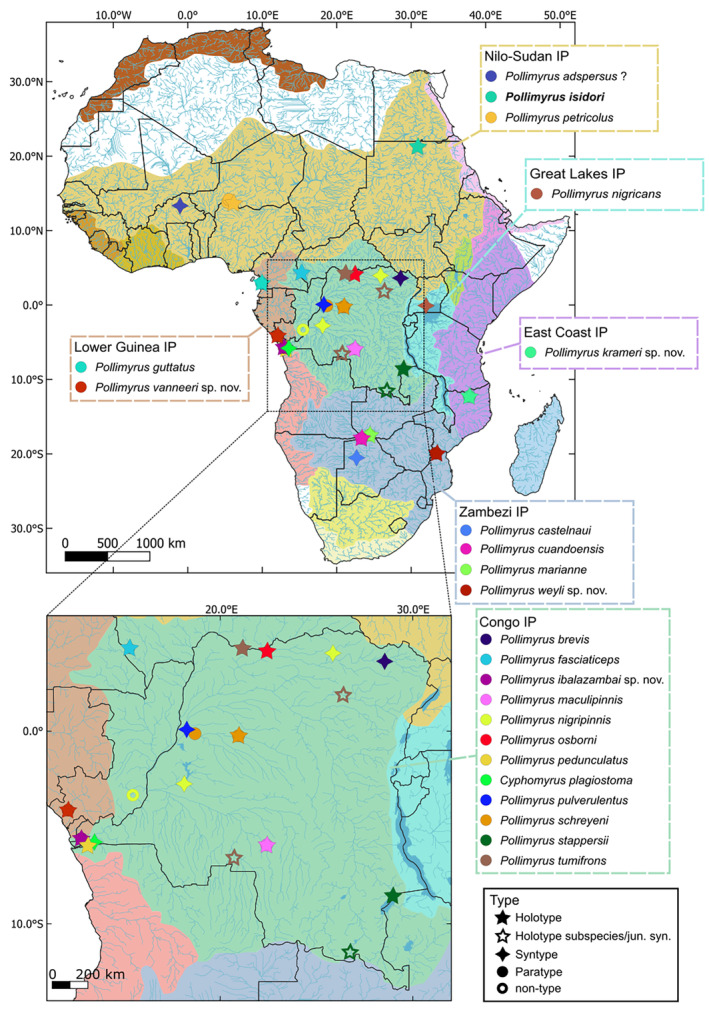 FIGURE 2
FIGURE 2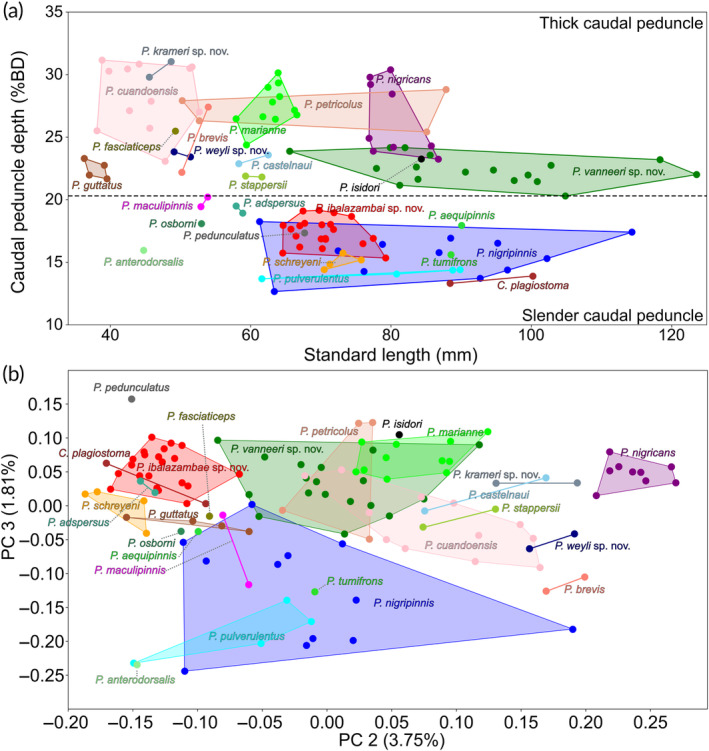 FIGURE 3
FIGURE 3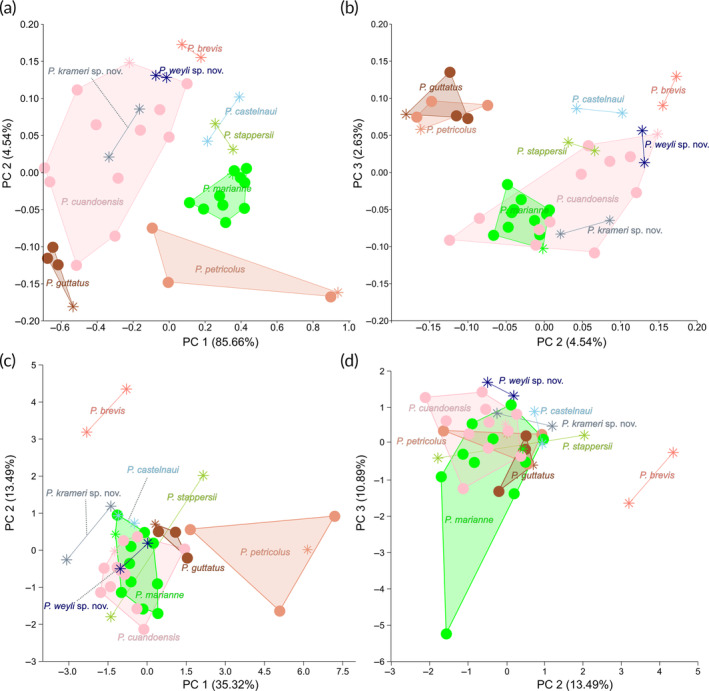 FIGURE 4
FIGURE 4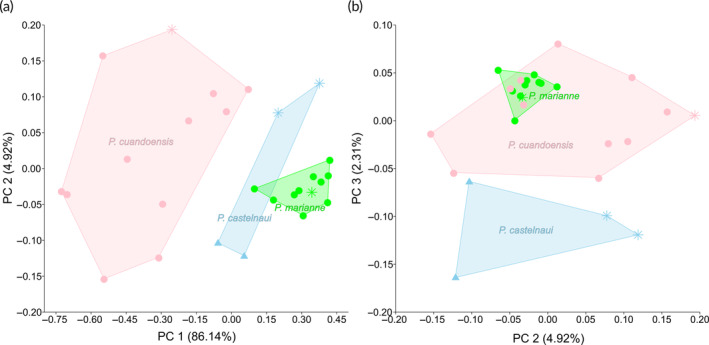 FIGURE 5
FIGURE 5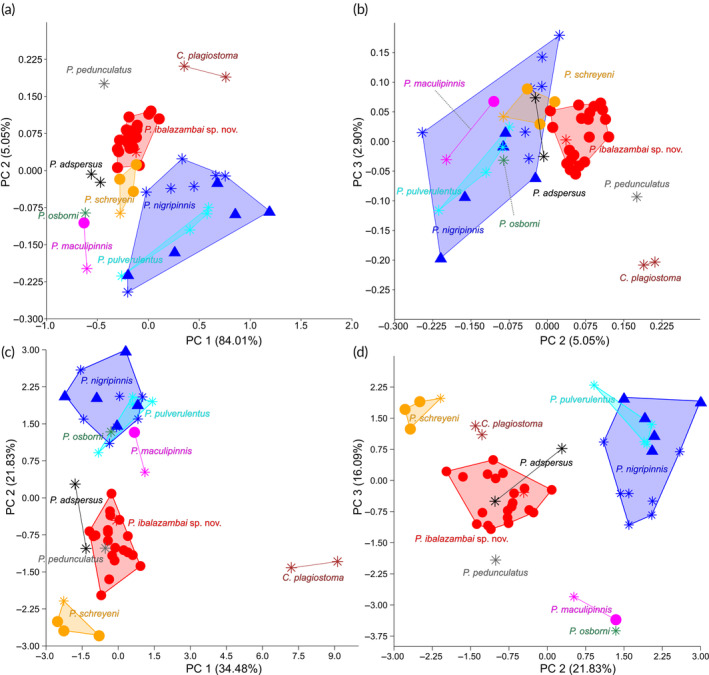 FIGURE 6
FIGURE 6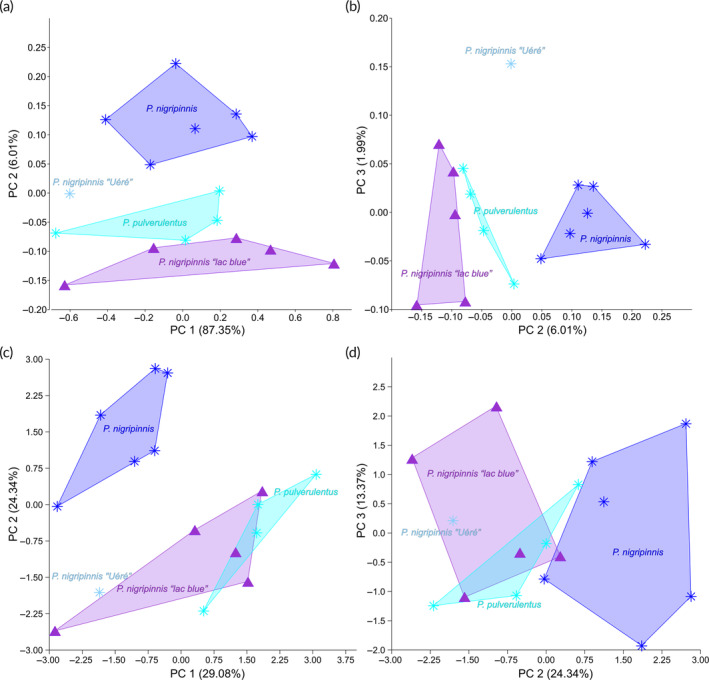 FIGURE 7
FIGURE 7 FIGURE 8
FIGURE 8 FIGURE 9
FIGURE 9 FIGURE 10
FIGURE 10 FIGURE 11
FIGURE 11 FIGURE 12
FIGURE 12 FIGURE 13
FIGURE 13 FIGURE 14
FIGURE 14 FIGURE 15
FIGURE 15 FIGURE 16
FIGURE 16 FIGURE 17
FIGURE 17| Species | Author(s) | Taverne, | Taverne, | Latest taxonomic revision | Current status | Type locality |
|---|---|---|---|---|---|---|
|
| Günther, |
|
| Taverne, 1971 |
| West Africa |
|
| Pellegrin, |
|
| Poll, |
| N'Gombe, Kasaï River |
|
| David & Poll, |
|
| Taverne, 1971 (jun. syn.) |
| Panga, Aruwimi River |
|
| Boulenger, 1913 |
|
| Taverne, 1971 |
| Dungu, Uele River |
|
| Boulenger, 1904 |
|
| Kramer & van der Bank 2011 (jun. syn.) |
| Assay, Niger River |
|
| Boulenger, 1900 |
|
| Hopkins et al., |
| “Kop‐Malafu,” Mayombe |
|
| Boulenger, |
|
| Taverne, 1971 |
| Lake Ngami |
|
| Kramer et al., | Kramer et al., |
| Kwando River | ||
|
| Boulenger, 1920 |
|
| Taverne, 1971 (subspecies) |
| Kinshasa, Pool Malebo |
|
| Pellegrin, 1909 |
|
| Taverne, 1971 (jun. syn.) |
| Bol, Lake Chad |
|
| Fowler, 1936 |
| Lavoué, 2010 |
| Kribi, Kineke River | |
|
| Boulenger, 1913 |
|
| Taverne, |
| Uele River |
|
| Valenciennes, 1847 |
|
| Taverne, 1971 |
| Nile River |
|
| Günther, 1896 |
|
| Hopkins et al., |
| Old Calabar, Ogooué River |
|
| Bigorne, | Rich et al., |
| Agnébi basin | ||
|
| Steindachner, 1870 |
|
| Bigorne, |
| Senegal |
|
| Non Steindachner | Bigorne, |
| Senegal | ||
|
| Boulenger, 1920 | Sullivan et al., |
| Poko, Bomokandi River | ||
|
| Fowler, 1936 |
|
| Bigorne, |
| Sibut |
|
| Nichols & La Monte, 1934 |
|
| Taverne, 1971 |
| Kananga, Lulua River |
|
| Sauvage, 1879 |
|
| Hopkins et al., |
| Doumé, Ogooué River |
|
| Kramer et al., | Kramer et al., |
| Zambezi River | ||
|
| Boulenger, 1906 |
|
| Taverne, 1971 |
| Mouth Katonga River |
|
| Boulenger, |
|
| Taverne, 1971 |
| Kutu, Lake Mai‐Ndombe |
|
| Nichols & Griscom, 1917 |
|
| Taverne, 1971 |
| Uele River |
|
| Steindachner, 1866 |
|
| Poll & Gosse, |
| Angola (Kwanza River?) |
|
| David & Poll, |
|
| Taverne, 1971 |
| Boma, Pool Malebo |
|
| Boulenger, 1898 |
|
| Levin & Golubtsov, |
| Khartum, Nile River |
|
| Daget, 1954 |
|
| Taverne 1971 |
| Markala |
|
| Boulenger, 1898 |
|
| Stiassny et al., |
| Matadi, Lower Congo River |
|
| Boulenger, |
|
| Taverne 1971 |
| Mbandaka, Middle Congo River |
|
| Svensson, 1933 |
|
| Taverne 1971 (jun. syn.) |
| Gambia River |
|
| Poll, 1972 |
| Taverne, |
| Boende |
| Species | Holotype | Paratype | Syntype | Non‐types | Total |
|---|---|---|---|---|---|
|
| 2 | 2 | |||
|
| 2 | 2 | |||
|
| 2 | 2 | |||
|
| 1 | 11 | 12 | ||
|
| 1 | 7 | 8 | ||
|
| 1 | 3 | 4 | ||
|
| 1 + 1 | 5 | 7 | ||
|
| 1 | 1 | 2 | ||
|
| 1 | 10 | 11 | ||
|
| 9 | 9 | |||
|
| 7 | 5 | 12 | ||
|
| 1 | 5 | 6 | ||
|
| 1 | 1 | |||
|
| 1 | 3 | 4 | ||
|
| 2 | 2 | |||
|
| 4 | 4 | |||
|
| 1 | 3 | 4 | ||
|
| 1 + 1 | 2 | |||
|
| 1 + 1 + 1 | 3 | |||
|
| 1 | 17 | 18 | ||
|
| 1 | 1 | 2 | ||
|
| 1 | 20 | 21 | ||
|
| 1 | 1 | 2 | ||
| Total | 16 + 4 | 77 | 28 | 15 | 140 |
| Measurements (Figure | Abbrev. | Description |
|---|---|---|
| Standard length | SL | Distance from snout to base of caudal fin |
| Total length | TL | Distance between snout and end of tail fin |
| Body depth at pelvic fins | BD | Distance between pelvic‐fin origin and dorsum perpendicular to pelvic‐fin origin |
| Pre‐dorsal length | PDL | Distance between snout and dorsal‐fin origin |
| Pre‐anal length | PAL | Distance between snout and anal‐fin origin |
| Pre‐pectoral length | PPL | Distance between snout and pectoral‐fin origin |
| Pre‐pelvic length | PVL | Distance between snout and pelvic‐fin origin |
| Length of dorsal fin | LD | Distance between dorsal‐fin origin and end |
| Length of anal fin | LA | Distance between anal‐fin origin and end |
| Length of pectoral fin | LP | Distance between pectoral‐fin origin and tip |
| Length of pelvic fin | LV | Distance between pelvic‐fin origin and tip |
| Posterodorsal distance | pD | Distance between dorsal‐fin origin and posterior end of last vertebra |
| Pectoral–pelvic fin distance | PPF | Distance between pectoral‐ and pelvic‐fin origins |
| Caudal peduncle length | CPL | Distance between anal‐fin end and posterior end of last vertebra |
| Caudal peduncle depth | CPD | Distance between anal‐fin end and dorsal side of caudal peduncle perpendicular to anal‐fin end |
| Belly length | BL | Distance between pelvic‐ and anal‐fin origins |
| Head length | HL | Distance between snout and the most posterior border of opercle |
| Head width | HW | Distance between the most posterior borders of left and right opercles |
| Postorbital length | pO | Distance between posterior border of eye and most posterior side of opercle |
| Snout–eye posterior distance | SPE | Distance between snout and posterior side of orbit |
| Snout–eye center distance | SCE | Distance between snout and center of eye |
| Snout–anterior nostril length | Snl | Distance between snout and anterior nostril |
| Internasal length | NA | Minimum distance between both nostrils |
| Eye diameter | OD | Distance between the most anterior and the most posterior edge of eye |
| Interorbital width | IOW | Distance between both left and right most dorsal sides of eyes |
| Upper‐jaw width | UJW | Distance between left and right most posterior parts of upper jaw |
| Lower‐jaw width | LJW | Distance between left and right most posterior parts of lower jaw |
|
| ||
| Dorsal‐fin rays | Number of dorsal‐fin rays | |
| Anal‐fin rays | Number of anal‐fin rays | |
| Pectoral‐fin rays | Number of pectoral‐fin rays | |
| Pelvic‐fin rays | Number of pelvic‐fin rays | |
| Upper‐jaw teeth | Number of teeth in the lower jaw | |
| Lower‐jaw teeth | Number of teeth in the upper jaw | |
| Lateral‐line scales | Number of scales along the lateral line | |
| Transverse dorsal–lateral lines scales | Number of scales between dorsal‐fin origin and lateral‐line scales | |
| Transverse anal–lateral‐line scales | Number of scales between anal‐fin origin and lateral‐line scales | |
| Transverse pelvic–lateral‐line scales | Number of scales between pelvic‐fin origin and lateral‐line scales | |
| Circumpeduncular scales | Number of scales around the caudal peduncle at the level of anal‐fin end | |
| Vertebrae | Number of vertebrae, counted using X‐ray photographs |
| Morphological group | Species | Snout shape | Mental lobe | Pectoral‐fin shape | Dorsal‐ and anal‐fin shape | CPD (% BD) |
|---|---|---|---|---|---|---|
| Thick‐tailed group |
| Rounded | Small | Rounded | Rounded | 22.2–27.4 |
|
| Rounded | Small | Rounded | Rounded | 22.9–23.6 | |
|
| Rounded | Small | Rounded | Rounded | 23.1–31.2 | |
|
| Rounded | Small | Rounded | Rounded | 21.7–23.3 | |
|
| Rounded | Small | Rounded | Rounded | 24.4–30.1 | |
|
| Rounded | Small | Rounded | Rounded | 25.4–28.8 | |
|
| Rounded | Small | Rounded | Rounded | 21.8–21.9 | |
|
| Rounded | Small | Rounded | Rounded | 23.4–23.8 | |
|
| Rounded | Small | Rounded | Rounded | 23.4 | |
|
|
| Blocky | None | Pointed | Longer, rounded | 23.3 |
|
| Blocky | None | Pointed | Longer, rounded | 23.2–30.4 | |
|
| Blocky | None | Pointed | Pointed | 25.5 | |
|
|
| Blocky | None | Rounded | Pointed or rounded | 20.3–24.2 |
| Slender‐tailed group |
| Blocky | None | Pointed | Pointed | 18.9–19.5 |
|
| Blocky | None | Pointed | Pointed | 19.4–20.2 | |
|
| Blocky | None | Pointed | Pointed | 15.3–18.3 | |
|
| Blocky | None | Pointed | Pointed | 18.1 | |
|
| Blocky | None | Pointed | Pointed | 17.3 | |
|
| Blocky | None | Pointed | Pointed | 13.3–13.9 | |
|
| Blocky | None | Pointed | Pointed | 12.7–14.4 | |
|
| Blocky | None | Pointed | Pointed | 14.9–15.7 | |
|
| Blocky | None | Pointed | Pointed | 15.3–19.2 | |
|
|
| Rounded | None | Pointed | Pointed | 15.6 |
|
| Rounded | None | Pointed or rounded | Pointed | 16.0 | |
|
| Rounded | Small | Pointed | Pointed | 17.9 |
| Measurement |
|
|
|
|
|---|---|---|---|---|
| Characteristic (in % SL) | ||||
| BD | 30.0–33.4 | 28.4–30.7 | 28.2 | 30.7–35.7 |
| BL | 18.1–23.6 | 22.2–24.7 | 21.6 | 19.0–22.4 |
| PPL | 29.2–30.1 | 26.6–28.7 | 29.8 | 27.9–30.2 |
| LD | 22.8–25.4 | 20.9–23.6 | 18.5 | 22.3–24.8 |
| LA | 28.6–30.8 | 27.1–28.7 | 28.1 | 28.0–29.8 |
| LV | 11.3–12.3 | 9.9–11.1 | 11.8 | 10.8–12.4 |
| CPL | 14.0–16.3 | 16.4–18.1 | 16.5 | 13.6–17.3 |
| HL | 24.9–26.8 | 23.5–25.3 | 27.1 | 25.0–27.7 |
| Characteristic (in % BD) | ||||
| CPD | 13.7–14.4 | 15.3–16.9 | 18.3 | 12.7–17.4 |
| Characteristic (in % HL) | ||||
| SnL | 16.4–16.7 | 9.8–17.4 | 15.4 | 18.2–20.0 |
| HW | 47.7–54.7 | 47.5–51.3 | 49.3 | 47.6–54.1 |
| Na | 7.7–8.3 | 6.8–8.2 | 7.8 | 8.5–9.1 |
| OD | 22.2–24.5 | 22.8–24.2 | 25.3 | 17.5–24.9 |
| IOW | 29.8–38.0 | 31.1–35.7 | 38.7 | 31.1–39.1 |
| UJW | 16.2–19.5 | 15.0–17.8 | 17.2 | 15.5–21.5 |
| LJW | 15.6–17.9 | 15.9–18.6 | 18.8 | 14.2–20.8 |
| Meristic | ||||
| Dorsal‐fin rays | 22–24 | 19–21 | 18 | 19–23 |
| Anal‐fin rays | 27–28 | 25 | 25 | 25–27 |
| Vertebrae | 38 | 38–40 | 37 | 37–39 |
| Upper‐jaw teeth | 9–10 | 8–9 | 10 | 9–10 |
| Lateral‐line scales | 49–54 | 47–54 | 47 | 42–52 |
| Caudal peduncle scales | 11–14 | 12–16 | 13 | 11–12 |
| Morphological group | Thick‐tailed group | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species |
|
|
|
| ||||||||||
| Syntype ( | Syntype ( | Holotype | Paratype ( | All | Holotype | Paratype ( | All | |||||||
| Min | Max | Mean | Min | Max | Mean | Min | Max | Mean | Min | Max | Mean | |||
| Standard length | 50.2 | 54.0 | 52.1 | 58.3 | 62.5 | 60.4 | 39.5 | 36.3 | 39.2 | 38.0 | 63.9 | 57.9 | 66.6 | 63.0 |
| Measurements in % SL | ||||||||||||||
| Total length | 115.4 | 117.0 | 116.2 | 112.6 | 113.9 | 113.2 | 123.6 | 115.6 | 119.6 | 118.8 | 113.1 | 111.4 | 118.1 | 113.7 |
| Body depth | 29.1 | 32.1 | 30.6 | 29.5 | 31.2 | 30.4 | 31.2 | 30.7 | 32.1 | 31.2 | 24.3 | 23.7 | 27.0 | 25.1 |
| Pre‐dorsal length | 67.4 | 67.5 | 67.5 | 66.0 | 66.1 | 66.0 | 65.3 | 65.0 | 66.4 | 65.4 | 64.1 | 61.2 | 65.5 | 63.4 |
| Pre‐anal length | 62.3 | 64.5 | 63.4 | 60.1 | 62.4 | 61.2 | 65.7 | 62.2 | 65.8 | 64.4 | 58.0 | 56.6 | 59.7 | 57.9 |
| Pre‐pectoral length | 27.2 | 28.4 | 27.8 | 25.9 | 26.1 | 26.0 | 29.0 | 28.0 | 29.8 | 29.0 | 25.0 | 24.3 | 26.4 | 25.4 |
| Pre‐pelvic length | 42.6 | 45.1 | 43.8 | 41.4 | 42.2 | 41.8 | 43.7 | 43.7 | 45.7 | 44.5 | 40.5 | 38.9 | 41.2 | 40.0 |
| Length of dorsal fin | 19.1 | 20.4 | 19.8 | 18.3 | 19.5 | 18.9 | 22.1 | 19.8 | 22.0 | 21.3 | 18.4 | 18.3 | 20.1 | 18.9 |
| Length of anal fin | 24.3 | 24.4 | 24.4 | 23.4 | 25.3 | 24.3 | 27.1 | 25.0 | 25.8 | 25.9 | 24.5 | 23.6 | 26.5 | 24.5 |
| Length of pectoral fin | 20.5 | 22.9 | 21.7 | 20.7 | 21.0 | 20.8 | 26.1 | 23.2 | 25.4 | 24.9 | 22.2 | 19.7 | 23.5 | 21.5 |
| Length of pelvic fin | 11.1 | 11.3 | 11.2 | 9.7 | 9.7 | 9.7 | 10.9 | 9.5 | 11.1 | 10.6 | 10.8 | 9.5 | 11.1 | 10.6 |
| Posterodorsal distance | 35.7 | 38.3 | 37.0 | 38.1 | 38.8 | 38.5 | 42.0 | 38.9 | 41.0 | 40.4 | 39.9 | 39.8 | 43.7 | 41.2 |
| Pectoral–pelvic distance | 16.2 | 21.4 | 18.8 | 17.1 | 19.1 | 18.1 | 17.0 | 17.9 | 19.6 | 18.2 | 15.7 | 15.3 | 17.0 | 15.9 |
| Caudal peduncle length | 15.2 | 15.2 | 15.2 | 16.4 | 16.9 | 16.7 | 18.1 | 16.2 | 17.9 | 17.5 | 20.5 | 17.2 | 19.9 | 19.0 |
| Caudal peduncle depth | 7.1 | 8.0 | 7.5 | 6.8 | 7.4 | 7.1 | 6.8 | 7.0 | 7.2 | 7.0 | 7.3 | 6.6 | 7.4 | 6.9 |
| Belly length | 18.7 | 20.9 | 19.8 | 19.6 | 20.7 | 20.1 | 19.4 | 18.4 | 19.0 | 18.9 | 17.9 | 17.0 | 19.1 | 18.1 |
| Head length | 24.6 | 24.9 | 24.7 | 22.4 | 23.1 | 22.7 | 24.6 | 23.9 | 26.4 | 25.3 | 22.5 | 21.5 | 23.3 | 22.3 |
| Measurements in % HL | ||||||||||||||
| Head width | 58.5 | 60.2 | 59.4 | 53.8 | 54.1 | 54.0 | 43.4 | 51.2 | 52.4 | 49.7 | 58.6 | 58.4 | 61.4 | 59.8 |
| Snout–posterior side of eye | 44.3 | 44.7 | 44.5 | 42.3 | 46.1 | 44.2 | 48.5 | 42.1 | 46.5 | 45.3 | 39.5 | 41.4 | 45.8 | 42.9 |
| Snout–center of eye | 34.4 | 38.0 | 36.2 | 33.0 | 36.3 | 34.6 | 34.8 | 28.3 | 33.2 | 31.7 | 29.0 | 31.9 | 36.3 | 33.6 |
| Internasal length | 9.7 | 10.9 | 10.3 | 10.1 | 10.1 | 10.1 | 11.1 | 8.9 | 9.6 | 9.7 | 8.3 | 8.6 | 10.2 | 9.3 |
| Eye diameter | 18.8 | 19.0 | 18.9 | 18.6 | 19.9 | 19.2 | 25.1 | 22.1 | 27.3 | 24.6 | 18.0 | 17.7 | 21.9 | 19.8 |
