Endothelial deletion of adipose triglyceride lipase protects against heart failure with preserved ejection fraction
Juliane Schwanbeck, Max Stahnke, Anna Eberlein, Madeleine Goeritzer, Arndt Schulze, Dominique Pernitsch, Dagmar Kolb, Gernot F. Grabner, Theda U.P. Bartolomaeus, Sofia K. Forslund, Holger Gerhardt, Gabriele G. Schiattarella, Lucia Cocera Ortega, Natalia López-Anguita

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
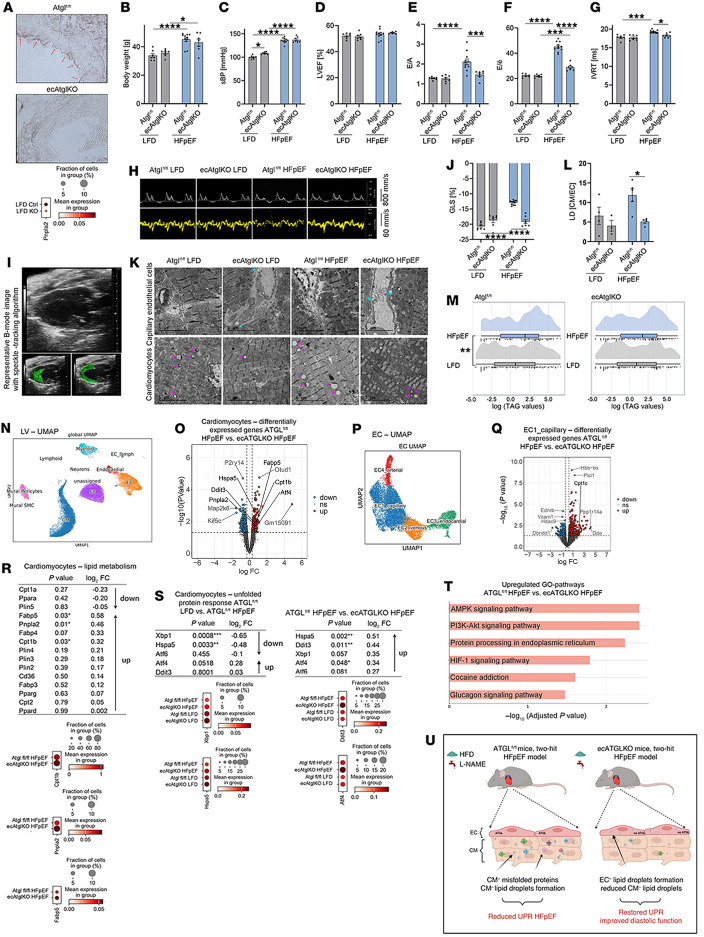 Figure 1
Figure 1- —Einstein Foundation/ Foundation Charité
- —Deutsche Forschungsgemeinschafthttps://doi.org/10.13039/501100001659
- —Deutsches Zentrum für Herz-Kreislaufforschunghttps://doi.org/10.13039/100010447
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAdipose Tissue and Metabolism · Lipid metabolism and biosynthesis · Diabetes, Cardiovascular Risks, and Lipoproteins
To the Editor:
Heart failure with preserved ejection fraction (HFpEF) pathophysiology is multifactorial, with alterations in cardiac lipid metabolism likely playing a central role (1). Cardiac lipid metabolism is governed by transcellular uptake and transport of circulating lipids across cardiac endothelial cells (EC). In HFpEF, however, the processes of endothelial lipid handling and their functional relevance for lipid handling in the myocardium are largely unknown. Lipid processing in ECs relies on the function of the lipid droplet–hydrolyzing (LD-hydrolyzing) enzyme adipose triglyceride lipase (ATGL), the deletion or pharmacological blockade of which results in the accumulation of LDs in ECs (2–5).
EC-specific Atgl-KO mice (ecAtglKO) and their Cre WT littermate controls (Atgl^fl/fl^) (Figure 1A) were subjected to the 2-hit (high-fat diet [HFD]/L-NAME) HFpEF protocol (Figure 1, B and C) (6). Echocardiography after 15 weeks revealed a preserved left ventricular (LV) ejection fraction (EF) and LV hypertrophy in both genotypes (Figure 1D and Supplemental Figure 1, A–C; supplemental material available online with this article; https://doi.org/10.1172/jci.insight.187145DS1). Surprisingly, ecAtglKO mice were protected against HFD/L-NAME–mediated diastolic dysfunction (Figure 1, E–H) and exhibited improved global longitudinal strain (GLS) (Figure 1, I and J, and Supplemental Figure 1, D and E).
