Three new species of the cockroach genus Nocticola Bolívar, 1892 (Blattodea, Corydioidea, Nocticolidae) from China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6 Figure 7
Figure 7 Figure 8
Figure 8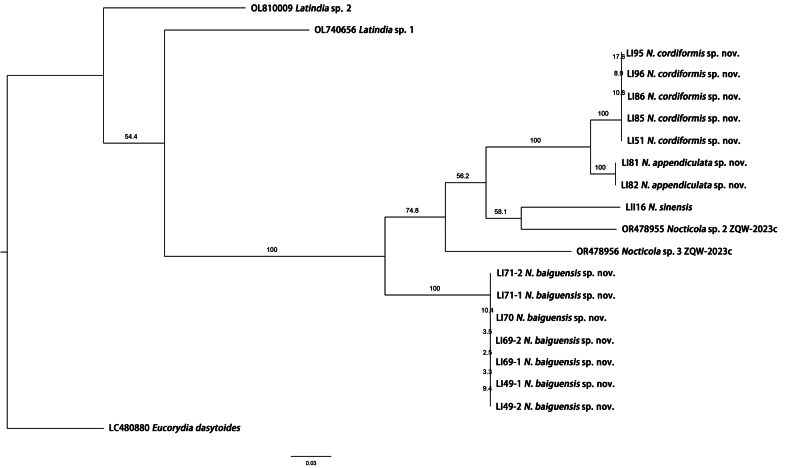 Figure 9
Figure 9| Species | Voucher number | Location | GenBank number |
|---|---|---|---|
| LI49-1 | Yanshan District, Guilin City, Guangxi Province |
| |
| LI49-2 |
| ||
| LI69-1 | Xiangshan District, Guilin City, Guangxi Province, |
| |
| LI69-2 |
| ||
| LI70 | Diecai District, Guilin City, Guangxi Province, |
| |
| LI71-1 |
| ||
| LI71-2 |
| ||
| LI51 | Lingui District, Guilin City, Guangxi Province |
| |
| LI85 |
| ||
| LI86 |
| ||
| LI95 |
| ||
| LI96 |
| ||
| LI81 | Lingchuan County, Guilin City, Guangxi Province, |
| |
| LI82 | Lingui District, Guilin City, Guangxi Province |
| |
|
| LII16 | Jiangyong County, Yongzhou City, Hunan Province |
|
| / | /China |
| |
| / | /China |
| |
| B467 | French Guiana |
| |
| B481 | French Guiana |
| |
|
| / | Taiwan, China |
|
| 1 | Fourth abdominal tergum specialized, with dense setae on median area |
|
| – | Fourth abdominal tergum not specialized |
|
| 2 | Tegmina not extending beyond the end of abdomen |
|
| – | Tegmina extending beyond the end of abdomen |
|
| 3 | Tegmina and wings almost equal in length |
|
| – | Tegmina developed; hind wings reduced |
|
| 4 | Subgenital plate weakly asymmetrical; accessory hook-like phallomere (L4N) parabola-like |
|
| – | Subgenital plate symmetrical; accessory hook-like phallomere (L4N) fin-shaped |
|
| Specimen | LI49-1 | LI49-2 | LI69-1 | LI69-2 | LI70 | LI70-1 | LI70-2 | LI51 | LI85 | LI86 | LI95 | LI96 | LI81 | LI82 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LI49-1 | 0 | 0 | 0 | 0 | 0 | 0 | 112 | 112 | 112 | 112 | 112 | 120 | 120 | |
| LI49-2 | 0.0000 | 0 | 0 | 0 | 0 | 0 | 112 | 112 | 112 | 112 | 112 | 120 | 120 | |
| LI69-1 | 0.0000 | 0.0000 | 0 | 0 | 0 | 0 | 112 | 112 | 112 | 112 | 112 | 120 | 120 | |
| LI69-2 | 0.0000 | 0.0000 | 0.0000 | 0 | 0 | 0 | 112 | 112 | 112 | 112 | 112 | 120 | 120 | |
| LI70 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0 | 0 | 112 | 112 | 112 | 112 | 112 | 120 | 120 | |
| LI70-1 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0 | 112 | 112 | 112 | 112 | 112 | 120 | 120 | |
| LI70-2 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 112 | 112 | 112 | 112 | 112 | 120 | 120 | |
| LI51 | 0.1940 | 0.1940 | 0.1940 | 0.1940 | 0.1940 | 0.1940 | 0.1940 | 0 | 0 | 0 | 0 | 27 | 27 | |
| LI85 | 0.1940 | 0.1940 | 0.1940 | 0.1940 | 0.1940 | 0.1940 | 0.1940 | 0.0000 | 0 | 0 | 0 | 27 | 27 | |