| Lower‐jaw width | 23.5 | 25.7 | 24.6 | 21.7 | 22.1 | 21.9 | 14.2 | 15.5 | 17.3 | 15.9 | 18.8 | 14.6 | 18.0 | 16.6 |
| Upper‐jaw width | 19.1 | 22.2 | 20.7 | 18.0 | 19.4 | 18.7 | 14.2 | 13.9 | 15.6 | 14.5 | 18.2 | 16.8 | 19.8 | 18.2 |
| Snout length | 14.3 | 15.1 | 14.7 | 11.9 | 13.4 | 12.7 | 13.7 | 12.1 | 13.3 | 13.1 | 14.7 | 12.9 | 15.8 | 14.0 |
| Interorbital width | 39.2 | 44.6 | 41.9 | 38.1 | 41.5 | 39.8 | 39.5 | 33.9 | 43.5 | 39.7 | 32.2 | 32.5 | 40.5 | 36.8 |
| Postorbital length | 61.3 | 63.5 | 62.4 | 60.3 | 63.0 | 61.6 | 56.6 | 55.8 | 58.7 | 56.8 | 62.8 | 53.3 | 62.1 | 60.5 |
| Measurements (other %) | ||||||||||||||
| Caudal peduncle depth (% BD) | 22.2 | 27.4 | 24.8 | 22.9 | 23.6 | 23.2 | 21.7 | 22.0 | 23.3 | 22.4 | 30.1 | 24.4 | 29.3 | 27.5 |
| Caudal peduncle depth (% CPL) | 46.8 | 52.5 | 49.7 | 41.2 | 43.5 | 42.4 | 37.3 | 39.4 | 40.0 | 38.9 | 35.8 | 33.4 | 39.6 | 36.4 |
| Anal‐fin length (% LD) | 119.2 | 127.7 | 123.5 | 127.7 | 130.0 | 128.9 | 123.1 | 116.7 | 130.3 | 121.7 | 133.4 | 121.7 | 140.1 | 129.6 |
| Morphological group | Thick‐tailed group | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Species |
|
|
|
| ||||||
| Syntype ( | Syntype ( | Holotype | Paratype ( | Holotype | Paratype ( | |||||
| Min | Max | Min | Max | Min | Max | Min | Max | |||
| Dorsal‐fin rays | 18 | 18 | 16 | 17 | 19 | 18 | 18 | 16 | 15 | 17 |
| Anal‐fin rays | 23 | 25 | 22 | 22 | 22 | 22 | 23 | 23 | 22 | 23 |
| Pectoral‐fin rays | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 9 | 9 | 10 |
| Pelvic‐fin rays | 6 | 6 | 6 | 6 | 6 | 6 | 6 | 6 | 6 | 9 |
| Vertebra | 39 | 39 | 38 | 38 | 38 | 37 | 38 | 39 | 38 | 39 |
| Teeth in upper jaw | 7 | 8 | 7 | 7 | 7 | 6 | 7 | 7 | 5 | 8 |
| Teeth in lower jaw | 7 | 7 | 8 | 9 | 8 | 7 | 8 | 8 | 6 | 8 |
| Lateral‐line scales | 40 | 46 | 47 | 47 | 52 | 43 | 51 | 48 | 48 | 54 |
| Circumpeduncular scales | 11 | 13 | 12 | 13 | 16 | 15 | 17 | 16 | 14 | 17 |
| Scales lateral line–anal‐fin origin | 9 | 11 | 10 | 11 | 10 | 11 | 13 | 10 | 10 | 12 |
| Scales lateral line–dorsal‐fin origin | 8 | 9 | 10 | 10 | 10 | 10 | 12 | 9 or 10 | 9 | 11 |
| Scales lateral line–pelvic‐fin origin | 10 | 10 | 13 | 13 | 12 | 14 | 14 | 10 | 11 | 13 |
| IP | Thick‐tailed morphological group | Slender‐tailed morphological group | Other morphological groups | Total |
|---|---|---|---|---|
| Nilo‐Sudan |
|
|
| 2 or 3 |
| Upper Guinea |
| 0 or 1 | ||
| Lower Guinea |
|
|
| 2 or 3 |
| Congo |
|
|
| 11 |
| Zambezi |
| 3 | ||
| East Coast |
| 1 | ||
| Great Lakes |
| 1 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFish biology, ecology, and behavior · Fish Biology and Ecology Studies · Fish Ecology and Management Studies
INTRODUCTION
1
The Mormyridae family, with 227 species distributed across 22 genera (Fricke et al., 2024), is a highly diverse freshwater fish endemic to Africa. These fish are well known for their unique appearance with a forked caudal fin and sometimes a protrusion on the head (elongated snout, chin swelling, or appendage), and for being weakly electric. Nevertheless, the species and generic taxonomy of this family remain understudied. This is illustrated by the regular description of new species (e.g., Petrocephalus petersi Kramer et al., 2012; Pollimyrus cuandoensis Kramer et al., 2013; Marcusenius kaninginii Kisekelwa et al., 2016; Paramormyrops ntotom Rich et al., 2017; Marcusenius wamuinii Decru et al., 2019; Cyphomyrus lufirae Mukweze Mulelenu et al., 2019; and Marcusenius verheyenorum Mambo et al., 2019) and even a new genus (Cryptomyrus Sullivan et al., 2016) in the scientific literature.
With the rise of the study of their electrical signals, that is, electric organ discharges (EODs), and genetics, it is possible to examine these fish in new, more integrative, ways. These new techniques also allowed for the discovery of new species that, based solely on the use of morphological characteristics, would have remained unnoticed (Feulner et al., 2006; Hopkins, 1999; Rich et al., 2017). However, morphological analyses continue to be an important tool to assess species diversity in fishes, especially when living fish and adequately preserved tissue material are unavailable for behavioral and molecular approaches. One of the mormyrid genera for which both its delimitation, in terms of included species, and species differentiation of included species have largely been neglected is Pollimyrus Taverne, 1971. This lack of knowledge hinders a better understanding of their hydro‐geographic distribution and thus also their conservation. Therefore, a synthesis of the morphological description of all species of the genus Pollimyrus is provided, which until now was lacking in the scientific literature.
The genus Pollimyrus was established by Taverne in 1971. He identified Mormyrus isidori Valenciennes, 1847, originally described from the “Nile” without further specifications, as its type species. Further, 24 valid species were added to the new genus by Taverne (1971a). These had previously been assigned to three other genera: Marcusenius Gill, 1862, Mormyrus Linnaeus, 1758, and Petrocephalus Marcusen, 1854 (Taverne, 1971a) (Table 1). The following year, Taverne (1972) rediagnosed the genus Pollimyrus on an osteological basis and reallocated some species to the genera Petrocephalus and Heteromormyrus Steindachner, 1866, the latter a former subgenus of Mormyrus, which he elevated to genus level (Table 1). During the decades following Taverne's revisions (1971a, 1971b, 1972), there have been quite some generic reassignments, with species being transferred between Pollimyrus and other genera and new species being described (e.g., Bigorne, 1990; Bigorne, 2003; Hopkins et al., 2007; Kramer et al., 2012; Kramer et al., 2013; Kramer & Van der Bank, 2011; Lavoué et al., 2010; Lévêque et al., 1991; Levin & Golubtsov, 2018; Rich et al., 2017; Skelton, 2019; Stiassny et al., 2021; Sullivan et al., 2022; Teugels & Hopkins, 1998) (Table 1). Nevertheless, some questions remain regarding the generic assignment of certain species, such as Petrocephalus guttatus Fowler, 1936, that might belong to Pollimyrus (Lavoué et al., 2010). To date, 18 valid species are recognized within the genus (Fricke et al., 2024; see Table 1). Taverne (1971a, 1971b, 1972) diagnosed the genus Pollimyrus osteologically as species with a small lateral ethmoid bone, a large and curved mesethmoid bone, six circumorbital bones with the antorbital and first infraorbital not fused, and four hypural bones (i.e., fused ventral hypurals as defined by Teugels & Hopkins, 1998). Further, with regard to the external morphology, the species of the genus have a short or elongated body. Their snout is shorter than the postorbital part of the head. The mouth is terminal, inferior, or subinferior. The caudal peduncle depth fits two to five times in the peduncle length. Meristically, they have 15–36 dorsal‐fin rays, 21–31 anal‐fin rays, 9–12 pectoral‐fin rays, 35–70 lateral‐line scales, 7–21/8–23 scales in transverse line, 8–17/6–20 scales in transverse line between the dorsal‐ and anal‐fin origins, 12–20 scales around the caudal peduncle, 5–9/6–10 bicuspid teeth on the oral jaws, and 39–45 vertebrae (Taverne, 1971a, 1971b, 1972). Besides these osteological and meristic diagnostic characteristics, these species can further be recognized morphologically based on the combination of the following, additional, qualitative characteristics: rounded head in lateral view; no mental lobe; nostrils well separated and posterior nostril close to anterior rim of eye; and dorsal‐ and anal‐fin origins at the same level (Bigorne, 2003; Hopkins et al., 2007).
Identifying collected specimens from the Luki (Democratic Republic of the Congo), Kouilou‐Niari (Republic of the Congo), Buzi (Mozambique), and the Lugenda (Mozambique) rivers was hindered by the lack of an updated alpha‐taxonomical synthesis of the genus since its original description by Taverne (1971a, 1971b, 1972), which goes beyond mere listing of species. Further, it soon became clear that there were also other taxonomical problems within this genus, at least, for the Congo basin (see also, e.g., Hopkins et al., 2007). Therefore, in this study, a taxonomic overview of the species of Pollimyrus is provided based on a detailed morphometric re‐examination of the type specimens of all nominal species; taxonomical issues encountered during this re‐examination are discussed and, based on this new synthesis, four potential new species for science are analysed: Pollimyrus sp. “luki,” Pollimyrus sp. “kouilou‐niari,” Pollimyrus sp. “buzi,” and Pollimyrus sp. “lugenda.” Finally, the potential implications of the geographical distribution of the genus and its constituting species on their conservation are also discussed.
MATERIALS AND METHODS
2
Taxa and specimens examined
2.1
To provide a morphological synthesis, the type specimens of all but one (i.e., Pollimyrus macroterops; see later) currently valid nominal species placed in the genus Pollimyrus were studied. Only type specimens were selected, except for a few specific cases (see later), to provide a morphometric delineation of each of the type series as an initial taxonomic basis for the genus. Non‐type specimens have been identified in the past based on resemblance to types and/or to descriptions of species or on geographical proximity to the type location. However, these could potentially represent other species than the one they are currently assigned. This is largely due to a lack of an updated taxonomic overview of the species in the genus (personal observation; Hopkins et al., 2007). Therefore, these were generally not included in this study.
Some species, currently assigned to the genera Cyphomyrus Myers, 1960, and Petrocephalus Marcusen, 1854, were also included because, according to literature, there has been some discussion about their generic placement (P. guttatus, Lavoué et al., 2010; Cyphomyrus plagiostoma, Stiassny et al., 2021; Peterson et al., 2022). Due to the confusion in the past concerning the validity of Pollimyrus isidori fasciaticeps as a distinct species from P. isidori and Pollimyrus osborni (Konan et al., 2013), the holotype of this species was also included in the study. Additionally, the type specimens of the subspecies and junior synonyms of Pollimyrus stappersii and Pollimyrus tumifrons, respectively, were included, as they were available at Royal Museum for Central Africa (RMCA), where the study was performed. Pollimyrus macroterops was not included in this study, as this species was not considered a (potential) Pollimyrus until recently (Sullivan et al., 2022), which was after the data collection phase for the present study.
In total, 140 specimens, including 125 type specimens of 25 nominal species, from collections of the American Museum of Natural History, New York (AMNH), Academy of Natural Sciences of Drexel University, Philadelphia (ANSP), Muséum National d'Histoire Naturelle, Paris (MNHN), Natural History Museum, London (NHM), Royal Museum for Central Africa, Tervuren (RMCA), South African Institute for Aquatic Biodiversity, Makhanda (SAIAB), and Zoologische Staatssammlung München, München (ZSM) were examined (Table 2). The four species that are new to science are described in detail. The previously described nominal species are only discussed within the scope of genus and species delineations.
Morphology
2.2
In total, 27 measurements, 12 meristics (including the number of vertebrae obtained through X‐rays, using a radiograph cabin VisiX–MedexLoncin SA 2011 [www.medex.be] with a DeReO WA detector and a GemX‐160 generator, housed at the RMCA) and 9 qualitative observations on the body, including fin shape, head shape, and dentition, were carried out. The measurements and meristics taken follow Boden et al. (1997) and Kramer et al. (2013) (Table 3). Two additional measurements and qualitative characteristics were also included (Table 3). Abbreviations and short explanations of these measurements and meristics are given in Table 3. A visual overview of the measurements and some of the qualitative features is given in Figure 1. Specimens were not sexed, as potential differences between males and females were hard to see using the shape of the anal fin (e.g., Greisman & Moller, 2005), and it is not clear if the studied species show the same sexual dimorphism as found in other mormyrid species.