HFpEF was associated with enhanced LD formation in cardiomyocytes with few LDs detected in capillary ECs (Figure 1, K and L). Endothelial ATGL deletion led to enhanced LD formation in ECs and a reduced LD cardiomyocyte/EC ratio (Figure 1, K and L). LV triacylglycerol (TAG) accumulation significantly increased in Atgl^fl/fl^-HFpEF mice but not in ecAtglKO-HFpEF mice (Figure 1M and Supplemental Figure 1F). Distinct differences in mitochondrial or sarcomeric structure were not detected between Atgl^fl/fl^-HFpEF and ecAtglKO-HFpEF mice (data not shown).
We next performed single nuclei RNA-Seq from LV samples (Figure 1, N–Q, and Supplemental Figure 2, A and B). Comparison of Atgl^fl/fl^-HFpEF and ecAtglKO-HFpEF mice revealed 264 genes significantly upregulated and 261 downregulated in cardiomyocytes as well as 475 upregulated and 165 downregulated genes in capillary ECs (Figure 1, O and Q). We could not detect any clear inflammatory response in EC subclusters (Supplemental Figure 2, C and D, and Supplemental Figure 3). In cardiomyocytes from ecAtglKO-HFpEF mice, genes involved in FA metabolism were significantly upregulated (Figure 1R). Dysregulation of the unfolded protein response (UPR) has been recently identified as a pathogenic driver of HFpEF connected to lipid metabolism (6). Similarly, we found a significant reduction of cardiomyocyte genes involved in the IRE1α/XBP1 signaling pathway of the UPR in Atgl^fl/fl^-HFpEF mice (Figure 1S, left), accompanied by a reduction of IREα phosphorylation (Supplemental Figure 4A) (6). This reduction was notably absent in ecAtglKO-HFpEF mice, and genes involved in protein processing in the ER were upregulated (Figure 1, S [right] and T, and Supplemental Figure 4A). Finally, pharmacological inhibition of ATGL in ECs resulted in increased LD formation in ECs and reduced LD detection in cardiomyocytes, associated with increased expression of Hspa5/BiP in cardiomyocytes (Supplemental Figure 4, B–D).
In conclusion, endothelial-specific deletion of ATGL improved diastolic function in HFpEF accompanied by changes of neutral lipid storage at the capillary EC–cardiomyocyte interface. Mechanistically, reduced LD accumulation in cardiomyocytes may reverse the suppression of the IRE1α/XBP1 axis of the UPR (Figure 1U) (6). In addition to the effects observed at the capillary EC–cardiomyocyte interface, systemic metabolic effects may have contributed to the HFpEF improvement in ecAtglKO mice. Recent publications demonstrate that endothelial ATGL deletion promotes endothelial dysfunction, arterial hypertension, and atherosclerosis mediated by the suppression of the eNOS/NO pathway (2–5). The unique property of our model is that this pathway was continuously blocked by L-NAME, which likely prevented the detection of detrimental effects of LD formation on NO-dependent function and contributed to the phenotype observed in ecAtglKO-HFpEF mice. Finally, it is important to note that the 2-hit HFpEF model used here also has limitations and that an analysis of endothelial ATGL in other HFpEF models would be recommended.
Supplementary Material
Supplemental data
Supporting data values
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mishra S Kass DA Cellular and molecular pathobiology of heart failure with preserved ejection fraction Nat Rev Cardiol 202118640042310.1038/s 41569-020-00480-633432192 PMC 8574228 · doi ↗ · pubmed ↗
- 2Boutagy NE et al Dynamic metabolism of endothelial triglycerides protects against atherosclerosis in mice J Clin Invest 20241344 e 17045310.1172/JCI 17045338175710 PMC 10866653 · doi ↗ · pubmed ↗
- 3Kim B et al Endothelial lipid droplets suppress e NOS to link high fat consumption to blood pressure elevation J Clin Invest 202313324 e 17316010.1172/JCI 17316037824206 PMC 10721151 · doi ↗ · pubmed ↗
- 4Kuo A et al Lipid droplet biogenesis and function in the endothelium Circ Res 201712081289129710.1161/CIRCRESAHA.116.31049828119423 PMC 5392152 · doi ↗ · pubmed ↗
- 5Sternak M et al Vascular ATGL-dependent lipolysis and the activation of c PLA 2-PGI 2 pathway protect against postprandial endothelial dysfunction Cell Mol Life Sci 202481112510.1007/s 00018-024-05167-638467757 PMC 10927860 · doi ↗ · pubmed ↗
- 6Schiattarella GG et al Nitrosative stress drives heart failure with preserved ejection fraction Nature 2019568775235135610.1038/s 41586-019-1100-z 30971818 PMC 6635957 · doi ↗ · pubmed ↗