| LI86 | 0.1940 | 0.1940 | 0.1940 | 0.1940 | 0.1940 | 0.1940 | 0.1940 | 0.0000 | 0.0000 | 0 | 0 | 27 | 27 | |
| LI95 | 0.1940 | 0.1940 | 0.1940 | 0.1940 | 0.1940 | 0.1940 | 0.1940 | 0.0000 | 0.0000 | 0.0000 | 0 | 27 | 27 | |
| LI96 | 0.1940 | 0.1940 | 0.1940 | 0.1940 | 0.1940 | 0.1940 | 0.1940 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 27 | 27 | |
| LI81 | 0.2105 | 0.2105 | 0.2105 | 0.2105 | 0.2105 | 0.2105 | 0.2105 | 0.0426 | 0.0426 | 0.0426 | 0.0426 | 0.0426 | 0 | |
| LI82 | 0.2105 | 0.2105 | 0.2105 | 0.2105 | 0.2105 | 0.2105 | 0.2105 | 0.0426 | 0.0426 | 0.0426 | 0.0426 | 0.0426 | 0.0000 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant and animal studies · Fossil Insects in Amber · Insect and Arachnid Ecology and Behavior
Introduction
The genus Nocticola, with the type species Nocticolasimoni Bolívar, was established by Bolívar (1892), who also described N.caeca Bolívar, 1892 in the same year; the first family-group name based on Nocticola is Nocticolinae (Bolívar 1892). Nocticolidae was first used by Bruner (1915) for two species described by Silvestri (1946), N.sinensis Silvestri and N.termitophila Silvestri, with N.sinensis being the earliest recorded Nocticola in China. Currently 29 known Nocticola species have been described all over the world (Beccaloni 2014; Trotter et al. 2017; Liu et al. 2017). They are very small and delicate, with reduced male wing veins. Among them, 17 species are cavernicolous, eight epigean, and four termitophilous (Fernando 1957; Roth 1988, 1991, 1995, 2003a; Roth and McGavin 1994; Andersen and Kjaerandsen 1995; Lucañas and Lit 2016; Lucañas et al. 2021; Lucañas and Maosheng 2023; Liu et al. 2017; Trotter et al. 2017). Handlirsch (1925) once classified Nocticola as Blattoidea based on the characteristics of the ovipositor valves in the seventh abdominal plate of Nocticola females, and subsequently, Princis (1966) listed Nocticolidae as a branch of Blattoidea. Roth (1988) suggested that Nocticolidae could be placed between Polyphagidae and Blattoidea. In recent years, molecular analysis has recovered Nocticolidae as sister to Corydiidae, particularly Latindiinae (Inward et al. 2007, Murienne 2009; Djernæs et al. 2012), supporting the position of Nocticolidae and Latindiinae as sister groups, and infer that Nocticolidae may be a specialized form of Latindiinae (Djernæs et al. 2015; Legendre et al. 2015; Li and Huang 2020). Subsequently, Han et al. (2023) assess the phylogenetic relationships of 35 Corydioidea species with mitochondrial genomes and two nuclear gene fragments, and they infer that the Latindiinae belong to the family Nocticolidae.