Methodology of measurements taken and qualitative observations made on Pollimyrus specimens. (a) Measurements (see Table 3 for abbreviations and further explanation) (based on Boden et al., 1997; Figure 1). (b) Shape of head: a blocky snout with the angle between the anterior most/vertical side of the snout and the dorsal part of the head positioned above the level of the eye (indicated by a dotted line) with mental lobe or a rounded snout with the angle at the level of the eye without mental lobe. (c) Pectoral‐fin shape: rounded with convex diminution of the shorter rays or pointed with concave diminution of shorter rays. (d) Dorsal‐ and anal‐fin shape: rounded with a rather straight overall edge or pointed with concave edge. (e) Dentition type: type 1 dentition, with one tooth occupying the most anterior and middle position, or type 2 dentition, with three teeth on one line occupying the most anterior and middle position together.
Analysis
2.3
Principal component analysis (PCA) was performed in Past (version 3.16; Hammer et al., 2001) to explore the multivariate dataset. Measurements were log transformed, and the covariance matrix used. Total length was excluded. When using log‐transformed measurements, the individual loads of all variables on the first principal component (PC 1) are of the same magnitude and sign, and PC 1 can therefore be regarded as a proxy for multivariate size (Jolicoeur, 1963; Snoeks, 2004; Van Steenberge et al., 2015; also see Figure S21 in the Supplementary file S1). The correlation matrix was used for the raw meristic data.
Based on the measurements, meristic, and qualitative observations collected, it was first verified whether the studied specimen of each species should be assigned to the genus Pollimyrus using its present diagnosis (following Bigorne, 2003; Hopkins, 2007; Taverne, 1971a, 1971b; Taverne, 1972) or, instead, might need reassignment to another genus. Subsequently, specimens and species within the genus Pollimyrus were classified into morphological groups. Every specimen and species was then compared to all other nominal species of the same group to re‐evaluate the validity of the individuals composing nominal species and/or identify it as a potential new species for science. A map with all the localities of the studied specimens per species is provided in Figure 2.
Map showing catch locations of the specimens studied of all nominal Pollimyrus species retained or reallocated to the genus and the four species identified as new to science, divided by ichthyogeographical province (IP; colored areas) following Lévêque and Paugy (2017). The precise type locality for Pollimyrus isidori and Pollimyrus adspersus is unclear.
DNA analysis was not included in this study as it intended to work only on the type series of the nominal valid species. Destructive sampling would have been too impactful for the preservation of these specimens, many of which have been collected over a century ago. Although DNA analyses on preserved mormyrid specimens have become more successful recently (e.g., Sullivan et al., 2022), this technique was not proven to be as successful during the time of the data collection of this study (2019). There was no guarantee that DNA data could be obtained for each species, making an in‐depth comparison not possible.
ETHICAL STATEMENT
2.4
The specimens from the Buzi River system in the Manica Province, Mozambique, were collected with a Samus electro‐fisher during a survey of the Chimanimani Transfrontier Conservation Area funded by the Transfrontier Conservation Areas and Institutional Strengthening Project based in Maputo. Specimens were killed by overdosing in clove oil and fixed in 10% formalin immediately after. The preserved specimens were sent to SAIAB (Makhanda, South Africa).
The specimens from the Rovuma River system in the Niassa Province, Mozambique, were collected with a 5 m seine net during a biodiversity survey for the Sociedade para a Gestão e Desenvolvimento da Reserva do Niassa (SRN) who manages the Niassa Reserve. Specimens were killed by overdosing in clove oil and immediately after were fixed in 10% formalin. The preserved specimens were sent to SAIAB (Makhanda, South Africa).
The specimens from the Luki Basin were collected using gill nets and traps as part of the MbiSa‐Congo project (2013–2019) by collaborators of the Institut Supérieur Pédagogique de Mbanza‐Ngungu. Specimens were also killed by overdosing in clove oil and fixed in 10% formalin immediately after. The collection missions were approved by the political‐administrative authorities of the province of Kongo Central (DRC). The preserved specimens were sent to the RMCA (Tervuren, Belgium).
RESULTS
3
Generic (re)assignments
3.1
In this section, the generic assignments for several species are investigated as these are still under debate or unresolved. These species have been the topic of recent assignments to other genera (Cyphomyrus plagiostoma; Stiassny et al., 2021; Peterson et al., 2022) or have been suggested to belong to Pollimyrus (Petrocephalus guttatus; Lavoué et al., 2010). Additionally, the generic status of Pollimyrus eburneensis is analysed due to its clear morphological differentiation from other Pollimyrus and specifically its type species P. isidori.
Pollimyrus eburneensis Bigorne, 1990 (Figure S1) was originally described from the Agnébi basin in Ivory Coast in a stream near the village Attienguié (Daget & de Rham, 1970) as a valid subspecies of Pollimyrus kingsleyae Günther, 1896 (now Paramormyrops kingsleyae). Despite P. kingsleyae having been reassigned to Brienomyrus Taverne, 1971 (Teugels & Hopkins, 1998) and later Paramormyrops Taverne, Thys van den Audenaerde and Heymer, 1977 (Hopkins et al., 2007), P. eburneensis was only recently elevated to species level but remained assigned to the genus Pollimyrus (Rich et al., 2017). However, P. eburneensis is clearly distinct from all other Pollimyrus species by having a more elongated body (body depth [BD] 18.7%–21.3% standard length [SL] vs. 22.2%–35.7% SL; 31.9% SL in P. isidori; 22.5%–25.7% SL in Pollimyrus petricolus, another elongated species found in West Africa), a deeper caudal peduncle (32.7%–33.6% BD vs. 12.8%–31.0% BD; 23.3% BD in P. isidori; 25.4%–28.8% BD in P. petricolus), and a, usually, smaller eye diameter (10.5%–14.6% head length [HL] vs. 13.7%–28.7% HL; 22.9% HL in P. isidori; 21.6%–23.7% HL in P. petricolus). Further, the ventral hypurals are visible and unfused on the X‐rays of the largest specimens (all those >53 mm SL), including the holotype of P. eburneensis, whereas fused is diagnostic for the genus Pollimyrus according to Taverne (1971a, 1971b). A sharp X‐ray photograph could not be obtained for the smallest of the P. eburneensis paratypes. In conclusion, this species is not considered a valid member of the genus Pollimyrus (see Discussion), but its characteristics currently match best with those of the genus Paramormyrops (see Sullivan et al., 2016).
Petrocephalus guttatus Fowler, 1936 (Figure S2) was originally described from near Kribi in Cameroon. This species has been reported to be morphologically more similar to the species of the genus Pollimyrus rather than Petrocephalus Marcusen, 1854 (Lavoué et al., 2010). Observations of the type specimens of this species show that its nostrils are placed further apart from each other than the posterior nostril is to the eye, as is typical in P. isidori and other members of the genus Pollimyrus, whereas in Petrocephalus the nostrils are positioned closer together than one is to the eye (see Figures S1–S23). The species has 7–8 teeth in the lower jaw (Table 6), whereas most Petrocephalus species have 14–37, including 22–29 in Petrocephalus bane (Lacepède, 1803), the type species of the genus (Kramer & van der Bank, 2000; Lavoué et al., 2004; Lavoué et al., 2010; Stiassny et al., 2007). Furthermore, the mouth is placed clearly anteriorly to the level of the eye (Figure S5), similar as in Pollimyrus, and all meristics (Table 7) fall within the ranges known for the other species of the genus Pollimyrus. Because there is no further resemblance to Petrocephalus, the species is reassigned to Pollimyrus (see Discussion) and is further referred to as Pollimyrus guttatus in this study.
Cyphomyrus plagiostoma (Boulenger, 1898) (Figure S3) was originally described from the Lower Congo River near Matadi (DRC). Although previously being placed in Pollimyrus (Taverne, 1971a, 1971b), the species has recently been reassigned to Cyphomyrus based on morphological evidence and tentative unpublished genetic evidence (Stiassny et al., 2021). A more recent study, however, placed the species in the genus Pollimyrus based on genetic analysis (Peterson et al., 2022). A morphological comparison of this species with the two genera is considered here. C. plagiostoma shares some characteristics with P. isidori and other species assigned to the genus Pollimyrus, whereas in others it more closely resembles Cyphomyrus spp., such as Cyphomyrus petherici (Figure S4). Its dorsal fin has more fin rays than in Pollimyrus species (32–33 vs. 14–28; 20 in P. isidori; 30–37 in Cyphomyrus psittacus, type species of Cyphomyrus), and its anal fin is shorter than the dorsal fin (73.3%–76.9% dorsal fin length (LD) vs. 90.0%–233.1% LD; 119.4% LD in P. isidori; 60.7%–63.6% LD in C. petherici), thus being more similar to the members of the genus Cyphomyrus. The dorsal‐fin origin, however, is positioned only slightly anterior to the anal‐fin origin like in P. isidori and other Pollimyrus species. Cyphomyrus plagiostoma has 9 teeth in the upper jaw and 8–10 in the lower jaw, whereas Pollimyrus species have 7–10 teeth in the upper jaw and 8–11 teeth in the lower jaw. The tail complex of C. plagiostoma also has fused ventral hypurals, which is diagnostic for the genus Pollimyrus following Taverne (1971a, 1971b). Further, the caudal peduncle is very thin relative to the BD compared to other Cyphomyrus species (13.3%–13.9% BD in C. plagiostoma vs. 22.7%–23.7% BD in C. petherici, although this falls within the ranges found in members of the genus Pollimyrus, 12.7%–31.2% BD; 23.3% BD in P. isidori). Due to this overall intermediate morphological state, its current generic position is inconclusive and awaits further data. For reasons of nomenclatorial stability, however, this species is further referred to as Cyphomyrus plagiostoma in this study, as this is the currently valid status (Fricke et al., 2024; see also Discussion).
Pollimyrus guttatus and C. plagiostoma are further included in this study and compared to the other species assigned to Pollimyrus. Paramormyrops eburneensis is not further analysed.
The delineation of morphological groups, species complexes, and species within the genus Pollimyrus
3.2
Between the type specimens of Pollimyrus, five morphological groups can be differentiated based on the caudal peduncle depth and several qualitative characteristics, that is, snout shape (Figure 1b), the presence or absence of a mental lobe (Figure 1b), the shape of the pectoral fins (Figure 1c), and dorsal and anal fins (Figure 1d) (Table 4; Figure 3). Meristic data did not show differentiation between these groups. Each morphological group is named after the main characteristic defining it, and for the smaller groups, the main taxon in it.
Differentiation of morphological groups. (a) Caudal peduncle depth (CPD) (% BD) against standard length (SL) (mm) for all Pollimyrus specimens examined (n = 140) (see Table 4); a differentiation can be seen between 0.20 and 0.21 CPD/BD (dotted line). (b) Scatterplot of second principal component (PC 2) against PC 3 for a principal component analysis (PCA), with explained variance between brackets, on 26 log‐transformed measurements for all Pollimyrus specimens examined (n = 140) (see Table S1 for the loadings and Figure S21 for a plot of PC 1 against SL).
One can divide those Pollimyrus species with a thick caudal peduncle (21.7%–31.2% BD) from those with a slender caudal peduncle (12.7%–20.2% BD) (Figure 3a). The caudal peduncle depth (% BD) does not show any allometric size effect (Figure 3a). A PCA on 26 log‐transformed measurements showed that specimens with thick and slender caudal peduncle are somewhat separated by a combination of PC 2 and PC 3 (Figure 3b). The most important loadings on PC 2 are for the caudal peduncle depth, eye diameter, anal‐fin length, lower‐jaw width, and dorsal‐fin length, and on PC 3 for the caudal peduncle depth, caudal peduncle length, snout–anterior nostril length, interorbital width, and lower‐jaw width (see Table S1), verifying the separation of Pollimyrus on the caudal peduncle depth (% BD) into two large groups, which are further divided into morphological groups using qualitative characteristics (Table 4). PC 1 has a strong correlation with the SL (Figure S21).
Because these five morphological groups are clearly distinct from each other, these will further be analysed separately in the sections below: (1) the thick‐tailed group, (2) the P. isidori group, (3) the Pollimyrus sp. “kouilou‐niari” group, (4) the slender‐tailed group, and (5) the P. tumifrons group. Per morphological group, the differentiating criteria and the taxonomic status of each species are provided and discussed. The order in which the species are listed and discussed is determined by the ease to distinguish each of them from the other species in the group. Those species already differentiated have been left out when providing the differentiating character set for the following species.
Thick‐tailed group
3.2.1
A PCA on 26 log‐transformed measurements shows a separation of P. guttatus and P. petricolus on a combination of PC 2, with the most important loadings being the lower‐jaw width, eye diameter, and the length of the dorsal fin, and PC 3, with the most important loadings being the caudal peduncle length, eye diameter, and the lower‐jaw width (Figure 4; Table S2 for the loadings). A PCA on the meristics shows a separation of P. petricolus on PC 1, with the most important loadings being the number of lateral‐line scales, dorsal‐fin rays, and vertebrae, and of P. brevis on PC 2, with the most important loadings being the number of anal‐fin rays, teeth in the upper jaw, and caudal peduncle scales (Figure 4c,d and Table S3 for the loadings).
Plots of morphometric data of thick‐tailed Pollimyrus. (a) First principal component (PC 1) (proxy for standard length [SL]) against PC 2 for a principal component analysis (PCA) on 26 log‐transformed measurements (n = 41). (b) PC 2 against PC 3 for a PCA on 26 log‐transformed measurements (n = 41). (c) PC 1 against PC 2 for a PCA on 12 meristics (n = 41). (d) PC 2 against PC 3 for a PCA on 12 meristics (n = 41) (also see Tables 6 and 7, S2 and S3). Stars indicate holotypes and syntypes, and circles indicate paratypes. Explained variance is noted between brackets for each PC axis.
Pollimyrus petricolus (Daget, 1954) (Figure S5) can be distinguished on meristics from all other thick‐tailed Pollimyrus species by its higher number of lateral‐line scales (58–67 vs. 40–54), usually higher number of dorsal‐fin rays (19–22 vs. 15–19), and, usually a higher number of vertebrae (41–43 vs. 36–41). The dorsal‐fin length is nearly equal (89.2%–112.2% BD) to the body depth in P. petricolus, whereas it is less than 82.0% (i.e., 82.0% BD for one paratype of Pollimyrus marianne being the closest to this range) for the other thick‐tailed Pollimyrus. Further, it has a shallower body depth compared to four other thick‐tailed species, P. brevis, P. castelnaui, P. guttatus, and P. sp. “buzi” (22.2–25.7% SL vs. 29.1%–32.1% SL). It differs from P. cuandoensis and P. sp. “lugenda” by having a longer dorsal fin (21.5%–25.0% SL vs. 16.1%–18.9% SL and 17.3%–17.5% SL, respectively). Finally, P. petricolus differs from P. stappersii by having more circumpeduncular scales (16–20 vs. 13) and from P. marianne by having more dorsal–lateral scales (13–14 vs. 9–11).
Pollimyrus guttatus (Fowler, 1936) (Figure S2) differs from P. petricolus by the aforementioned characteristics (see above). Pollimyrus guttatus differs from all other thick‐tailed species, except P. cuandoensis and P. marianne, by its larger eye diameter (22.1%–27.3% HL vs. 14.3%–19.4% HL). It differs from P. brevis by having a wider head (43.4%–52.4% HL vs. 58.5%–60.2% HL), a slenderer upper jaw (13.9%–15.6% HL vs. 19.1%–22.2% HL) and lower jaw (14.2%–17.3% HL vs. 23.5%–25.7% HL), a shorter postorbital distance (55.8%–58.7% HL vs. 61.3–63.5% HL), and more circumpeduncular scales (15–17 vs. 11–13). It differs from P. castelnaui by having a longer pectoral fin (23.2%–26.1% SL vs. 20.7%–21.0% SL), slenderer upper jaw (13.9%–15.6% HL vs. 18.0%–19.4% HL) and lower jaw (14.2%–17.3% HL vs. 21.7%–22.1% HL), and more circumpeduncular scales (15–17 vs. 12–13). It can be distinguished from P. cuandoensis by having a longer pectoral fin (23.2%–26.1% SL vs. 19.4–21.9% SL) and more dorsal‐fin rays (18–19 vs. 15–16). It differs from P. stappersii by having longer pectoral fins (23.2%–26.1% SL vs. 19.9%–20.6% SL), a shorter postorbital distance (55.8%–58.7% HL vs. 61.5%–62.3% HL), and more circumpeduncular scales (15–17 vs. 13). It differs from P. sp. “buzi” by having a slenderer upper jaw (13.9%–15.6% HL vs. 22.6%–24.5% HL), a longer anal fin (25.0%–27.1% SL vs. 20.2%–22.5% SL), and a longer pectoral fin (23.2%–26.1% SL vs. 18.1%–19.6% SL). It differs from P. sp. “lugenda” by having a deeper body (30.1%–32.1% SL vs. 25.3%–26.7% SL), a longer pectoral fin (23.2%–26.1% SL vs. 19.2%–20.5% SL), and more dorsal‐fin rays (18–19 vs. 15). Finally, it differs from P. marianne by having a deeper body (30.1%–32.1% SL vs. 23.7%–27.0% SL), a longer pre‐anal distance (62.2%–65.8% SL vs. 56.6%–59.7% SL), longer pre‐ventral distance (43.7%–45.7% SL vs. 38.9%–41.2% SL), and a slenderer head (43.4%–52.4% HL vs. 58.4%–61.4% HL).
Pollimyrus brevis (Boulenger, 1913) (Figure S6) differs from P. marianne by its deeper body (29.1%–32.1% SL vs. 23.7%–27.0% SL), a longer pre‐dorsal distance (67.4%–67.5% SL vs. 61.2%–65.5% SL), a longer pre‐anal distance (62.3%–64.5% SL vs. 56.6%–59.7% SL), a shorter caudal peduncle length (15.2% SL vs. 17.2%–20.5% SL), a wider lower jaw (23.5%–25.7% HL vs. 14.6%–18.8% HL), fewer lateral‐line scales (40–46 vs. 48–54), and by its most anterior teeth being the same size as the posterior teeth, and not bigger as in P. marianne. It can be distinguished from P. castelnaui by its longer pelvic fin (11.1%–11.3% SL vs. 9.7% SL), a wider head (58.5%–60.2% HL vs. 53.8%–54.1% HL), and fewer scales between the pelvic fin and the lateral line (10 vs. 13). P. brevis differs from P. cuandoensis by having a shorter caudal peduncle length (15.2% SL vs. 16.4–16.9% SL) and more dorsal‐fin rays (18 vs. 15–16). It differs from P. stappersii (including P. stappersii kapangae) by having a wider head (58.5%–60.2% HL vs. 52.7%–54.0% HL), a wider lower jaw (23.5%–25.7% HL vs. 17.9%–18.3% HL), and fewer lateral‐line scales (40–46 vs. 48–53). It can be distinguished from P. sp. “buzi” by having a longer anal fin (24.3%–24.4% SL vs. 20.2%–22.5% SL), a wider lower jaw (23.5%–25.7% HL vs. 17.1%–19.1% HL), and fewer lower‐jaw teeth (seven vs. nine). It differs from P. sp. “lugenda” by having a deeper body (29.1%–32.1% SL vs. 25.3%–26.7% SL), a longer dorsal fin (19.1%–20.4% SL vs. 17.3%–17.5% SL), a wider head (58.5%–60.2% HL vs. 52.7%–53.8% HL), a larger eye (18.8%–19.0% HL vs. 14.3%–15.5% HL), a wider lower jaw (23.5%–25.7% HL vs. 14.7%–16.2% HL), a wider interorbital width (39.2%–44.6% HL vs. 31.7%–33.1% HL), and more dorsal‐fin rays (18 vs. 15).
Pollimyrus castelnaui (Boulenger, 1911) (Figure S7) differs from P. marianne by having a deeper body (29.5%–31.2% SL vs. 23.7%–27.0% SL), a slenderer head (53.8%–54.1% SL vs. 58.4%–61.4% SL), a wider lower jaw (21.7%–22.1% HL vs. 14.6%–18.8% HL), a pelvic fin that is less than half the length of the belly (vs. slightly longer than half the belly length), and by its most anterior teeth being the same size as the posterior teeth (vs. anterior teeth being a bit larger). It can also be distinguished from P. cuandoensis by having a pelvic fin that is less than half the length of the belly. It can be distinguished from P. stappersii/kapangae by having a wider lower jaw (21.7%–22.1% HL vs. 17.9%–18.3% HL), having fewer scales between the lateral‐line and the anal‐fin origin (10–11 vs. 12–13), and by their geographical isolation. Pollimyrus castelnaui differs from P. sp. “buzi” by having a slenderer upper jaw (18.0%–19.4% HL vs. 22.6%–24.5% HL), a pelvic fin that is less than half the length of the belly (vs. slightly longer than half the belly length), and in its geographical occurrence. It differs from P. sp. “lugenda” by having a deeper body (29.5%–31.2% SL vs. 25.3%–26.7% SL), a longer snout–eye distance (42.3%–46.1% HL vs. 39.4%–39.7% HL), a larger eye (18.6%–19.9% HL vs. 14.3%–15.5% HL), a wider lower jaw (21.7%–22.1% HL vs. 14.7%–16.2% HL), a wider interorbital width (38.1%–41.5% HL vs. 31.7%–33.1% HL), more scales between the pelvic fin and the lateral line (13 vs. 11), and by having a pelvic fin that is less than half the length of the belly (vs. slightly longer than half the belly length).
Specimens found in the Buzi basin in Mozambique, here preliminarily identified as Pollimyrus sp. “buzi,” differ from P. marianne, P. cuandoensis, P. stappersii/kapangae, and P. sp. “lugenda” by having a wider upper jaw (22.6%–24.5% HL vs. 13.3%–20.2% HL). They can further be differentiated from P. marianne by having a deeper body (29.6%–29.9% SL vs. 23.7%–27.0% SL), a shorter post‐dorsal distance (37.6% SL vs. 39.8%–43.7% SL), and its most anterior teeth being the same size as the posterior teeth. They are distinguished from P. stappersii/kapangae by having a shorter belly length (17.5%–18.2% SL vs. 20.8%–21.6% SL). They further differ from P. sp. “lugenda” by having a deeper body (29.6%–29.9% SL vs. 25.3%–26.7% SL), a wider head (55.2%–58.7% HL vs. 52.7%–53.8% HL), a larger eye (17.1%–18.9% HL vs. 14.3%–15.5% HL), a wider lower jaw (22.6%–24.5% HL vs. 19.5%–19.7% HL), and a wider interorbital width (37.2%–40.9% SL vs. 31.7%–33.1% SL). As these specimens are morphologically distinct from all other Pollimyrus, they are proposed as a new species to science: Pollimyrus weyli sp. nov. (Figure 12). The detailed description of this species new to science is provided below.
Specimens found in the Ruvuma basin in Mozambique, here preliminarily identified as Pollimyrus sp. “lugenda,” differ from P. marianne by its shorter post‐dorsal distance (36.8%–37.0% SL vs. 39.8%–43.7% SL), a slenderer head (52.7%–53.8% HL vs. 58.4%–61.4% HL), a smaller eye (14.3%–15.5% HL vs. 17.7%–21.9% HL), and its most anterior teeth being the same size as the posterior teeth. They differ from P. stappersii/kapangae by having a short snout–eye length (39.4%–39.7% HL vs. 44.3%–44.5% HL). There are slight differences between these specimens and P. cuandoensis in eye diameter (14.3%–15.5% HL vs. 15.5%–21.9% HL) and snout–eye length (39.4%–39.7% HL vs. 40.5%–45.3% HL). As these specimens are morphologically distinct from all other Pollimyrus, they are proposed as a new species to science: Pollimyrus krameri sp. nov. (Figure 8). The detailed description of this species new to science is provided below.
Pollimyrus marianne Kramer et al., 2003 (Figure S8) differs from P. stappersii/kapangae by having a wider head (58.4%–61.4% HL vs. 52.7%–54.0% HL) and a shorter pre‐anal distance (56.6%–59.7% SL vs. 61.6%–63.5% SL). It cannot be distinguished from P. cuandoensis (see below).
Pollimyrus cuandoensis Kramer et al., 2013 (Figure S8) differs slightly from P. stappersii/kapangae by having a thicker caudal peduncle (6.6%–8.8% SL vs. 5.9%–6.1% SL), a shorter belly (17.3%–20.1% SL vs. 20.8%–21.6% SL), and more circumpeduncular scales (14–16 vs. 13). It cannot be distinguished from P. marianne. Because P. marianne and P. cuandoensis both occur in the Upper Zambezi basin in adjacent rivers (main Zambezi River and Kwando River, respectively), and the species don't separate on a PCA of 26 log‐transformed measurements (Figure 5; Table S6), it is suggested here to place the two species in synonymy (also see Discussion). Pollimyrus cuandoensis has large ranges for many measurements and meristics (Tables S11 and S12). Despite these large ranges there is no indication that these specimens belong to a different species. All type specimens of P. cuandoensis seem to be conspecific.
Plots of morphometric data of the marianne species complex, including Pollimyrus marianne Kramer et al., 2003, (n = 11) and Pollimyrus cuandoensis Kramer et al., 2013, (n = 12), and comparing with morphological similar yet distinct Pollimyrus castelnaui (Boulenger, 1911) (n = 4) from the Zambezi IP (see also Table S6). (a) First principal component (PC 1) (proxy for standard length [SL]) against PC 2 for a principal component analysis (PCA) on 26 log‐transformed measurements. (b) PC 2 against PC 3 for a PCA on 26 log‐transformed measurements. Stars indicate holotypes or syntypes, circles indicate paratypes, and triangles indicate non‐type specimens. Explained variance is noted between brackets for each PC axis.
Pollimyrus stappersii (Boulenger, 1915) (Figure S9) differs from all other thick‐tailed species by the aforementioned characteristics. It consists of two subspecies, P. s. stappersii and P. s. kapangae. The holotype of P. s. kapangae shows some differences compared to that of P. s. stappersii and other non‐type specimens currently identified to belong to this complex (see Supplementary file S2), such as slenderer teeth, more anal‐fin rays (26 vs. 22), and fewer lateral‐line scales (48 vs. 53; although there remains some uncertainty due to the scales being covered by skin), which in our view are important differences between both (sub)species when considering the difference found between other species within the genus. However, these are insufficient to elevate P. s. kapangae to species level due to the lack of differences in measurements and the present lack of additional topotypic specimens. As there are no non‐type specimens available in museum collections from the same localities as the holotypes of both subspecies, this complex cannot be further analysed. Therefore, for the time being, a single species P. stappersii is recognized here with two valid subspecies, considering the differences found.
P. isidori group
3.2.2
This group consists of three currently valid species: P. isidori (type species of Pollimyrus), P. fasciaticeps, and P. nigricans.
Pollimyrus isidori (Valenciennes, 1847) (Figure S10) differs from both the other species of the group by its higher number of dorsal‐fin rays (20 vs. 14–17 in P. nigricans and 17 in P. fasciaticeps) and more pelvic–anal scales (16 vs. 12 and 14, respectively). It can further be distinguished from both the other species by its deeper body (33.3% SL vs. 23.5%–29.5% SL in P. nigricans and 29.7% SL in P. fasciaticeps). Finally, it has very short pectoral fins that do not reach halfway of the pelvic fin (vs. almost reaching the distal tip of the pelvic fin in P. nigricans and P. fasciaticeps).
Pollimyrus nigricans (Boulenger, 1906) (Figure S11) differs from the other two species, P. isidori and P. fasciaticeps, by having a shorter dorsal fin (16.0%–19.1% SL vs. 22.5% SL and 22.3% SL), a shorter post‐dorsal distance (35.5%–37.5% SL vs. 42.8% SL and 43.5% SL), a larger postorbital distance (62.4%–67.3% SL vs. 57.3% SL and 52.2% HL), a slightly shorter anal fin (21.0%–24.6% SL vs. 26.8% SL and 27.7% SL), and fewer pelvic–anal scales (12 vs. 16 and 14). This species also has longer but not pointed anterior parts of the dorsal and anal fins (vs. pointed in P. fasciaticeps). The pectoral fins almost reach the posterior tip of the pelvic fins (vs. not reaching halfway the pelvic fin in P. isidori).
Pollimyrus fasciaticeps (Boulenger, 1920) (Figure S12) was originally described based on a single specimen. The holotype can be distinguished from the type specimens of the other two species by the longer distance between the snout and the anterior eye margin (50% HL vs. 41.4% HL in P. isidori and 37.5%–42.6% HL in P. nigricans), slightly shorter pre‐dorsal distance (61.6% SL vs. 63.8% SL in P. isidori and 64.5%–67.8% SL in P. nigricans), and having a higher number of dorsal–lateral scales (16 vs. 14 in P. isidori and 11–14 in P. nigricans). It can further be distinguished from P. isidori by the wider head (59.7% HL vs. 50.9% HL). This species also has pointed anterior parts of the dorsal and anal fins (vs. longer but not pointed in P. isidori and P. nigricans). The pectoral fins almost reach the posterior part of the pelvic fins (vs. not reaching halfway the pelvic fin in P. isidori).
Pollimyrus sp. “kouilou‐niari” group
3.2.3
The specimens originating from the Kouilou‐Niari River, a coastal river basin of the Lower Guinea ichthyogeographical province (IP) in the Republic of the Congo, is here preliminarily identified as Pollimyrus sp. “kouilou‐niari.” It can be distinguished from all other Pollimyrus species by having a thick caudal peduncle compared to the slender caudal peduncle of all Pollimyrus species (20.3%–24.2% BD vs. 12.7%–20.2% BD), a blocky snout (vs. usually round), and no mental lobe (vs. small mental lobe) compared to thick‐tailed Pollimyrus, and rounded pectoral fin (vs. pointed) compared to the members of the P. isidori group.
Only one other congeneric species occurs in the same IP, P. guttatus, which occurs near Kribi, in the Kineke River in southern Cameroon. The specimens from the Kouilou‐Niari River can be distinguished from P. guttatus by having a subterminal mouth (vs. terminal mouth), a shallower body depth (22.6%–28.7% SL vs. 30.7%–32.1% SL), a shorter pre‐ventral distance (33.7%–40.2% SL vs. 43.7%–45.7% SL), more dorsal‐fin rays (21–24 vs. 18–19), more vertebrae (41–43 vs. 37–38), and more lateral‐line scales (59–73 vs. 43–52).
As these specimens are morphologically distinct from all other Pollimyrus, they are here identified as a new species to science: Pollimyrus vanneeri sp. nov. (Figure 11). The detailed description of this species new to science is provided later.
Slender‐tailed group
3.2.4
A PCA on 26 log‐transformed measurements shows a slight separation of C. plagiostoma, P. pedunculatus, and P. sp. “luki” on a combination of PC 2, with the most important loadings being the caudal peduncle length, length of the dorsal fin, the posterodorsal distance, and PC 3, with the most important loadings being the snout length, the length of the dorsal fin, and the eye diameter (Figure 6b; Table S4 for the loadings; also see Figure S22). A PCA on the meristics shows a separation of C. plagiostoma on PC 1, with the most important loadings being the number of dorsal‐fin rays, pectoral‐fin rays, and scales between the anal‐fin origin and lateral line, P. pulverulentus, P. nigripinnis, P. maculipinnis, P. osborni, and P. schreyeni on PC 2, with the most important loadings being the number of teeth in the upper jaw, teeth in the lower jaw, and vertebrae, and P. maculipinnis, P. osborni, and P. schreyeni on PC 3, with the most important loadings being the number of anal‐fin rays, caudal peduncle scales, and the scales between the lateral line and dorsal‐fin origin (Figure 6d; Table S5 for the loadings).
Plots of morphometric data of slender‐tailed Pollimyrus. (a) PC 1 (proxy for SL) against PC 2 for a principal component analysis (PCA) on 26 log‐transformed measurements (n = 49). (b) PC 2 against PC 3 for a PCA on 26 log‐transformed measurements (n = 49). (c) PC 1 against PC 2 for a PCA on 12 meristics (n = 49). (d) PC 2 against PC 3 for a PCA on 12 meristics (n = 49) (also see Tables 6, 7 and S4, S5). Stars indicate holotypes and syntypes, circles indicate paratypes, and triangles indicate non‐type specimens. Explained variance is noted between brackets for each PC axis.
Cyphomyrus plagiostoma (Boulenger, 1898) (Figure S4) differs from all the other slender‐tailed Pollimyrus by having more dorsal‐fin rays (32–33 vs. 14–28) and a dorsal fin that is longer than the anal fin (130.0%–136.4% LD vs. 42.9%–111.1% LD). These characteristics also differentiate this species from all other Pollimyrus species.
Pollimyrus schreyeni Poll, 1972 (Figure S13) has fewer scales between the pelvic fin origin and the lateral line compared to the other slender‐tailed Pollimyrus (9–11 [median 10] vs. 11–18 [median 14]). It differs by the number of dorsal‐fin rays from P. pedunculatus (17 vs. 19), from P. adspersus (17 vs. 19–20), from P. pulverulentus (17 vs. 22–24), and P. nigripinnis (17 vs. 18–23). It has a dorsal fin that originates clearly posteriorly to the level of the anal‐fin origin, whereas in all other Pollimyrus species, it originates slightly anteriorly, slightly posteriorly, or at the same level as the anal fin. Its dorsal fin is slightly shorter than half the anal fin, whereas in all other species it is longer than half this length or even slightly longer than the entire length of the anal fin (42.9%–46.8% LA vs. 63.1%–136.98% LA). The anal fin is slightly longer than one third of the standard length in P. schreyeni (34.7%–35.4% SL vs. 19.9%–32.1% SL in other Pollimyrus). It has more fin rays (30–32 vs. 20–28) than all other Pollimyrus species. Further, compared to all other Pollimyrus species, the four type specimens included in the study have a distinct colouration after preservation. These fish are generally light brown with large‐sized, dark brown spots, covering several scales, whereas most other Pollimyrus species are plainly colored, with sometimes a vague midlateral line or small‐sized spots, which are smaller than a scale (see Figure S16).
Pollimyrus pedunculatus (David & Poll, 1937) (Figure S14), only known from the holotype, is the only slender‐tailed Pollimyrus species in which the anterior nostril is positioned lower than the posterior one. Further, it has a long and slim caudal peduncle compared to the other slender‐tailed Pollimyrus species (CPD%CPL; 20.6% CPL vs. 24.4%–37.6% CPL), except for C. plagiostoma, which also has a rather slender tail (19.3%–25.6% CPL). The pre‐pelvic distance is shorter compared to that of all other Pollimyrus (34.3% SL vs. 36.0%–42.9% SL). Its pelvic fin seems shorter than in all other Pollimyrus species (7.0% SL vs. 8.4%–13.4% SL). However, it cannot be ruled out that this is caused by some damage, as the distal tip of the holotype's fin seems slightly tapered.
Pollimyrus maculipinnis (Nichols and LaMonte, 1934) (Figure S15) has a holo‐ and a paratype that are small (52.9–53.9 mm SL). This species clearly differs from the other slender‐tailed Pollimyrus species by several measurements and meristics. It differs from P. nigripinnis and P. pulverulentus by having a thicker caudal peduncle (5.6%–6.4% SL vs. 3.9%–5.3% SL and 4.1%–4.7% SL) and fewer teeth in the upper jaw (7 vs. 8–10 and 9–10). It differs from P. adspersus, P. sp. “luki,” P. osborni, and P. pedunculatus by having a longer head (25.3%–25.3 SL vs. 21.5%–23.7% SL). P. maculipinnis has fewer dorsal‐fin rays compared to the specimens identified as P. nigripinnis, P. adspersus, and P. pedunculatus (17–17 vs. 19–23), and fewer anal fin rays than P. pulverulentus and P. schreyeni (24–24 vs. 27–32).
Pollimyrus nigripinnis (Boulenger, 1899) (Figure S16) and P. pulverulentus (Boulenger, 1899) (Figure S16) are hard to distinguish from each other. Based on the collected morphometric data, the type specimens of P. nigripinnis and P. pulverulentus overlap in measurements and meristics (also see Table 5). A PCA on 26 log‐transformed measurements shows a clear separation of the two nominal species on PC 2, with the most important loadings being the snout length, belly length, and the caudal peduncle length. The same is true for a PCA on the meristics on PC 2, with the most important loadings being the number of vertebrae, caudal peduncle scales, and lateral‐line scales (Figure 7; also see Tables S7 and S8 for the loadings). However, they cannot be distinguished visually on any qualitative or quantitative characteristic, and the ranges for the measurements and meristics do not allow for a clear separation due to overlap. With the type localities of both nominal species being in the Middle Congo basin (Figure 2), they might belong to one and the same species. Therefore, both species are regarded as synonyms and from now on referred to as P. pulverulentus (Boulenger, 1899) (see Discussion for more details). Non‐type specimens identified to the P. pulverulentus complex found in Lac Bleu appear to resemble the types of P. pulverulentus most morphologically, as seen in the observed ranges for the different nominal species (Table 5) and in PCAs on the measurements and meristics (Figure 7).
Plots of morphometric data of the Pollimyrus pulverulentus complex (n = 16). (a) PC 1 (proxy for SL) against PC 2 for a principal component analysis (PCA) on 26 log‐transformed measurements (n = 16). (b) PC 2 against PC 3 for a PCA on 26 log‐transformed measurements (n = 16). (c) PC 1 against PC 2 for a PCA on 12 meristics (n = 16). (d) PC 2 against PC 3 for a PCA on 12 meristics (n = 16) (also see Tables 6, 7, S2 and S3). Stars indicate syntypes, and triangles indicate non‐type specimens. Explained variance is noted between brackets for each PC axis.
Pollimyrus pulverulentus differs from P. adspersus, P. osborni, P. pedunculatus, and P. sp. “luki” by having a longer pre‐pectoral distance (26.6%–30.2% SL vs. 20.1%–25.9% SL). It also differs in dentition, having 8–10 teeth in the upper jaw and 9–11 teeth in the lower jaw, from P. osborni (7 teeth in the upper jaw) and P. adspersus (8 teeth in the lower jaw). Their teeth have small cusps that are not as obvious as those of P. adspersus, P. fasciaticeps, P. maculipinnis, P. osborni, and Pollimyrus sp. “luki”.
One of the syntypes of P. nigripinnis shows some morphological differences from the other syntypes. Six syntypes were collected from Kutu at Lake Mai‐Ndombe, and one was collected from the Uéré River (RMCA P.344: 61.3 mm SL) (Figure 2; also see Supplementary file S1 for hydro‐geographic locations). The latter has a deeper caudal peduncle relative to its body depth (18.3% CPL vs. 15.3%–16.9% CPL), a longer head (27.1% SL vs. 23.5%–25.3% SL), a shorter dorsal fin (18.5% SL vs. 20.9%–23.6% SL), a wider lower jaw (18.8 vs. 15.9%–18.6% HL), and a wider interorbital distance (38.7% HL vs. 31.1%–35.7% HL) compared to the other syntypes of P. nigripinnis from Kutu. Its mouth is positioned subterminally, whereas in the Kutu syntypes, it is positioned terminally. Further, it also has fewer dorsal‐fin rays (18 vs. 19–21) and vertebrae (37 vs. 38–40), but more upper‐jaw teeth (10 vs. 8–9) (Table 5). Finally, it is also clearly separated on a PCA of the 26 log‐transformed measurements on PC 3, with the most important loadings being the lower‐jaw width, dorsal‐fin length, and eye diameter (Figure 7). This specimen is, however, not as well preserved as the other syntypes. Even though it is possible that this specimen is simply badly preserved, some characteristics indicate that the syntype series of P. nigripinnis is polyspecific. This idea is further supported by the wide hydro‐geographical separation of Kutu and Uéré. Thus, syntype RMCA P.344 is further referred to as P. nigripinnis “Uéré,” given the clear differences in morphology from the other syntypes. Although P. osborni has been described from the Uele River, of which the Uéré River is an affluent, P. nigripinnis “Uéré” clearly differs from the holotype of P. osborni by its lower number of circumpeduncular scales (13 vs. 17 in P. osborni) and in, for example, head width (49.3% HL vs. 57.5% HL in P. osborni) (also see Tables 6 and 7). Thus, both these two specimens are clearly not conspecific despite occurring in the same (sub)basin. It is currently unclear if P. nigripinnis “Uéré” could represent a species new to science due to the lack of similar specimens from the same locality or (sub)basin.
Pollimyrus osborni (Nichols and Griscom, 1917) (Figure S17) is only known from the holotype. The species differs from most slender‐tailed species by the aforementioned characteristics for each of the species already differentiated in the complex. Further, it can be distinguished from P. adspersus by having a wider head length (57.5% HL vs. 51.1%–51.9% HL), a longer pre‐ventral distance (42.2% SL vs. 36.0%–36.7% SL), and fewer dorsal‐ (17 vs. 19–20) and anal‐fin rays (24 vs. 27). Finally, it differs from P. sp. “luki” by having more circumpeduncular scales (17 vs. 12–15) and relatively long and broad teeth that are packed closely together (vs. teeth that are smaller and have an open space between them).