Materials and methods
Morphological study
Specimens were collected in Guangxi Province from 2023–2024. The examined material of N.xiai Liu, Zhu, Dai & Wang, 2017 (Fig. 1A–C) is deposited in the Institute of Entomology of the Shanghai Entomological Museum, Chinese Academy of Sciences, Shanghai (SHEM). The type specimens of the three new species and the examined specimens of N.sinensis Silvestri, 1946 (Fig. 2A–L) were deposited in the Zhongshan Customs Technology Center (ZSCTC).
Nocticolaxiai Liu, Zhu, Dai & Wang, 2017: A holotype male, lateral B head C phallomeres (photographs provided by Han-Qiang Wang, SHEM). Abbreviations: L1, L2, L3, L4G, L4N, L4M: sclerites of the left phallomere (L1 situated in the central dorsal wall; L2 arch-shaped sclerite situated in the ventral to L1; L3 situated in the left wall protrudes a large hook-process; L4G situated in the posterior ventral wall of the ventral lobe; L4N accessory hook-like phallomere; L4M situated in the ventral wall); R1, R3: sclerites of the left phallomere (R1 situated in the dorsal or ventral walls, or in the posterior part of dorsal and ventral wall; R3 plate-like situated in the anteriormost ventral wall).
Nocticolasinensis Silvestri, 1946: A male, dorsal view B male, ventral view C pronotum D head E fore femur F tegmen G hind wings H tarsal claw I hind tarsus J supra-anal plate, dorsal view; male genitalia K phallomeres L subgenital plate, ventral view. Abbreviations: L1, L2, L3, L4G, L4N, L4M sclerites of the left phallomere (L1 situated in the central dorsal wall; L2 arch-shaped sclerite situated in the ventral to L1; L3 situated in the left wall protrudes a large hook-process; L4G situated in the posterior ventral wall of the ventral lobe; L4N accessory hook-like phallomere; L4M situated in the ventral wall); R1, R2, R3, R1H sclerites of the left phallomere (R1 situated in the dorsal or ventral walls, or in the posterior part of dorsal and ventral wall; R2 a ridge on the ventral margin; R3 plate-like situated in the anteriormost ventral wall; R1H a larger lobed situated in the dorsal wall, with extensions into the ventral wall).
The lateral tergum behind the seventh abdominal tergum (T7) of the male specimen was cut off, placed into a 1.5 ml centrifuge tube with 10% NaOH and digested at 70 °C for 30–45 min. After digestion, the NaOH was removed from the centrifuge tube, and the specimen was rinsed thrice with water before examination. The specimens were dissected and observed under a ZEISS Discovery V12 stereo microscope. Photographs were taken with a ZEISS/Smart Zoom5 and Canon EOS 5D Mark III, and illustrated with Adobe Photoshop 2022 software. After illustration, the genitalia were stored in 0.5 ml centrifuge tubes containing 50% glycerol, and preserved with the remainder of the specimen, which was stored in ethyl alcohol. Terminologies used for male genitalia follow Klass (1997) and for other characters follow Roth (2003b).
Molecular biology study
DNA was obtained from hind tarsus of adult and nymph cockroaches using TIANamp Genomic DNA Kit produced by Tiangen. The primers used for PCR amplification were the universal primers for cytochrome C oxidase subunit I (COI) gene: LOC1490 5’-GGTCAACAAATCATAAAGATATTGG-3’, HCO2198 5’-TAAACTTCAGGGTGACCAAAAAATCA-3’ (Folmer et al. 1994). The reagents for primers synthesized, EX-Taq DNA polymerase and dNTP were purchased from Takara Biotechnology (Dalian) Co., Ltd. The amplification conditions were denaturation at 95 °C for 5 min; 95 °C for 30 s, 50 °C for 30 s, 72 °C for 60 s, 35 cycles; final extension at 72 °C for 10 min, then held at 4 °C. The amplified samples with bands in Gel imaging System were sequenced by Tianyi Huiyuan Gene Technology Co., Ltd. (Guangzhou).