Specimens found in the Luki River in the Democratic Republic of the Congo, here preliminarily identified as Pollimyrus sp. “luki,” differ from all previously mentioned slender‐tailed species as described earlier. They have overlapping ranges of all measurements with P. adspersus (Table 6). No differences in meristics could be found (Table 7). Nevertheless, the teeth of these specimens are small, whereas the teeth of P. adspersus are larger and take in more space in the upper jaw. In the lower jaw, the teeth of these specimens are more embedded and have mostly symmetrical cusps, whereas the teeth of P. adspersus are asymmetrical, with one cusp being larger than the other. In the upper jaw, all specimens found in the Luki River have a type 1 dentition (Figure 1e), whereas the type specimens of P. adspersus have either a type 2 or have an additional tooth behind the most anterior tooth. As such, the specimens found in the Luki River can be regarded as separate species based on their well‐distinct dentition. Furthermore, the species probably differ in their known geographic occurrence, with the syntypes of P. adspersus having been reported from “West Africa” (Günther, 1866). As these specimens are morphologically distinct from all other Pollimyrus, they are proposed as a new species to science: Pollimyrus ibalazambai sp. nov. (Figure 15). The detailed description of this species new to science is provided below.
Pollimyrus adspersus (Günther, 1866) (Figure S18) differs from all other slender‐tailed Pollimyrus as mentioned earlier.
P. tumifrons group
3.2.5
P. tumifrons (Boulenger, 1902) (Figure S19) and its two junior synonyms, P. aequipinnis (Pellegrin, 1924) and P. anterodorsalis (David & Poll, 1937) differ from the other Pollimyrus species (except C. plagiostoma) by having an anal fin that is slightly shorter than the dorsal fin, whereas in the other species it is about equal or longer than the dorsal fin (90.0%–92.2% LD vs. 95.7%–223.1% LD). The dorsal fin is longer in P. tumifrons than in other Pollimyrus species (29.3%–30.6% SL vs. 15.2%–25.4% SL) and shorter than in C. plagiostoma (29.3%–30.6% SL vs. 34.4%–37.9% SL). The mouth is clearly placed inferior in P. tumifrons, whereas it is rather subinferior to terminal in other Pollimyrus species. The posterior edge of the mouth is positioned beneath the eye in P. tumifrons, whereas it is never positioned under the eye in other Pollimyrus species. They also have, although only slightly, more pectoral‐fin rays (12 vs. 9–11) than most other Pollimyrus specimens (except one specimen of P. sp. “kouilou‐niari” and most specimens of C. plagiostoma). The anterior nostril is positioned lower than the posterior one in P. tumifrons, whereas the anterior one is positioned higher than the posterior one in all other Pollimyrus species, except P. pedunculatus. Although this species complex shows characteristics that are different from most congenerics and is therefore easily differentiated from the other Pollimyrus species, it remains unclear to which genus P. tumifrons would correspond better than Pollimyrus. For this reason, the species is retained within Pollimyrus.
Pollimyrus tumifrons has been described from the Ubangi River near Banzyville (nowadays: Mobayi‐Mbongo) in north‐west DRC, P. anterodorsalis from Panga (David & Poll, 1937) along the Aruwimi in north‐eastern DRC, and P. aequipinnis from the Kasaï River at N'Gombe (Pellegrin, 1924) in south‐western DRC (Figure 2; also see Supplementary file S1 for hydro‐geographic locations). Pollimyrus anterodorsalis was synonymized with P. tumifrons, by Taverne (1971a, 1971b), although, unfortunately, without further explanations. Later on, P. aequipinnis was also synonymized with P. tumifrons by Poll (1976), based on its overall similar appearance but, unfortunately, also without any further detailed explanation. However, morphologically, the holotype of P. aequipinnis has a visually small snout, whereas the holotypes of the other two nominal species have large snouts. Further, P. aequipinnis differs from both by having a shorter pre‐pelvic distance (38.5% SL vs. 40.1% SL in P. tumifrons and 43.9% SL in P. anterodorsalis) and more lateral‐line scales (67 vs. 62 in P. tumifrons and 63 in P. anterodorsalis) and more circumpeduncular scales (18 vs. 16 in P. tumifrons and 13 in P. anterodorsalis). These differences would suggest that P. aequipinnis can be a species distinct from the other two synonymized species within the P. tumifrons group. Furthermore, two studied specimens identified as P. tumifrons (RMCA 158002 and 158003) morphologically resemble the type specimens of both P. tumifrons and P. anterodorsalis in the aforementioned characteristics more than that of P. aequipinnis, although they were collected sympatrically with the last one. Indeed, both RMCA specimens originate from the Luachimo River, which is near the type locality of P. aequipinnis, the Kasaï River, with the former being an affluent of the latter. However, there are no data for specimens similar to the holotype of P. aequipinnis, and no other specimens are available from the type localities of all three synonymized species to confirm the observed differences. Therefore, the alpha‐taxonomic issues within this group cannot be resolved until more specimens become available. We therefore suggest to retain all three synonymized nominal species within a single valid species, P. tumifrons, for the time being.
New species descriptions in Pollimyrus
3.3
Class Actinopterygii Klein, 1885
Order Osteoglossiformes Berg, 1940
Family Mormyridae Bonaparte, 1831
Subfamily Mormyrinae Bonaparte, 1831
Genus Pollimyrus Taverne, 1971
Pollimyrus Taverne, 1971: 140 (type species: Mormyrus isidori Valenciennes, 1847, by original designation).
Pollimyrus ibalazambai sp. nov.
Zoobank registration: urn:lsid:zoobank.org:act:EA58D11A‐D2BB‐43E4‐982B‐4A461985C857
Figures 8 and 9; Tables 6 and 7.
Photographs of preserved specimens of Pollimyrus ibalazambai sp. nov. (a) Holotype (Royal Museum for Central Africa [RMCA] 2017.014.P.0002: 70.2 mm SL [standard length]). (b) Paratype (RMCA 2017.014.P.0020‐0021, ID = 22: 70.7 mm SL).
Photograph of a live type specimen of Pollimyrus ibalazambai sp. nov. from the Luki River, near the Kimbozi Bridge (by S.W.L., Mbisa‐Congo I, August 10, 2016).
Pollimyrus sp. “luki”
Type material
Holotype
DEMOCRATIC REPUBLIC OF THE CONGO • Luki River near Kimbozi Bridge; 5°42′3.6″ S, 12°57′35.1″ E; June 24, 2017; S. Wamuini Lukayilakio and Z.J. Kosi leg.; 70.2 mm SL; RMCA 2017.014.P.0002.
Paratypes
DEMOCRATIC REPUBLIC OF THE CONGO • one paratype; same data as holotype; 76.0 mm SL; RMCA 2017.014.P.0001 • three paratypes; same data as holotype; 67.0–79.3 mm SL; RMCA 2017.014.P.0003‐0005 • two paratypes; same data as holotype; 67.5–67.7 mm SL; AMNH 281713 • two paratypes; same data as holotype; 70.7–71.4 mm SL; RMCA 2017.014.P.0008‐0009 • six paratypes; same data as holotype; 64.6–71.9 mm SL; RMCA 2017.014.P.0010‐0015 • two paratypes; same data as holotype; 70.1–74.4 mm SL; BMNH 2024.9.20.2‐3 • two paratypes; same data as holotype; 69.8–70.2 mm SL; ZSM 49655 • two paratypes; same data as holotype; 70.7–77.5 mm SL; RMCA 2017.14.P.0020‐0021.
Diagnosis
Distinguished from its congeners by the following unique combination of characters: a slender tail (15.3%–19.2% BD vs. thicker, 21.7%–31.2% BD, in P. brevis, P. castelnaui, P. fasciaticeps, P. guttatus, P. isidori, P. krameri, P. marianne, P. nigricans, P. petricolus, P. stappersii, P. vanneeri, and P. weyli); a blocky snout (vs. rounded snout in P. brevis, P. castelnaui, P. guttatus, P. krameri, P. marianne, P. petricolus, P. stappersii, P. vanneeri, and P. weyli); pointed pectoral fin (vs. rounded in P. brevis, P. castelnaui, P. guttatus, P. krameri, P. marianne, P. petricolus, P. stappersii, P. tumifrons, P. vanneeri, and P. weyli); concave anal and dorsal fins (vs. rounded in P. brevis, P. castelnaui, P. guttatus, P. krameri, P. marianne, P. petricolus, P. stappersii, P. vanneeri, and P. weyli); absence of chin (vs. obvious chin in P. brevis, P. castelnaui, P. guttatus, P. krameri, P. marianne, P. petricolus, P. stappersii, P. vanneeri, and P. weyli); small, widely spaced, symmetrical teeth with clear cusps; a short head (20.9%–22.8% SL vs. 23.2%–27.7% SL in P. adspersus, P. maculipinnis, P. osborni, and P. pulverulentus); a short dorsal fin (20.4%–23.9% SL vs. shorter, 15.2%–19.5% SL, in P. maculipinnis and P. schreyeni, and longer, 25.0%–37.9% SL, in P. pedunculatus and C. plagiostoma); and anterior nostril positioned higher than the posterior one (vs. vice versa in P. pedunculatus). Also see Supplementary file S1 for details and a differential diagnosis with each species.
Description
Based on holotype and paratypes. Measurements and meristics are given in Tables 6 and 7. A relatively small species, with a maximum observed size of 79.3 mm SL. Body oblong or diamond shaped and dorso‐ventrally compressed. Deepest point of body around anal‐fin origin. Head blocky and less deep than body. Mouth small and subterminal, not reaching level of eye. No obvious chin. Anterior nostril positioned higher than posterior one. Posterior nostril close to eye. Nostrils separated by less than half snout–eye distance. Very small, close‐set, bicuspid teeth in a single row, six to eight in the upper jaw and seven to nine in the lower jaw. Arch of teeth not filling entire upper and lower jaw. Most anterior tooth on upper jaw clearly positioned anteriorly of second most teeth left and right of it. Anal‐fin origin slightly in front of dorsal‐fin origin. Anal fin longer than dorsal fin. Anterior parts of anal and dorsal fins higher than posterior part with concave diminution (see Figure 1 for definition). Pectoral fin long, reaching tip of or even extending beyond pelvic fin. First three pectoral‐fin rays longer than others with concave diminution. Caudal‐fin lobes somewhat rounded and long. Long and slender caudal peduncle.
Color in life
Body generally light silvery‐grayish with some greenish/olive spots on the head and dorsum and some dark gray spots on the underside of the head. Dorsal, anal, and caudal fins darker gray at base and translucent at distal end. Pectoral fins light to dark gray at base and translucent at distal end. Pelvic fins slightly pinkish and translucent at distal end. Slight darker gray midlateral stripe of one to two scales wide running from behind the head to the caudal peduncle (Figure 9).
Color in preserved specimens
Body generally light brownish with large darker brown spots on the head and smaller ones over the entire body. Head slightly darker. Dorsal side of the body and caudal peduncle dark brown. Dorsal, anal, and caudal fins yellow to brown at base and translucent at distal end. Pectoral and pelvic fins light yellow to light brown at base and translucent at distal end. Lateral line gray to black. Some specimens with darker supralateral line(s) (Figure 8).
Distribution and habitat
The specimens of P. ibalazambai were collected near the bridge of Kimbozi Village in the Luki River, a left bank affluent of the Lukunga, itself a right bank affluent of the Lower Congo River (DRC) (Figures 2 and 10). The river where the specimens were caught was about 12 m wide and 60 cm deep. The river flowed at a speed of 0.3 m/s. The banks of the river were grassy, and the canopy above the river covered about 10% of its surface only. The floor of the river consisted of mostly sand with some mud, gravel, and pebbles. The following physicochemical parameters were collected at the type locality: water temperature about 22°C, pH 7.6, salinity 0.1 ppm, and oxygen level of 7.3 mg/L.
Photograph of the habitat where the type specimens of Pollimyrus ibalazambai sp. nov. were caught: Luki River downstream from the Kimbozi Bridge (by S.W.L., MbiSa‐Congo I, August 9, 2016).
Etymology
The specific epithet is a noun honoring Professor Dr. Armel Ibala Zamba (1975–) (Université Marien Ngouabi, the Republic of the Congo) for his contributions to African ichthyology and his work in the Luki River basin (DRC) within the framework of his PhD (2005–2010).
Pollimyrus krameri sp. nov.
Zoobank registration: urn:lsid:zoobank.org:act:48E2A84F‐625C‐4891‐A7E8‐F6368805AE1E
Figures 11 and 12; Tables 6 and 7
Photographs of preserved specimens of Pollimyrus krameri sp. nov. (a) Holotype (South African Institute for Aquatic Biodiversity [SAIAB] 73892: 45.6 mm SL [standard length]). (b) Paratype (SAIAB 237203: 48.7 mm SL).
Photograph of dead, but not yet fixed, holotype specimen of Pollimyrus krameri sp. nov. (South African Institute for Aquatic Biodiversity [SAIAB] 73892: 45.6 mm SL [standard length]) from the Lugenda River (by R.B., August 22, 2003).
Pollimyrus sp. “lugenda”
Type material
Holotype
MOZAMBIQUE • Lugenda River near the Mpamanda Fishing camp; 12°26′51″ S, 37°35′45″ E; August 22, 2003; R. Bills leg.; 45.6 mm SL; SAIAB 73892.
Paratypes
MOZAMBIQUE • one paratype; same data as holotype; 48.7 mm SL; SAIAB 237203.
Diagnosis
Distinguished from its congeners by the following unique combination of characters: thick caudal peduncle (23.4%–23.4% BD vs. shallower, 12.7%–21.9% BD, in P. adspersus, P. maculipinnis, P. osborni, P. pedunculatus, P. pulverulentus, P. schreyeni, P. stappersii, P. tumifrons, P. ibalazambai, and C. plagiostoma, and deeper, 25.4%–28.8% BD, in P. petricolus); rounded pectoral fins (vs. pointed in P. adspersus, P. fasciaticeps, P. isidori, P. maculipinnis, P. nigricans, P. osborni, P. pedunculatus, P. pulverulentus, P. schreyeni, P. ibalazambai, and C. plagiostoma); rounded (similar height of rays) anterior part of dorsal and anal fins (vs. pointed in P. adspersus, P. maculipinnis, P. osborni, P. pedunculatus, P. pulverulentus, P. schreyeni, P. tumifrons, P. ibalazambai sp. nov, and C. plagiostoma); small chin (vs. no chin in P. adspersus, P. fasciaticeps, P. isidori, P. maculipinnis, P. nigricans, P. osborni, P. pedunculatus, P. pulverulentus, P. schreyeni, P. vanneeri, P. ibalazambai, and C. plagiostoma); shallow body (25.3%–26.7% SL vs. thicker, 28.2%–35.8% SL, in P. adspersus, P. brevis, P. castelnaui, P. fasciaticeps, P. guttatus, P. isidori, P. maculipinnis, P. osborni, P. pedunculatus, P. pulverulentus, P. schreyeni, P. weyli, and C. plagiostoma); short snout (13.5% HL vs. shorter, 10.5%–11.8% HL, in P. adspersus and longer, 14.9%–22.0% HL, in P. isidori, P. schreyeni, P. tumifrons, P. vanneeri, and C. plagiostoma); shorter snout–posterior side eye distance (39.4%–39.7% HL vs. longer, 39.5%–53.8% HL, in P. adspersus, P. brevis, P. castelnaui, P. fasciaticeps, P. guttatus, P. isidori, P. maculipinnis, P. marianne, P. osborni, P. pedunculatus, P. petricolus, P. pulverulentus, P. schreyeni, P. stappersii, P. tumifrons, P. ibalazambai, and C. plagiostoma); small eye (14.3%–15.5% HL vs. larger, 15.5%–28.7% HL, in P. adspersus, P. brevis, P. castelnaui, P. fasciaticeps, P. guttatus, P. isidori, P. maculipinnis, P. marianne, P. osborni, P. pedunculatus, P. petricolus, P. pulverulentus, P. schreyeni, P. tumifrons, P. weyli, P. ibalazambai, and C. plagiostoma); short posterodorsal distance (36.8%–37.0% SL vs. longer, 38.1%–52.8% SL, in P. adspersus, P. castelnaui, P. fasciaticeps, P. guttatus, P. isidori, P. maculipinnis, P. osborni, P. pedunculatus, P. petricolus, P. pulverulentus, P. schreyeni, P. tumifrons, P. vanneeri, P. ibalazambai, and C. plagiostoma); long head (24.8%–25.2% SL vs. shorter, 22.4%–23.3% SL, in P. castelnaui and P. isidori); few dorsal‐fin rays (15 vs. more, 17–33, in P. adspersus, P. brevis, P. fasciaticeps, P. guttatus, P. isidori, P. maculipinnis, P. pedunculatus, P. petricolus, P. pulverulentus, P. schreyeni, P. tumifrons, P. vanneeri, P. weyli, P. ibalazambai, and C. plagiostoma); and few circumpeduncular scales (13–14 vs. more, 16–23, in P. fasciaticeps, P. isidori, P. nigricans, P. osborni, P. pedunculatus, P. petricolus, and P. vanneeri). Also see Supplementary file S1 for details and a differential diagnosis with each species.
Description
Based on holotype and paratype. Measurements and meristics are given in Tables 6 and 7. A relatively small species, with a maximum observed size of 48.7 mm SL. Body oblong, elongated, and dorso‐ventrally compressed. Deepest point of body at anal‐fin origin, and not changing drastically over entire body. Head round and less deep than body. Mouth small and subterminal, not reaching level of eye. Slight chin. Snout protruding. Anterior nostril positioned higher than posterior one. Posterior nostril close to eye. Nostrils separated by less than snout–eye distance. Bicuspid teeth in single row, eight to nine in upper jaw and seven in lower jaw. Arch of teeth filling entire upper and lower jaw. Most anterior tooth on upper jaw clearly positioned anteriorly of second most teeth left and right of it. Anal‐fin origin posterior of dorsal‐fin origin. Anal fin longer than dorsal fin. Anterior parts of anal and dorsal fins slightly higher than posterior part. Anal‐ and dorsal‐fin edges straight. Pectoral fin rounded, reaching halfway pelvic fin. Caudal‐fin lobes rounded and broad, overlapping in middle. Thick caudal peduncle.
Color in life
Body dark brown or gray. Ventral side of head and belly light yellowish. Caudal peduncle slightly darker brown or gray. Dorsal, anal, and caudal fins dark brown at base and slightly translucent at distal end. Pectoral fins dark brown or gray and slightly translucent. Pelvic fins light gray and slightly translucent. Lateral line dark gray (Figure 12).
Color in preserved specimens
Body dark brown/reddish. Ventral side of head and belly yellow or pale yellow. Caudal peduncle slightly darker. Dorsal and anal fins brown at base and translucent at distal end. Caudal fin light brown or gray and translucent. Pectoral fins light brown or gray and translucent. Pelvic fin light brown or gray and translucent. Lateral line dark brown (Figure 11).
Distribution and habitat
The specimens of P. krameri were found in a braid channel, the Nkupo Stream, during the dry season, which is part of the Lugenda River, itself a right bank affluent of the Rovuma River (Mozambique) (Figures 2 and 13). The stream has dense marginal vegetation. The bottom of the river is sandy and rocky, without aquatic weed beds. Marginal Phragmites reeds, grasses, and trees result in significant marginal root stocks along some river edges. The small Nkupo Stream was not flowing and did not have aquatic macrophytes, but it was filled with filamentous algae and emergent plant root stocks. During the wet season, this stream would be part of the main river. The specimens were collected using seine netting into and underneath root stocks.
Photographs showing the habitat during the dry season where the specimens of Pollimyrus krameri sp. nov. were caught. (a) Main river, the Lugenda. (b) The Nkupo Stream (by R.B., August 22, 2003).
Etymology
The specific epithet is a noun honoring Professor Dr. Bernd Kramer (1943–) (University of Regensburg, Germany) for his contributions to ichthyology and study of weakly electric fish, southern African Mormyridae in particular.
Pollimyrus vanneeri sp. nov.
Zoobank registration: urn:lsid:zoobank.org:act:5925507B‐C01A‐4319‐8221‐146366363783
Photographs of preserved specimens of Pollimyrus vanneeri sp. nov. (a) Holotype (Royal Museum for Central Africa [RMCA] 1990.057.P.2878: 100.9 mm SL [standard length]); (b) Paratype (RMCA 1990.057.P.2890‐2892: 99.5 mm SL).
Pollimyrus sp. “kouilou‐niari”
Type material
Holotype
REPUBLIC OF THE CONGO • Kouilou‐Niari River near Kakamoeka; 4°8′ S, 12°4′ E; October 10, 1990; G. Teugels, L. De Vos and J. Snoeks leg.; 100.9 mm SL; RMCA 1990.057.P.2878.
Paratypes
REPUBLIC OF THE CONGO • eight paratypes; same data as holotype; 79.9–118.3 mm SL; RMCA 1990.057.P.2879‐2885 • one paratype; same data as holotype; 81.3 mm SL; AMNH 281712**•** one paratype; same data as holotype; 85.6 mm SL; BMNH 2024.9.20.1 • one paratype; same data as holotype; 83.9 mm SL; MNHN 2024‐0897 • one paratype; same data as holotype; 90.6 mm SL; ZSM 49654 • two paratypes; same data as holotype; October 12, 1990; 65.6–123.6 mm SL; RMCA 1990.057.P.2893‐2894 • three paratypes; confluence of Kissafou and Kouilou‐Niari rivers; 4°7′ S, 12°5′ E; October 11, 1990; G. Teugels, L. De Vos and J. Snoeks; 77.9–99.5 mm SL; RMCA 1990.057.P.2890‐2892.
Diagnosis
Distinguished from its congeners by the following unique combination of characters: thick caudal peduncle (20.3%–24.4% BD vs. slender, 12.7%–20.2% BD, in P. adspersus, P. maculipinnis, P. osborni, P. pedunculatus, P. pulverulentus, P. schreyeni, P. tumifrons, P. ibalazambai, and C. plagiostoma); rounded pectoral fins (vs. pointed in P. adspersus, P. fasciaticeps, P. isidori, P. maculipinnis, P. nigricans, P. osborni, P. pedunculatus, P. pulverulentus, P. schreyeni, P. ibalazambai, and C. plagiostoma); shallow body (22.6%–28.7% SL vs. deep, 29.1%–34.6% SL, in P. adspersus, P. brevis, P. castelnaui, P. guttatus., P. isidori, P. maculipinnis, P. osborni, P. schreyeni, and C. plagiostoma); short pre‐anal distance (52.4%–58.8% SL vs. long, 59.2%–65.8% SL, in P. brevis, P. castelnaui, P. guttatus, P. isidori, P. krameri, P. nigricans, P. stappersii, and P. weyli); more vertebrae (41–43 vs. fewer, 37–40, in P. adspersus, P. brevis, P. castelnaui, P. fasciaticeps, P. guttatus, P. isidori, P. maculipinnis, P. marianne, P. nigricans, P. osborni, P. krameri, P. pedunculatus, P. pulverulentus, P. stappersii, and P. weyli, and more, 44, in C. plagiostoma); 21–24 dorsal‐fin rays (vs. fewer, 14–20, in P. adspersus, P. brevis, P. castelnaui, P. fasciaticeps, P. guttatus, P. isidori, P. krameri, P. maculipinnis, P. marianne, P. osborni, P. pedunculatus, P. schreyeni, P. stappersii, and P. weyli, and more, 32–33, in C. plagiostoma); long snout (15.3%–21.0% HL vs. shorter, 10.5%–14.9% HL, in P. adspersus, P. castelnaui, P. guttatus, P. fasciaticeps, P. krameri, P. maculipinnis, P. nigricans, P. osborni, P. pedunculatus, P. petricolus, P. stappersii, and P. weyli); small eye (13.7%–21.6% HL vs. larger, 22.9%–28.7% HL, in P. adspersus, P. fasciaticeps, P. maculipinnis, P. osborni, and P. ibalazambai); long dorsal fin (22.1%–25.1% SL vs. shorter, 15.2%–20.2% SL, in P. brevis, P. castelnaui, P. krameri, P. maculipinnis, P. marianne, P. nigricans, P. schreyeni, P. stappersii, and P. weyli, and longer, 29.3%–37.9% SL, in P. tumifrons and C. plagiostoma). Also see Supplementary file S1 for details and a differential diagnosis with each species.
Description
Based on holotype and paratypes. Measurements and meristics are given in Tables 6 and 7. A relatively small species, with a maximum observed size of 123.6 mm SL. Body oblong, somewhat elongated, and dorso‐ventrally compressed. Deepest point of body around anal‐fin origin, and not changing drastically over entire body length. Head blocky and about as deep as body. Mouth small and subterminal, reaching level of eye. No obvious chin. Snout protruding. Anterior nostril positioned higher than posterior one. Posterior nostril close to eye. Nostrils separated by less than half snout–eye distance. Large close‐set, bicuspid teeth in single row, six to eight in upper jaw and five to nine in lower jaw. Arch of teeth filling entire upper and lower jaw. Three most anteriorly teeth positioned at about same level in upper jaw. Anal‐fin origin slightly in front of dorsal‐fin origin. Anal fin longer than dorsal fin. Anterior parts of anal and dorsal fin higher than posterior part with concave diminution. Pectoral fin long, reaching tip of pelvic fin. First three pectoral‐fin rays longer than others with concave diminution. Caudal‐fin lobes somewhat rounded and long. Long and deep caudal peduncle.
Color in life
Unknown.
Color in preserved specimens
Body generally light brownish or red‐brownish. Head slightly darker. Dorsal side of body darker brown in some specimens. Dorsal and anal fins brown at base and translucent at distal end. Caudal‐fin base white, yellow, or light brown, and distal end translucent. Pectoral and pelvic fins yellow/white and translucent at distal end. Lateral line light brown. Second, darker brown supralateral curved line visible across body (Figure 14).
Distribution and habitat
The specimens of P. vanneeri were collected near Kakamoeka in the Kouilou‐Niari River and at the confluence of the Kouilou‐Niari River with the Kissafou River, the latter a right bank affluent of the former, Republic of the Congo (Figure 2). The following physicochemical parameters were collected at the type locality: water temperature about 27.7°C in the afternoon; pH 7.8; oxygen level 6.8 mg/L; ammonium concentration 0.1 mg/L; nitrites concentration 0.05 mg/L; silicium concentration 1 mg/L; calcium concentration 18–26 ppm; and phosphate level 0.25 mg/L.
Etymology
The specific epithet is a noun honoring Professor Dr. Wim Van Neer (1954–) (Royal Belgian Institute of Natural Sciences, Brussels, and KU Leuven, Leuven, Belgium) for his contributions to ichthyoarchaeology in Europe and northern Africa.
Pollimyrus weyli sp. nov.
Zoobank registration: urn:lsid:zoobank.org:act:FB535D7B‐2446‐4DB6‐850A‐E688C2E3F65D
Figures 15 and 16; Tables 6 and 7
Photographs of preserved specimens of Pollimyrus weyli sp. nov. (a) Holotype (South African Institute for Aquatic Biodiversity [SAIAB] 67639: 51.43 mm SL [standard length]). (b) Paratype (SAIAB 67706: 49.1 mm SL).
Photograph of dead, but not yet fixed, holotype of Pollimyrus weyli sp. nov. (South African Institute for Aquatic Biodiversity [SAIAB] 67639: 51.43 mm SL [standard length]) from the Mussapa River (by R.B., September 27, 2002).
Pollimyrus sp. “buzi”
Type material
Holotype
MOZAMBIQUE • Mussapa River, Krongwa Stream near Dombe; 19°54′44″ S, 33°20′22″ E; September 27, 2002; R. Bills leg.; 51.4 mm SL; SAIAB 67639.
Paratypes
MOZAMBIQUE • one paratype; Lucite River, Mukombe Stream near Dombe; 19°56′38″ S, 33°29′06″ E; September 29, 2002; R. Bills leg.; 49.1 mm SL; SAIAB 67706.
Diagnosis
Distinguished from its congeners by the following unique combination of characters: wide upper jaw (22.6%–24.5% HL vs. slender, 12.7%–21.5% HL, in P. adspersus, P. castelnaui, P. fasciaticeps, P. guttatus, P. isidori, P. krameri, P. maculipinnis, P. marianne, P. nigricans, P. osborni, P. pedunculatus, P. petricolus, P. pulverulentus, P. schreyeni, P. stappersii, P. tumifrons, P. vanneeri, P. ibalazambai, and C. plagiostoma); thick caudal peduncle (23.4%–23.8% BD vs. slender, 12.7%–21.9% BD, in P. adspersus, P. maculipinnis, P. osborni, P. pedunculatus, P. pulverulentus, P. schreyeni, P. stappersii, P. tumifrons, P. ibalazambai, and C. plagiostoma, and thicker, 25.4%–28.8% BD, in P. petricolus); rounded pectoral fins (vs. pointed in P. adspersus, P. fasciaticeps, P. isidori, P. maculipinnis, P. nigricans, P. osborni, P. pedunculatus, P. pulverulentus, P. schreyeni, P. ibalazambai, and C. plagiostoma); rounded dorsal and anal fins (vs. pointed in P. adspersus, P. maculipinnis, P. osborni, P. pedunculatus, P. pulverulentus, P. schreyeni, P. tumifrons, P. ibalazambai, and C. plagiostoma); small chin (vs. no chin in P. adspersus, P. fasciaticeps, P. isidori, P. maculipinnis, P. nigricans, P. osborni, P. pedunculatus, P. pulverulentus, P. schreyeni, P. vanneeri, P. ibalazambai, and C. plagiostoma); deep body (29.6%–29.9% SL vs. shallow, 22.2%–28.7% SL, in P. fasciaticeps, P. krameri, P. pedunculatus, P. petricolus, P. vanneeri, and P. stappersii, and deeper, 31.9%–34.6% SL, in P. isidori, P. osborni, and C. plagiostoma); wide head (55.2%–58.7% HL vs. slender, 43.4%–53.8% HL, in P. adspersus, P. guttatus, P. isidori, P. krameri, P. pedunculatus, P. petricolus, and P. tumifrons); short anal fin (20.2%–22.5% SL vs. longer, 24.0%–35.4% SL, in P. adspersus, P. brevis, P. castelnaui, P. fasciaticeps, P. guttatus, P. isidori, P. maculipinnus, P. osborni, P. pedunculatus, P. pulverulentus, P. schreyeni, P. tumifrons, P. vanneeri, P. ibalazambai, and C. plagiostoma); short pectoral fin (18.1%–19.6% SL vs. longer, 21.1%–27.5% SL, in P. adspersus, P. fasciaticeps, P. guttatus, P. nigricans, P. osborni, P. pulverulentus, P. schreyeni, P. tumifrons, and P. ibalazambai); and 17 dorsal‐fin rays (vs. fewer, 15 in P. krameri, and more, 19–33, in P. adspersus, P. isidori, P. pedunculatus, P. petricolus, P. tumifrons, P. vanneeri, and C. plagiostoma). Also see Supplementary file S1 for details and a differential diagnosis with each species.
Description
Based on holotype and paratype. Measurements and meristics are given in Tables 6 and 7. A relatively small species, with a maximum observed size of 51.4 mm SL. Body oblong, somewhat elongated, and dorso‐ventrally compressed. Deepest point of body at pelvic‐fin origin, and not changing drastically over entire body length. Head round and as deep as body. Mouth small and subterminal, not reaching level of eye. Slight chin. Snout protruding. Anterior nostril positioned higher than posterior one. Posterior nostril close to eye. Nostrils separated by less than half snout–eye distance. Bicuspid teeth in single row, nine on upper jaw and seven on lower jaw. Arch of teeth filling entire upper and lower jaw. Most anterior tooth on upper jaw clearly positioned anteriorly of second most teeth left and right of it. Anal‐fin origin posterior of dorsal‐fin origin. Anal fin longer than dorsal fin. Anterior parts of anal and dorsal fin slightly higher than posterior part. Anal‐ and dorsal‐fin edges straight. Pectoral fin rounded, not reaching tip of pelvic fin. Caudal‐fin lobes rounded and broad, overlapping in middle. Deep caudal peduncle.
Color in life
Body dark brown to reddish. Ventral side of head and belly olive. Dorsal, anal, and caudal fins dark brown or olive at base and slightly translucent at distal end. Pectoral and pelvic fins light gray to light olive, mostly translucent. Lateral line slightly yellowish/red (Figure 16).
Color in preserved specimens
Body generally light brownish or yellowish. Small brown spots across body. Dorsal and anal fins brownish or yellowish at base and grayish and slightly translucent at distal end. Caudal fin brownish and slightly translucent. Pectoral and pelvic fins light yellow or light brown, slightly translucent. Lateral line slightly visible as slightly darker brown (Figure 15).
Distribution and habitat
The specimens of P. weyli were collected in the Mukombe and the Krongwa streams near Dombe of the Mussapa, an affluent to the Lucite River, and Lucite River respectively, a left bank affluent of the Buzi River, Mozambique (Figures 2 and 17). The holotype was caught in a weed‐filled non‐flowing stream close to the main channel of the Buzi River, some 154 m a.s.l. The vegetation consisted mostly of a fine leafed Potamogeton (like Potamogeton pusillus) and a few water lilies (Nymphaea). The vegetation cover was close to 100%, where the specimen was caught by raking a big hard‐framed hand‐net through the weeds. The water depth was less than 1 m with a mixed substrate of sand and rocks. The paratype was caught in a lower‐altitude tributary, at 138 m a.s.l., with a good flow, completely covered over by emergent Phragmites. There was no submerged aquatic vegetation. Collected by electrofishing with a Samus backpack. Water depth was less than 1 m with a substrate of cobbles and banks undercut with a lot of rootstocks.
Photographs showing the habitat where the holotype of Pollimyrus weyli sp. nov. was caught. (a) Upstream, where the specimens were caught. (b) Downstream (by R.B., September 27, 2002).
Etymology
The specific epithet is a noun honoring the late Professor Dr. Olaf L.F. Weyl (1972–†2020) (SAIAB) for his contributions to African ichthyology and his work in the Buzi River system within the framework of his PhD, expanding the collections housed at SAIAB and increasing the understanding of the biodiversity in the region.
Identification key to species, based on the type specimens of the genus Pollimyrus Taverne, 1971
3.4
A dichotomous identification key using the morphological features (see methods for definitions) of the type specimens of the Pollimyrus species (and C. plagiostoma for completeness: see Results) is provided below. As this key is based only on type specimens, the morphological variation possible within a species is not reflected here due to the low number of specimens available for some species. Morphological features, especially measurements, should be used cautiously if the differences between species are minimal. Photos of all species can be found in the Supplementary file S1 (Figures S1–S19).
- Anal fin twice as long as dorsal fin (LA 34.7%–35.4% SL vs. LD 15.2%–16.2% SL); 30–32 anal‐fin rays ** P. schreyeni Poll, 1972** ‐ Anal fin not twice as long as dorsal fin (LA 20.2%–30.8% SL vs. LD 16.0–37.9% SL); 29 or fewer anal‐fin rays .…………………… 2
- 32–33 dorsal‐fin rays; dorsal fin slightly longer than anal fin (LD 34.4%–37.9% SL vs. LA 26.5%–27.8% SL) ** C. plagiostoma (Boulenger, 1898)** ‐ 14–28 dorsal‐fin rays; dorsal and anal fin of similar length (LD 16.0%–30.6% SL vs. LA 20.2%–30.8% SL)…………………………..3
- 12 pectoral‐fin rays; anterior nostril positioned lower than the posterior nostril; inferior mouth; posterior part of the mouth reaching level of the eye ** P. tumifrons (Boulenger, 1902)** ‐ No such combination of characteristics ………………………………… 4
- Anterior nostril positioned lower than the posterior nostril; very long (CPL 22.8% SL) and slender (CPD 20.6% CPL) caudal peduncle ** P. pedunculatus (David & Poll, 1937)** ‐ Anterior nostril positioned higher or same level as posterior nostril; caudal peduncle not as long (CPL 13.6%–20.8% SL) and slender (CPD 25.0%–52.5% CPL) ………………………………5
- Caudal peduncle depth about five times or more in body depth at pelvic‐fin origin (CPD 12.7%–20.2% BD)6 ‐ Caudal peduncle depth less than five times in body depth at pelvic‐fin origin (CPD 21.5%–31.2% BD) ………………………………10
- Caudal peduncle depth about five times in body depth at pelvic‐fin origin (CPD 19.4%–20.2% BD); 7 teeth in upper jaw; 17 dorsal‐fin rays; 24 anal‐fin rays** P. maculipinnis (Nichols & LaMonte, 1934)** ‐ No such combination of characteristics ………………………………7
- Long pre‐pectoral distance (PPL 26.6%–30.2% SL); tooth cusps not protruding and small ** P. pulverulentus (Boulenger, 1899) (including P. nigripinnis [Boulenger, 1899])** ‐ Short pre‐pectoral distance (PPL 22.5%–25.9% SL); protruding tooth cusps 8
- Teeth small and with clear spaces between them P. ibalazambai sp. nov.‐ Teeth large and filling the jaws, that is, almost without space between them 9
- Head wide (HW 57.5% HL); long pre‐pelvic distance (PVL 42.2% SL); 17 dorsal‐ and 24 anal‐fin rays ** P. osborni (Nichols & Griscom, 1917)** ‐ Head slender (HW 51.1%–51.9% HL); short pre‐pelvic distance (PVL 36.0%–36.7% SL); 19–20 dorsal‐ and 27 anal‐fin rays ** P. adspersus (Günther, 1866)**
- Rounded snout; small chin (i.e., enlargement at frontal side of lower jaw) 11 ‐ Blocky snout; no chin ………………………………18
- 58–67 lateral‐line scales; 19–22 dorsal‐fin rays; 41–43 vertebrae; long dorsal fin (LD 89.2%–112.3% BD) ** P. petricolus (Daget, 1954)** ‐ 40–54 lateral‐line scales; 15–19 dorsal‐fin rays; 36–41 vertebrae; short dorsal fin (LD 58.6%–81.9% BD) ………………………… 12
- Slender caudal peduncle (CPD 5.9%–6.1% SL); long belly (BL 20.8%–21.6% SL) ** P. stappersii (Boulenger, 1915)** ‐ Thick caudal peduncle (CPD 6.6%–8.8% SL); short belly (BL 17.0%–20.9% SL) ………………………………13
- Broad lower jaw (LJW 23.5%–25.7% HL); 18 dorsal‐fin rays; 23–25 anal‐fin rays; 39 vertebrae ** P. brevis (Boulenger, 1913)** ‐ Slender lower jaw (LJW 13.6%–22.1% HL); no such combination of characteristics ………………………………14
- Broad upper jaw (UJW 22.6%–24.4% HL) P. weyli sp. nov.‐ Slender upper jaw (UJW 13.3%–20.2% HL) ………………………… 15
- Large eye diameter (OD 22.1%–27.3% HL); long pectoral fins (LP 23.2%–26.1% SL); deep body (BD 30.7%–32.1% SL); 18–19 dorsal‐fin rays ** P. guttatus (Fowler, 1936)** ‐ No such combination of characteristics ……………………………… 16
- Deep body (BD 29.5%–31.2% SL); wide lower jaw (LJW 21.7%–22.1% HL); pelvic fin less than half the length of belly (LV 9.7% SL vs. BL 19.6%–20.7% SL) ** P. castelnaui (Boulenger, 1911)** ‐ Shallow body (BD 25.2%–29.8% SL); slender lower jaw (LJW 13.6%–21.1% HL); pelvic fin slightly longer than half the belly length (LV 9.5%–11.9% SL vs. BL 17.0%–20.1% SL) ………………17
- Short post‐dorsal distance (pD 36.8%–37.0% SL); slender head (HW 52.7%–53.8% HL); small eye (OD 14.3%–15.5% HL); short snout length (SPE 39.4%–39.7% HL) P. krameri sp. nov.‐ No such combination of characteristics ………………… ** P. marianne Kramer et al., 2013 (including P. cuandoensis Kramer et al., 2013)**
- Rounded pectoral fins; 21–24 dorsal‐fin rays; 59–73 lateral‐line scales; 41–43 vertebrae P. vanneeri sp. nov.‐ Pointed pectoral fins; 14–20 dorsal‐fin rays; 48–55 lateral‐line scales; 37–39 vertebrae ………………………………19
- Short dorsal fin (LD 16.0%–19.1% SL); short post‐dorsal distance (pD 35.5%–37.5% SL); 12 scales between pelvic‐fin origin and lateral line ** P. nigricans (Boulenger, 1906)** ‐ Long dorsal fin (LD 22.3%–22.5% SL); long post‐dorsal distance (pD 42.8%–43.5% SL); 14–16 scales between pelvic fin origin and lateral line ………………………………20
- 20 dorsal‐fin rays; deep body (BD 33.3% SL); slender head (HW 50.9% HL) ** P. isidori (Valenciennes, 1847)** ‐ 17 dorsal‐fin rays; shallow body (BD 29.7% SL); wide head (HW 59.7% HL) ** P. fasciaticeps (Boulenger, 1920)**
Comparative material
3.5
All studied specimens of previously described species are listed below, ordered alphabetically, first, by their genus and second, by species name. The nomenclature and taxonomy proposed in this study are followed. Proposed junior synonyms are listed under their senior synonyms.