A total of 20 COI sequences were analyzed, of which 15 sequences were obtained in this study and Five sequences were downloaded from GenBank (Yanagisawa et al. 2021; Han et al. 2023; Li et al. 2022). Latindia sp. 1, Latindia sp. 2, and Eucorydiadasytoides were selected as outgroups (Table 1). Unfortunately, molecular data for N.xiai were not obtained, so could not be included in this study. All 15 sequences have been submitted to GenBank (https://www.ncbi.nlm.nih.gov/nuccore) with accession numbers PQ601347–PQ601361. The COI fragments were processed using the MUSCLE algorithm within MEGA 6.0 (Tamura et al. 2013), and a phylogenetic tree was constructed using Maximum Likelihood (ML) tree (Kimura 1980) and visualized using FigTree v. 1.4.4 (http://tree.bio.ed.ac.uk/software/figtree/). Nucleotide sites differences, genetic distances, both interspecific and intraspecific were calculated using the Kimura-2-paramaters (K2P) with 1,000 ultrafast bootstrap replicates (Hoang et al. 2018).
Results
Taxonomy
Family Nocticolidae Bolívar, 1892
Nocticola
Taxon classificationAnimaliaBlattodeaNocticolidae
Genus
Bolívar, 1892
20E84954-832B-51F8-A9F6-F9721D7587E8
Nocticola Bolívar, 1892: 29.
Type species.
Nocticolasimoni. First used as Nocticolidae Brunner 1915. Roth (1988) diagnosed and discussed Nocticolidae.
Diagnosis.
The following description is in accordance with the traits proposed by Roth (1988) and the diagnosis of Andersen and Kjaerandsen (1995). Habitus small and delicate. Eyes well developed, variably reduced or absent; ocelli present or absent. Male wings are either reduced or well developed; front and hind wings are similar if well developed, membranous hyaline with minute pubescence and with few, essentially straight veins. Anteroventral margin of front femur with a row of piliform setae only, terminating at one or more large spines (= Type C); arolia and pulvilli absent; tarsal claws very small, simple, and symmetrical. Left and right phallomeres of the male genitalia are complex, always with a hook. Modification of the male abdomen tergal gland divides Nocticola into two groups: the simoni species group (male terga unspecialized) and the uenoi species group (male fourth abdominal terga specialized).
Differential diagnosis.
Gravely (1910) compared the wing veins of Alluaudellinahimalayensis and Cardacuswilleyi, and concluded that the wing veins cannot be an important feature to distinguish these two genera. He distinguished the two genera by the presence or absence of ocelli, and whether the head is exposed or covered by the pronotum. Karny (1924) distinguished Cardacopsis, Alluaudellina and Cardacus by the degree of eye development, wing venation, and the presence or absence of ocelli, but he did not include Nocticola in the discussion. Roth (1988) considered that Chopard (1946) established the genus Typhloblattodes using a nymph specimen, questioning the validity of this genus. Chopard (1932, 1946, 1966) believed that there was polymorphism in the wing veins of Alluaudellinahimalayensis, and, in Nocticola, polymorphism in wings and eyes development, making these features of questionable value in generic diagnosis. Consequently, he was unable to distinguish Nocticola from Alluaudellina and believed that they may be synonyms. The genus Nocticola can be distinguished from Typhloblatta and Pholeosilpha by the following characteristics: anteroventral margin of front femur Type C, in contrast forefemoral spination type B2 in Typhloblatta and Pholeosilpha. The genus Nocticola can be distinguished from Spelaeoblatta by the following characteristics: 1) anteroventral margin of front femur Type C, in contrast fore femoral spination type B1 in Spelaeoblatta; 2) abdominal terga are unspecialized or have a gland on the fourth segment, while in Spelaeoblatta tergal glands on the second and third abdominal tergum; 3) female apterous, whereas female of Spelaeoblatta lack hind wings but have reduced lateral tegminal pads; 4) male tegmina membranous, with distinctive venation, while in Spelaeoblatta tegmina corneous, with poorly defined veins. The genus Nocticola can be distinguished from Helmablatta by the following characteristics: 1) anteroventral margin of front femur Type C, in contrast fore femoral spination type intermediate between A1 and B1 in Helmablatta; 2) abdominal terga are unspecialized or have a gland on the fourth segment, while in Helmablatta third to fifth abdominal tergum form a composite gland, and eighth abdominal tergum is specialized. The genus Nocticola can be distinguished from Metanocticola by the following characteristics: abdominal terga are unspecialized or have a gland on the fourth segment, while in Metanocticola male has a sex gland on the metanotum. According to current molecular analysis research, it has been shown that Nocticola is polyphyletic (Kovacs et al. 2024). The three new species are placed in the genus Nocticola based on features of the eyes, male wings, anteroventral margin of front femur type, arolia, pulvilli, abdomen tergal, and male genitalia.