Cyphomyrus petherici (Boulenger, 1898)
SUDAN • three syntypes; Upper Nile River, Khartoum; ~ 15°36′ N, 32°32′ E; Petherick leg.; 127.5/175.3/194.2 mm SL; BMNH 1862.6.17.92/BMNH 1862.6.17.22‐23.
Cyphomyrus plagiostoma (Boulenger, 1898)
DEMOCRATIC REPUBLIC OF THE CONGO • two syntypes; Matadi; ~ 5°49′ S, 13°27′ E; Wilverth leg.; 88.4–100.2 mm SL; BMNH 1898.11.12.13‐14.
Paramormyrops eburneensis (Bigorne, 1991)
IVORY COAST • one holotype; Agnébi, Agnibilékrou; ~ 7°08′ N, 3°12′ E; April 1964; de Rham leg.; 72.9 mm SL; MNHN IC‐1990.0376 • one paratype; same data as holotype; 75.6 mm SL; MNHN IC‐1970.0006 • one paratype; same data as holotype; May 1970; 48.1 mm SL; MNHN IC‐1970.0086 • one paratype; San Pedro; ~ 4°44′ N, 6°37′ E; February 1977; Lévêque and Paugy leg.; 53.2 mm SL; MNHN IC‐1979.0135 • four paratypes; Banco, close to Abidjan, stream flowing from the Ebié Lagune; ~ 5°22′ N, 4°03′ W; 1966; Thys van den Audenaerde leg.; 59.1–75.9 mm SL; RMCA 1973.005.P.0049‐0053.
Pollimyrus adspersus (Günther, 1866)
WEST AFRICA • two syntypes; Steven and Damon leg.; 57.9–58.9 mm SL; BMNH 1865.5.3.41.
Pollimyrus brevis (Boulenger, 1913)
DEMOCRATIC REPUBLIC OF THE CONGO • one syntype; Upper Uele River near Dungu; 3°37′ N, 28°33′ E; Hutereau leg.; 54.0 mm SL; RMCA P.1804 • one syntype; same data as previous; 50.2 mm SL; BMNH 1912.12.6.1.
Pollimyrus castelnaui (Boulenger, 1911)
BOTSWANA • two syntypes; Lake Ngami; ~ 20°30′ S, 22°40′ E; Woosman leg.; 58.3–62.5 mm SL; BMNH 1910.5.31.11‐12.
Pollimyrus fasciaticeps (Boulenger, 1920)
DEMOCRATIC REPUBLIC OF THE CONGO • one holotype; Léopoldville (now Kinshasa); 4°18′ S, 15°18′ E; 1912; Christy leg.;49.3 mm SL; RMCA P.7243.
Pollimyrus guttatus (Fowler, 1936)
CAMEROON • one holotype; 30 km east of Kribi; 2°57′ N, 9°55′ E; November 26, 1934; Vanderbilt and von Blixen leg.; 39.5 mm SL; ANSP 65504 • one paratype; same data as holotype; 36.3 mm SL; ANSP 65505 • two paratypes; same data as holotype; November 24–25, 1934; 37.0–39.2 mm SL; ANSP 65506.
Pollimyrus isidori (Valenciennes, 1847)
EGYPT • one holotype; Nile River; 1799; Geoffroy Saint‐Hilaire leg.; 84.3 mm SL; MNHN IC‐4209.
Pollimyrus maculipinnis (Nichols and LaMonte, 1934)
DEMOCRATIC REPUBLIC OF THE CONGO • one holotype; Lulua River, near Luluabourg (now Kananga); ~ 5°54′ S, 22°25′ E; August 7, 1932; Callewaert leg.; 52.9 mm SL; AMNH I‐12355 • one paratype; same data as holotype; August 2, 1932; 53.9 mm SL; AMNH I‐12400.
Pollimyrus marianne Kramer et al., 2003.
NAMIBIA/ZAMBIA • 1 holotype; Zambezi River, Lisikili backwater; 17°29′ S, 24°26′ E; April 7, 1996; Kramer and van der Bank leg.; 63.9 mm SL; SAIAB 66943 • 10 paratypes; same data as holotype; 57.9–66.6 mm SL; SAIAB 66944 • 1 specimen.
Pollimyrus cuandoensis Kramer et al., 2013 (now synonym of P. marianne)
NAMIBIA • 1 holotype; Kwando River at Kongola Bridge; 17°47′33″ S, 23°20′33″ E; August 25, 1999; Kramer, van der Bank and Wink leg.; 44.1 mm SL; ZSM 41805 • 11 paratypes; same data as holotype; 38.1–52.7 mm SL; ZSM 39522.
Pollimyrus nigricans (Boulenger, 1906)
UGANDA • nine syntypes; mouth of Katonga River near Lake Victoria; ~ 0°02′ S, 32°01′ E; November 5, 1905; Degen leg.; 76.9–86.8 mm SL; BMNH 1906.5.30.85‐94.
Pollimyrus osborni (Nichols and Griscom, 1917)
DEMOCRATIC REPUBLIC OF THE CONGO • one holotype; Uele River; ~ 4°09′ N, 22°26′ E; 1909; Lang and Chapin leg.; 53.0 mm SL; AMNH I‐6934.
Pollimyrus pedunculatus (David & Poll, 1937)
DEMOCRATIC REPUBLIC OF THE CONGO • one holotype; Congo River, Boma; 5°50′ S, 13°3′ E; Van Delft leg.; 67.7 mm SL; RMCA P.22664.
Pollimyrus petricolus (Daget, 1954)
MALI • one holotype; Upper Niger, Markala; 13°41′ 15″ N, 6°05′ 13″ W; 1954?; 57.9 mm SL; MNHN IC‐1954.0008 • two paratypes; same data as holotype; April 24, 1951; Daget leg.; 50.3–85.1 mm SL; MNHN IC‐1960.0406 • one paratype; same data as holotype; May 25, 1952; Daget leg.; 52.7 mm SL; MNHN IC‐1960.0407.
Pollimyrus pulverulentus (Boulenger, 1899)
DEMOCRATIC REPUBLIC OF THE CONGO • one syntype; Coquilhatville (now Mbandaka); 0°4′ N, 18°16′ E; Delhez leg.; 89.9 mm SL; RMCA P.612 • one syntype; same data as previous; 88.8 mm SL; RMCA P.613 • two syntypes; same data as previous; 61.6–80.8 mm SL; BMNH 1899.9.26.26‐27.
Pollimyrus nigripinnis (Boulenger, 1899)
DEMOCRATIC REPUBLIC OF THE CONGO • one syntype; Lake Mai‐Ndombe/Lake Leopold II (now Lake Mai Ndombe), Kutu; 2°44′ S, 18°8′ E; Delhez leg.; 78.8 mm SL; RMCA P.608 • one syntype; same data as previous; 88.5 mm SL; RMCA P.609 • one syntype; same data as previous; 102.2 mm SL; RMCA P.610 • one syntype; same data as previous; 86.9 mm SL; RMCA P.611 • two syntypes; same data as previous; 72.5–95.2 mm SL; BMNH 1899.9.26.28–29 • one syntype; Uéré River; 4°2′ N, 25°51′ E; Debauw leg.; 61.3 mm SL; RMCA P.344.
REPUBLIC OF THE CONGO • five specimens; Lefini River basin, Lac Bleu; 3°19.07′ S, 15°28.74′ E; August 16, 2007; Ibala Zamba leg.; 63.4–114.3 mm SL; RMCA 2007.031.P.0388–0392.
Pollimyrus schreyeni Poll, 1972
DEMOCRATIC REPUBLIC OF THE CONGO • one holotype; river 15 km from Boende, along route Boende‐Watsi; 0°14′ S, 20°57′ E; May 1970; Brichard leg.; 70.5 mm SL; RMCA P.174708 • two paratypes; same data as holotype; 71.3–75.8 mm SL; RMCA P.174709–19 • one paratype; Bokuma; 0°6′ S, 18°41′ E; June 1953; Lootens leg.; 73.15 mm SL; RMCA P.88293.
Pollimyrus stappersii (Boulenger, 1915)
DEMOCRATIC REPUBLIC OF THE CONGO • one holotype; Lukinda River basin; 8°31′ S, 28°58′ E; February 26, 1912; Stappers leg.; 59.3 mm SL; RMCA P.12689 • one holotype of P. kapangae (David, 1935); Katanga, Kapanga; 11°30′ S, 26°45′ E; 1933; Overlaet leg.; 61.6 mm SL; RMCA P.39492.
Pollimyrus tumifrons (Boulenger, 1902)
DEMOCRATIC REPUBLIC OF THE CONGO • one holotype; Ubangi River, Banzyville (now Mobayi‐Mbongo); 4°18′ N, 21°10′ E; 1901; Royaux leg.; 88.6 mm SL; RMCA P.1160 • one holotype of P. aequipinnis (Pellegrin, 1924); Kasaï River, N'Gombe; 6°35′ S, 20°43′ E; Schoutenden leg.; 90.1 mm SL; RMCA P.15188 • one holotype of P. anterodorsalis (David & Poll, 1937); Aruwimi River, Panga; 1°52′ N, 26°23′ E; May 26, 1926; Bock leg.; 44.8 mm SL; RMCA P.22167.
DISCUSSION
4
An updated diagnosis of the genus Pollimyrus
4.1
This study provides a long‐overdue morphometric alpha‐taxonomic summary of the genus Pollimyrus. To date, this genus contained 18 valid species (see Table 1), which has now been revised to 20. Since its original description by Taverne (1971a, 1971b), this genus has been dealing with quite numerous reallocations of species to different genera (Table 1). Nevertheless, this seems to have resulted in some apparently incorrect or undecisive generic attributions. By morphologically analysing most available type specimens of these species (Table 2), their generic attribution status has been clarified or, otherwise, discussed.
A closer look at the type specimens of “Pollimyrus” eburneensis reveals that these do not belong to Pollimyrus, as suggested by Rich et al. (2017) as a taxonomic note with no further explanation, but rather Paramormyrops Taverne, Thys van den Audenaerde, and Heymer, 1977. Several features, especially the fusion of ventral hypurals and body depth, do not match the characteristics of Pollimyrus spp. This confirms the need to reallocate this species to another genus. Following the identification key to the mormyrid genera, as provided by Sullivan et al. (2016), “P”. eburneensis fits in the genus Paramormyrops. Paramormyrops eburneensis (Bigorne, 1991) was originally described as a subspecies of Paramormyrops kingsleyae (Günther, 1896), which at the time was assigned to the genus Pollimyrus. Pollimyrus kingsleyae has been reassigned to Brienomyrus by Teugels and Hopkins (1998) and later to Paramormyrops by Hopkins et al. (2007) due to this species having the two ventral hypurals unfused, resulting in a total of five hypurals, whereas in Pollimyrus these are fused, resulting in a total number of four hypurals only. In addition, its elongated body shape (body depth 4.7 times in SL vs. 2.7–4.2 times in species of the genus Pollimyrus [Teugels & Hopkins, 1998]) confirms its identification as a member of that genus. Regarding the status of Paramormyrops eburneensis as a potential subspecies of Paramormyrops kingsleyae, no further remarks are made here, as this falls outside the scope of this study.
As already mentioned by Lavoué et al. (2010), “Petrocephalus” guttatus does not show some of the diagnostic characteristics of that genus, such as the closely positioned nostrils and the position of the mouth at the level of the eyes. It does, however, resemble P. isidori and other Pollimyrus spp. for those characteristics (Lavoué et al., 2010; see Results). In addition, all meristics fall within the ranges known for Pollimyrus species (see Tables 6 and 7). Because there is no further resemblance to Petrocephalus, the species is here reassigned to Pollimyrus.
C. plagiostoma shares diagnostic characteristics not only with Pollimyrus but also with Cyphomyrus. The number of teeth and the caudal peduncle depth would indicate that this species can indeed be assigned to Pollimyrus, whereas the fin implantation and dorsal‐fin length contradict this, making Cyphomyrus a better generic assignment. Therefore, Myers (1960) placed this species in Cyphomyrus when erecting the genus, after which Taverne (1971a, 1971b) reassigned the species to the genus Pollimyrus based on it having only fused instead of unfused ventral hypurals, which he considered diagnostic for the genus Pollimyrus. The usefulness of the fusion of the ventral hypurals to distinguish Pollimyrus from Cyphomyrus is, however, uncertain. The type specimens of C. psittacus, the type species of the genus Cyphomyrus, have unfused ventral hypurals (Taverne, 1971a, 1971b), whereas the three syntype specimens of C. petherici have only fused ventral hypurals (see X‐rays in Supplementary Information S1), explaining its assignment to Pollimyrus by Taverne (1971a, 1971b). This, however, shows that this osteological feature is not sufficient to differentiate both genera. Recently C. plagiostoma was reassigned to Cyphomyrus based on morphological similarities to other Cyphomyrus species and unpublished genetic data (Stiassny et al., 2021). A more recent, whole‐genome analysis, however, placed this species within Pollimyrus (Peterson et al., 2022). Although the results of Peterson et al. (2022) are not in discussion, due to the absence of published photographs, it was not possible to assess morphological similarities of these genetically analysed specimens (one being CUMV 96188 from Boyoma Falls), with the type specimens from Matadi. Therefore, the present generic assignment of C. plagiostoma is here retained as Cyphomyrus for reasons of nomenclatorial stability. A thorough morphological, osteological, and genetic comparison of both genera Pollimyrus and Cyphomyrus is needed to further clarify the issue. At this point, only the dorsal‐fin length and the number of dorsal‐fin rays make it possible to unmistakably differentiate Cyphomyrus from Pollimyrus. Cyphomyrus plagiostoma is therefore easily distinguished from all Pollimyrus species (see Results and Tables 6 and 7 for details).
Pollimyrus tumifrons was retained within the genus for nomenclatorial stability, although several characteristics seem atypical for the genus (see Results). Following Sullivan et al. (2016)'s identification key, Pollimyrus remains the best fit for this species, despite there being some confusion regarding the relative length of dorsal and anal fins and the number of fin rays as diagnostic features between Pollimyrus and Hippopotamyrus Pappenheim, 1906. Recently, Hippopotamyrus macroterops (Boulenger, 1920) was reassigned to Pollimyrus by Sullivan et al. (2022) due to morphological similarities with P. tumifrons and C. plagiostoma. We agree that P. macroterops and P. tumifrons are morphologically very similar based on photographs of P. macroterops (Sullivan & Lavoué, 2024) and have a similar hydro‐geographical distribution in the Ubangi basin. As P. macroterops was not assigned to Pollimyrus at the start of this study, it was overlooked as a potential Pollimyrus species and not included in the comparative material. This recent reassignment and the uncertainty of P. tumifrons being correctly assigned to the genus Pollimyrus show that the diagnosis between Pollimyrus and Hippopotamyrus should be re‐examined.
These examples illustrate that the generic placement of some species within the Mormyridae is still in need of further taxonomic attention. Further, due to these generic misplacements, it also illustrates the difficulties one might be confronted with when trying to attribute specimens to an existing or potentially new species for science. Nevertheless, this study has allowed for an update of the morphological diagnosis of the genus Pollimyrus, sensu Taverne (1971a, 1971b, 1972), Bigorne (2003), and Hopkins et al. (2007), after the inclusion and exclusion of species since its original description. The species presently attributed to the genus Pollimyrus have a small lateral ethmoid bone, a large and curved mesethmoid bone, six circumorbital bones, with the antorbital and first infraorbital not fused, and fused or potentially unfused ventral hypural bones. Further, they have a short or rather elongated body. Their snout is shorter than the postorbital part of the head. The mouth is terminal, inferior, or subinferior. The caudal peduncle depth fits two to five times in the peduncle length. Meristically, they have 14–28 dorsal‐fin rays, 20–32 anal‐fin rays, 9–12 pectoral‐fin rays, 35–73 lateral‐line scales, 7–21/8–23 scales in transverse line, 8–18/6–20 scales in transverse line between the dorsal‐ and anal‐fin origins, 11–23 scales around the caudal peduncle, 5–10/5–11 bicuspid teeth on the oral jaws, and 37–45 vertebrae (Taverne, 1971a, 1971b, 1972; this study). Besides the aforementioned characters, these species can further be recognized by newly added characteristics following later authors: rounded or blocky head; no or very small mental lobe; nostrils that are well separated with posterior one lying close to anterior rim of eye; and dorsal‐ and anal‐fin origins at the same level (Bigorne, 2003; Hopkins et al., 2007; this study).
Intrageneric taxonomic changes
4.2
The morphological groups identified based on the caudal peduncle depth and several qualitative characteristics are confirmed by whole‐genome evidence, albeit using non‐type specimens (see Peterson et al., 2022; Figure 2). These authors found that the 12 analysed taxa, including C. plagiostoma, clearly form two well‐supported clades, one with thick‐tailed and one with slender‐tailed Pollimyrus. If confirmed with further genetic evidence, this distinction between the morphological groups could potentially be formalized.
Several of the studied nominal species were found to show little‐to‐no morphological differentiation and therefore confirmed to be synonyms or, for the first time here, identified as synonyms.
Pollimyrus pulverulentus and P. nigripinnis cannot be distinguished from each other based on the studied meristic and morphometric characteristics (Table 5). Nevertheless, they can be separated using a PCA analysis of both log‐transformed measurements and meristics when considering only these taxa (Figure 6; Supplementary file S1). The measurements and meristics with the most important loadings found in the PCA, however, show a strong overlap. Both species occur relatively close hydro‐geographically, that is, Mbandaka (formerly Coquilhatville), situated on the Middle Congo River itself, for the former, and Kutu at Lake Mai‐Ndombe, which is part of the Middle Congo River for the latter (Figure 2). The minor morphometric differences between the two species detected using PCA could be linked to their distinct hydro‐geographical occurrences or local environmental influences. Therefore, both these species are here regarded as synonyms. Both species seem to be phylogenetically closely related, as shown using non‐type specimens, although a specimen identified as P. osborni falls in the same clade (Peterson et al., 2022; Figure 2). Because both species were described in the same publication (Boulenger, 1899), there is no seniority of either of these names (ICZN, 1999: Art. 23). Both species are reported to have black spots on the body (Boulenger, 1899). As such, the name “pulverulentus,” which could refer to black freckles (Scharpf, 2024), still applies to all type specimens of both the nominal species. Pollimyrus nigripinnis is here identified as a junior synonym of P. pulverulentus.
Nevertheless, one of the syntypes of P. nigripinnis (RMCA P.344), now a junior synonym of P. pulverulentus, was collected from the Uéré River, a right bank affluent of the Uele River, itself a tributary of the Ubangi River (Middle Congo basin) in the north of the DRC near the village of Ango, whereas all other syntypes of the species originate from Lake Mai‐Ndombe, a right bank affluent of the Kasai basin (Cuvette Centrale/Middle Congo basin) in the west of the DRC near the town of Kutu. The type specimen from Uéré clearly differs from the other syntypes in its morphology, that is, by having a deeper caudal peduncle, wider and longer head, shorter dorsal fin, wider lower jaw, smaller interorbital distance, subterminal mouth, fewer dorsal‐fin rays and vertebrae, and more upper jaw teeth (Table 5 and Results). Because this is the only type specimen from that locality and given that it is not very well preserved, it remains difficult to assess its conspecificity with the other type specimens of the species. Therefore, one should be cautious to make any interpretations regarding P. nigripinnis based on this specimen.
No morphometric differences were found between P. cuandoensis and P. marianne (Figure 4), both species that occur in the Upper Zambezi basin (Figure 2). They only separate on PC 1 in the PCA, which is a proxy for size (see Methodology). Kramer et al. (2013) described morphological differences in the mean and medians of measurements and meristics between both P. cuandoensis and P. marianne, such as the number of circumpeduncular scales, lateral‐line scales, and dorsal‐fin rays, caudal peduncle depth, length of anal fin, body depth, eye diameter, snout–posterior side of the eye, and pectoral–pelvic fin distance. However, these differences were not found in the present study. When considering the full ranges of measurements and meristics, these species cannot be morphologically distinguished on any of the studied characteristics (see Tables 6 and 7). In the original species description of P. cuandoensis, genetic ISSR profiling did show some differentiation between both species (Kramer et al., 2013; Table 2). However, the suggested diagnostic black electrogel band did not show consistently within P. cuandoensis. Further, a molecular phylogram based on cytochrome b did not show a clear clustering of P. marianne versus P. cuandoensis either (Kramer et al., 2013; Figure 7). Therefore, we do not consider the genetic differences to be sufficient to validate P. cuandoensis as a separate species. These two species are most easily distinguished by their EOD patterns (Kramer et al., 2013). We are, however, hesitant to only use EOD signals and their characteristics to define species, due to their largely unknown intra‐/interspecific variation and differentiation, which is dependent on, for example, life‐history traits and the local environments (e.g., Bass & Hopkins, 1985; Carlson et al., 2000; Nguyen et al., 2020; Terleph & Moller, 2003; Westby & Kirschbaum, 1978). The EOD differences reported could thus also be due to size differences, as the P. cuandoensis specimens are of smaller size (38.11–52.7 mm SL) compared to the larger‐sized specimens (57.93–66.6 mm SL) identified as P. marianne. Therefore, those EOD differences do not necessarily support the identification of distinct species (Nguyen et al., 2020; Westby & Kirschbaum, 1978). The difference in size, and thus likely in age, could also explain the small difference in dentition observed. The teeth on the frontal part of the jaws seem slightly larger than those on the posterior part of the jaw in P. marianne; however, this was not true for P. cuandoensis. No information on ontogenetic changes in dentition of Mormyridae in literature, however, could be found. Therefore, P. cuandoensis is here identified as a junior synonym of P. marianne, as there is no clear argument that they are, in fact, separate parapatric species occurring in adjacent rivers of the Upper Zambezi basin.
Possible synonymy of P. stappersii and P. castelnaui was already noticed by Bell‐Cross (1976). Their close phylogenetic relationship has also recently been confirmed using whole‐genome analysis of non‐type specimens (Peterson et al., 2022; Figure 2). The only morphological difference reported by Bell‐Cross (1976) was the higher number of lateral‐line scales of P. castelnaui (46–53) compared with P. stappersii (no numbers reported). This difference was, however, not observed in the present study. In fact, the opposite seems to be true (47 lateral‐line scales in P. castelnaui and 48–53 in P. stappersii). Only one difference in meristics has been found in this study: the number of scales between the lateral line and the anal‐fin origin (Table 7). Further, a single difference in measurements was found during this study that allows for their diagnosis, that is, the width of the lower jaw. As a result, two independent characteristics, one meristic and one measurement, seem to support their distinct species status. Because they occur in different river systems, with P. castelnaui originally described from Lake Ngami basin of the Okavango basin in Botswana (Zambezi IP) (Boulenger, 1911), whereas P. stappersii was described from the Lukinda River basin in the Upper Congo (Congo IP) (Boulenger, 1915), they are considered valid species here due to their clear geographical isolation.
Although P. fasciaticeps has been regarded as a synonym of P. isidori or P. osborni in the past, in this study, we have found sufficient morphological evidence in the length between snout and eye, head width, and caudal peduncle depth to revalidate P. fasciaticeps as a species, distinct from P. isidori and P. osborni. There remain, however, several uncertainties regarding the identification of non‐type specimens of the complex of P. fasciaticeps and P. osborni. Both species are regarded as valid species in this study. No morphological differences were found between the examined non‐type specimens previously identified as P. fasciaticeps collected from the Lower Congo and the Léfini River in the Middle Congo, the non‐type specimens of P. osborni collected from the Lower and Middle Congo, and the holotype of P. osborni originating from Uele (Ubanghi) in the Middle Congo. As such, all the specimens from the Congo basin can be identified as P. osborni, as also previously reported by Konan et al. (2013). As a result, the holotype of P. fasciaticeps remains the only known specimen of this species, as no morphologically similar specimens, to our knowledge, have been found. The holotype represents a well‐distinct species as it differs from both P. isidori and P. osborni, two species to which it has been placed as a synonym and/or as subspecies in the past (Fricke et al., 2024), in several morphological characteristics (see Results for details).
Species complexes in need of further alpha‐taxonomic attention
4.3
Several nominal (sub)species of Pollimyrus have been synonymized with other species throughout the taxonomic history of the genus (Table 1). For a few of these, the holotypes of these synonymized species were available for study. This resulted in some new insights into their morphology and taxonomic validity.
Within P. tumifrons, there appears to be some morphological differentiation between the three previously synonymized species, with P. tumifrons and P. anterodorsalis resembling each other more than they each resemble P. aequipinnis (see Results). Unfortunately, Poll (1976) did not provide an explanation for the synonymization of P. aequipinnis with P. tumifrons. Non‐types from near the type locality of the latter species also resemble P. tumifrons and P. anterodorsalis more. It is therefore possible that both these identified morphotypes have large and, somehow, overlapping geographical distributions. However, to avoid more taxonomic confusion and possible nomenclatorial instability, P. aequipinnis is kept as a doubtful junior synonym of P. tumifrons until a further in‐depth comparative study, encompassing morphological, biogeographical, and, if possible, genetic data, clarifies this morphological group.
The present morphological and distribution data for P. stappersii and its subspecies P. kapangae seem to suggest that both might be valid species: P. stappersii itself and P. kapangae. The former was described from the Lukinda River basin, which is part of the Upper Congo basin. The latter subspecies was originally described as such by David (1935) from Shaba (now Katanga) in the Upper Congo basin based on three morphological differences in the following characteristics: number of anal‐fin rays (found here as well), number of lateral‐line scales (found here as well, but uncertainty remains), and the colouration (could not be verified due to discolouration in preservation). In addition to these, we also found a difference in the shape of the teeth. There were only a handful of specimens assigned to this species complex available for the study. However, none of these non‐type specimens originate from the same localities or habitat as the holotypes of both these current subspecies. Three specimens in the collections at RMCA were found near Lukonzolwa at Lake Mweru, which is potentially a different habitat close to the Lukinda River, the type locality of P. s. stappersii. No non‐type specimens of P. s. kapangae were known at the time of the study. Although two clear morphological differences were found, these preliminary results should be handled with care, considering the small amount of currently available specimens. Therefore, both species are here considered conspecific until more specimens become available for a more in‐depth integrative, comparative study, encompassing morphological, biogeographical, and genetic data.
The two syntypes of P. adspersus are reported to have been collected from West Africa (Günther, 1866). Unfortunately, no precise location is known for these specimens. Pollimyrus petricolus has also been reported from this part of the continent, that is, the upper and middle Niger (Bigorne, 2003), and many specimens found from the Senegal to the Niger and Chad basins have been assigned to the species P. isidori (Bigorne, 2003). Both these two species differ from P. adspersus morphologically and even belong to different morphological groups (see Results). As a result of the imprecise type locality of P. adspersus and the lack of a detailed morphological comparison of Pollimyrus species available till now, the species has been used as a waste‐basket taxon. Museum specimens caught from the Nilo‐Sudan IP (including the Volta River in Ghana), the Lower Guinea IP (e.g., Cross River and Wouri River in Cameroon) (Hopkins et al., 2007), and even the Congo IP (e.g., in Tshopo, Yangole, and Lindi rivers of the Middle Congo near Kisangani, and the towns of Boma and Matadi on the Lower Congo; see RMCA fish collection) have been identified as P. adspersus. A simple analysis of both syntypes and two non‐type specimens from the Zio River near the town of Lomé in Togo (RMCA fish collections), which were previously identified as P. adspersus, was performed. The two analysed specimens from the Zio River slightly differed morphologically from both the syntypes in body depth, head width, eye diameter, snout length, and caudal peduncle depth (see Supplementary file S2). This indicates that the taxonomy of the West African Pollimyrus requires further attention. However, due to a lack of specimens available during the study and as it fell outside the scope of this study, it was not possible to include an in‐depth comparison of West African Pollimyrus here. Other non‐type museum specimens were not included, as they first have to be reidentified to a valid or new species, potentially using the identification key for the type specimens provided (see Results). Those two non‐type specimens that were available during the time of the study were not further included, as there was no indication that they might represent novel distinct morphotypes, and thus potentially new species for science, although further study is required.
Limitations
4.4
Working only with the available type specimens in this case resulted in only having data of one or a few specimens for some of the taxa. Most species included in this study were described well over a century ago, often on one or a few type specimens only, and in several cases no material of the same locality is available. The resulting small sizes of the groups complicated, making definitive conclusions based solely on measurement data, as the observed ranges might not reflect the complete variation present in a taxon. The only remedy to this issue is to include more non‐type specimens in follow‐up studies once their species identification has been confirmed using the data presented here. As all species recognized here were differentiated from others by at least two morphological characteristics here, the limited sample sizes should not have impacted the results in a significant way. For those taxa, where there are only a few diagnostic or somewhat overlapping measurement ranges available, the remaining uncertainties were highlighted.
As this study only focused on available type specimens and did not include a molecular approach, the need for further consolidation of the present decisions and conclusions remains. Previous studies have shown that genetic analyses are highly needed to resolve species delineations, and cryptic species occur in this family (e.g., Rich et al., 2017; Sullivan et al., 2016). However, due to the complexity exactly, it remains pertinent to not lose track of morphological and geographical data either to validate a genetic specimen's taxon or to use all information available to identify and confirm identifications. In this study, several changes in the taxonomic status of species and some remaining issues and questions have been uncovered. Although these await confirmation and further study, respectively, with genetic analyses, the morphological synthesis provided here should be taken into consideration when working with Pollimyrus specimens.
With the present study mostly focusing on type specimens to delineate species and describe new species for science, more alpha‐taxonomical issues might be identified and in need to be solved when dealing with non‐type specimens. It is therefore recommended that any specimen currently available in collections and newly collected specimens should be (re‐)examined using the identification key. Only then will it be possible to better evaluate the geographical distribution of the 20 valid Pollimyrus species identified in the present paper. This will then also provide the much‐needed data to better evaluate the IUCN Red List conservation status and open the way to extend insights into their evolution and ecology.
Biogeography of the Pollimyrus species diversity
4.5
Based on the current study of most available type specimens, a first overview of the biogeography of the known Pollimyrus species has been summarized (Table 8). This overview is only preliminary and needs more verified non‐type specimen distribution data. Nevertheless, it shows that, with 11 species, the highest species diversity in Pollimyrus can be found in the Congo IP, whereas only a few species are found in the Nilo‐Sudan (two or three), Lower Guinea (two or three), Zambezi (three; but see Schedel et al., 2024 for a potential new species), East Coast (one), and Great Lakes (one) IPs. Except for their absence from the Angolan and Ethiopian Rift valley IPs, this known distribution for Pollimyrus coincides with that of the family Mormyridae as a whole (Lévêque & Paugy, 2017). Noticeable is also the dominance of slender‐tailed species in the Congolese IP, with seven(eight) slender‐tailed versus two(three) thick‐tailed species. In addition, slender‐tailed species only occur in the Congo IP, except for P. adspersus, whose original collecting locality remains unclear (Günther, 1866). The precise type locality of P. adspersus is unknown and therefore placed, tentatively, in three different IPs in western Africa. On the contrary, thick‐tailed species occur in most IPs from which Pollimyrus species have been reported, but only in low numbers.
In some species complexes and morphological groups, the lack of a clear morphological distinction between type specimens of nominal species complicated the species delineations. However, in some cases, multivariate statistical analysis (PCAs) detected subtle differences in morphology between (hydro)geographically distinctly occurring specimens. This was observed, for instance, in the P. pulverulentus, the P. tumifrons, and the P. stappersii/P. castelnaui species complexes (see Results). These examples show that the Pollimyrus alpha‐taxonomy might be strongly linked to allopatry, although, at present, the distribution ranges of several species in the Congo basin seem to overlap, such as those of P. tumifrons, P. pulverulentus, and P. schreyeni (Figure 2).
As this summary was compiled using mostly data from type specimens only and, as a result, does not take into account the full geographical distribution of the species identified, it could be possible that more species co‐occur in some of these IPs. So far, this is unknown, and each species seems to be restricted to one IP due to the low number of specimens available. Remarkably, P. krameri is the first Pollimyrus species described from the East Coast IP. So far, only P. nigricans is known to occur in one of the great lake basins in Africa, namely the mouth of the Katonga River at Lake Victoria (Great Lakes IP). As the type specimens from P. isidori and P. nigricans are morphologically similar and were found in the Nile River and Lake Victoria basin, respectively, two systems that are hydrologically connected, although the Ripon Falls and Owen Falls separate the Lake from the Nile basin (with presently the Nalubaale Dam), both these species could be phylogenetically closely related. Junior synonyms of P. isidori have been described from several river basins, P. isidori gaillardi from Lake Chad (Nilo‐Sudan IP), P. isidori rudebeckii from the Gambia River (Nilo‐Sudan IP), and P. isidori vanderbilti from the Central African Republic (Nilo‐Sudan/Congo IP). However, these junior synonyms were not included in the present morphological study, as the main scope of this study was to work with the valid species. Therefore, it remains unclear whether these junior synonyms are indeed conspecific with P. isidori. If correct, that would mean that the geographic range of P. isidori is the largest known range for a Pollimyrus species so far, and that one species can occur in multiple river basins and IPs. However, further study is needed to assess the conspecificity and re‐evaluate the distribution of these nominal species. It is to be noted that other nominal species once thought to be junior synonyms of P. isidori occurring in different IPs are now recognized as valid species, that is, P. fasciaticeps and P. osborni from the Congo IP.
The little amount of data on the distribution of species, combined with the small number of specimens known per species, which often come from the same river(basin), might indicate that Pollimyrus species, in general, are not very abundant. Due to their restricted distribution, they might be more vulnerable than more common and widespread species. However, for most species (15), the IUCN Red List (IUCN, 2024) categorized them as Least Concern (LC) (P. adspersus, P. brevis, P. castelnaui, P. isidori [including P. osborni and P. fasciaticeps as subspecies], P. marianne, P. nigricans, P. nigripinnis, P. petricolus, P. pulverulentus, P. schreyeni, P. stappersii, P. tumifrons, and C. plagiostoma), as there is little indication for any threats or population declines (Diouf, 2020; FishBase team RMCA and Geelhand, 2016; Moelants, 2010a, 2010b, 2010c, 2010d, 2010e, 2010f, 2010g, 2010h; Olaosebikan & Lalèyè, 2020; Olaosebikan & Moelants, 2020; Tweddle & Marshall, 2007; Tweddle et al., 2019), whereas three others remain Data Deficient (DD) (P. pedunculatus, P. guttatus, and P. maculipinnis: see Moelants, 2010i, 2010j, 2010k). The four new species were not assessed yet, but there are indications for severe environmental impacts on the type localities of P. krameri and P. weyli (R.B. personal communication). The available assessments, however, consider the range of the species based on identifications of non‐types (e.g., see presence of P. adspersus in the Congo basin), which are, most likely, considering our presented results, not using the correct species identifications (also see Hopkins et al., 2007). As such, the current assessments might not be reflective of the species vulnerability (see also Palacio et al., 2023). Similarly, the distribution details provided for each species on FishBase (Froese & Pauly, 2024) have been compiled from various existing publications. However, considering the numerous identification issues identified, the former distribution data should be handled with care. A revision of their current conservation status is certainly needed, which will only be possible after reidentifying museum specimens and summarizing the new resulting distribution for each of these.
Precarious mormyrid taxonomic knowledge: Pollimyrus as an illustrative case study
4.6
The Mormyridae, endemic to Africa and with 227 valid species known today (Fricke et al., 2024), is one of its important fish diversity components. Nevertheless, even during the past decade, new mormyrid species have been described on a regular basis, for example, Petrocephalus petersi (Kramer et al., 2012), Pollimyrus cuandoensis (Kramer et al., 2013), Marcusenius kaninginii (Kisekelwa et al., 2016), Paramormyrops ntotom (Rich et al., 2017), Marcusenius wamuinii (Decru et al., 2019), Marcusenius verheyenorum (Mambo et al., 2019), and Cyphomyrus lufirae (Mukweze Mulelenu et al., 2020). This, however, contrasts with our current overall alpha‐taxonomical, ecological, and other knowledge about mormyrid species diversity. For many taxa, both genera and species, a taxonomic revision is still lacking, genetic sampling has been limited, and ecological and behavioral, including EOD, information is often unavailable. The genus Pollimyrus and its species are no exception to this observation.
Taxonomic studies form the basis for many other kinds of knowledge, whether genetic, behavioral, evolutionary, ecological, etc., which could be combined in integrative studies. This makes these kinds of alpha‐taxonomic syntheses, even if based on type specimens alone, crucial for any further and future research on Mormyridae. The lack of alpha‐taxonomic knowledge and insights in the species distribution and ecology also makes it more difficult to effectively protect these species. Indeed, conservation actions are only possible based on sound knowledge of a species and its distribution. As this synthesis of Pollimyrus has also shown, the issue of generic delineations can still be problematic in Mormyridae, with several species having been the topic of reallocations in the past and present study. Furthermore, many unresolved issues, due to a lack of available non‐type specimens from certain type localities and regions, hinder the precise species delineations and the (re)evaluation of the status of junior synonyms and (sub)species. Therefore, Pollimyrus, with its now 20 recognized valid species, can serve as an example for the alpha‐taxonomic and generic issues in several mormyrid genera occurring across the African continent. Thus, research on much more type specimens is needed to come to a better overview of the alpha‐taxonomy of the more than 227 valid species, currently distributed across 22 genera (Fricke et al., 2024). As illustrated by the present revision of species within the genus Pollimyrus, many more hidden new species are probably to be discovered as well.
CONCLUSIONS
5
Pollimyrus is a species‐rich mormyrid genus with a known distribution across six ichthyogeographical provinces. A morphometric synthesis of all species in this genus resulted in (i) the confirmation of C. plagiostoma to Cyphomyrus and reallocation of P. eburneensis to Paramormyrops, (ii) the confirmation of P. guttatus, previously assigned to Petrocephalus, within Pollimyrus, (iii) a re‐evaluation of the generic characters and species status of all nominal species of the genus Pollimyrus, and (iv) the description of four new species for science, resulting in the identification of a total of 20 valid species in this genus. As this study allowed for a better understanding of the generic and morphometric species delineations, additional follow‐up studies will be able to provide a more profound understanding of the genetic diversity, geographic distribution, ecology, and behavior of these species. Furthermore, this synthesis has highlighted that certain (sub)basins, such as those in West Africa, situated within the known distribution area of the genus, remain highly understudied. If considering this case study on Pollimyrus as an illustrative example with broader implications for taxonomic diversity patterns within Mormyridae, it suggests that the species richness within this family is most likely still underestimated despite the significant taxonomic work conducted in certain genera and geographical regions, as several other mormyrid genera are still awaiting a similar, comprehensive synthesis.
AUTHOR CONTRIBUTIONS
Katrien Dierickx: conceptualization, formal analysis, investigation, writing—original draft, writing—review and editing, visualization. Soleil Wamuini Lunkayilakio: resources, writing—review and editing. Roger Bills: resources, writing—review and editing. Emmanuel Vreven: conceptualization, writing—review and editing, supervision.
Supporting information
File S1. Figures and additional results.
File S2. Raw data tables.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bass, A. H. , & Hopkins, C. D. (1985). Hormonal control of sex differences in the electric organ discharge (EOD) of mormyrid fishes. Journal of Comparative Physiology, 156(4600), 587–604. 10.1126/science.6844924 · doi ↗
- 2Bell‐Cross G. 1976. The fishes of Rhodesia. Trustees of the National Museums and Monuments of Rhodesia.
- 3Bigorne, R. (1990). Révision systématique du genre Pollimyrus (Teleostei: Mormyridae) en Afrique de l'ouest. Revue d'Hydrobiologie Tropicale, 23(4), 313–327.
- 4Bigorne, R. (2003). Mormyridae. In D. Paugy , C. Lévêque , & G. G. Teugels (Eds.), The fresh and brackish water fishes of west Africa: 122–184. Muséum national d'Histoire naturelle.
- 5Boden, G. , Teugels, G. G. , & Hopkins, C. D. (1997). A systematic revision of the large scaled Marcusenius with description of a new species from Cameroon (Teleostei; Osteoglossomorpha; Mormyridae). Journal of Natural History, 31(11), 1645–1682. 10.1080/00222939700770881 · doi ↗
- 6Boulenger, G. A. (1899). Matériaux pour la faune du Congo. Zoologie.–Série I. Poissons nouveaux du Congo. Quatrième Partie. Polyptères, Clupes, Mormyres, Characins. Annales du Musee du Congo (Ser. Zoology), 1, 59–96.
- 7Boulenger, G. A. (1911). On a collection of fishes from the Lake Ngami Basin, Bechuanaland. Transactions of the Zoological Society of London, 18, 399–431.
- 8Boulenger, G. A. (1915). Diagnoses de poissons nouveaux. II. Mormyrides, Kneriides, Characinides, Cyprinides, Silurides. (Mission Stappers au Tanganika‐Moero.). Revue de Zoologie Africaine, 4, 162–171.