Key to species of Nocticola from China (males)
**: **
Nocticola
sinensis
Taxon classificationAnimaliaBlattodeaNocticolidae
Silvestri, 1946
7E1E7D49-BFD5-55CE-8C30-6F1501954424
Nocticola sinensis Silvestri, 1946: 329; Silvestri 1947: 15; Princis 1952: 43; Princis 1966: 603; Roth 1988: 299; Liu et al. 2017.
Material examined.
China • 1 ♂ (deposited in SHEM); Hong Kong • 1 ♂, (deposited in ZSCTC), Hunan Province, Yongzhou City, Jiangyong County; 25°20.51'N, 111°20.34'E; 330 m; 14 July 2024, Hao-fei Fan leg.
Description.
Small size, adult yellowish. Male. (Fig. 2A, B). Head: vertex of head exposed (Fig. 2C); eyes reduced to a few ommatidia narrowly grouped near antennal sockets (Fig. 2D); ocelli absent. Pronotum suboval, middle of posterior margin with weak invagination, anterior margin, and lateral margin with 12 setae (Fig. 2C). Tegmina and hind wings: tegmina not extending beyond the end of abdomen (Fig. 2A), veins reduced in number, densely pubescent (Fig. 2F). Hind wings reduced, veins reduced in number (Fig. 2G). Legs: legs long and slender. Anteroventral margin of front femur Type C1 (Fig. 2E); the first tarsus of the hind leg longer than the sum of the remaining tarsi; tarsal claws symmetrical and unspecialized (Fig. 2H), arolium and pulvillus absent (Fig. 2I). Abdomen and genitalia: abdominal tergal gland unspecialized. Supra-anal plate symmetrical, middle of the hind margin triangular concave. Cerci 10 segments; ventral surface of segments not spinous setae (Fig. 2J). Subgenital symmetrical, middle of the hind margin concave (Fig. 2L). Style absent. Male genitalia: genital hook (L3) elongate, ventral to hook with approximately 14 strong setae; accessory hook-like phallomere (L4N), curved inward to approach a right angle, heavily sclerotized; L2 narrow; left process of L1 extends downward, and the right process curved inward; R1 membranous; R2 surface with scale-like tubercles; R3 membranous; R1H reduced, rounded margin (Fig. 2K).
Measurements (mm).
Male, pronotum: length × width: 0.76–0.90 × 0.98–1.20, tegmen: 1.73–2.00, body length (the length from the tip of vertex up to the tip of abdomen): 2.70–3.00;
Distribution.
China (Hong Kong, Hunan Prov.).
Nocticola
baiguensis
sp. nov.
Taxon classificationAnimaliaBlattodeaNocticolidae
CA279ABF-1D85-553A-8D9A-532731C3FF62
https://zoobank.org/63BC7718-864D-4327-84F6-9433E063A45D
Type material.
Holotype: China • 1 ♂; Guangxi Province, Guilin City, Yanshan District, Wangjia Village, Baigu Cave; 25°13.85'N, 110°20.52'E; 162 m; 1 November 2023, Hao-fei Fan leg; ZSCTC-LI0001. Paratype: China • 9 ♂; same data as for holotype; 14 March 2024, Ting-Ting Li leg; ZSCTC-LI-0002- LI-00010 • 8 ♀; same collection data as for preceding; 14 March 2024, De-Xing Liu leg; ZSCTC-LI-00011- LI-00018 • 6 nymphs; same collection data as for preceding; 14 March 2024, Ting-Ting Li leg; ZSCTC-LI-00019- LI-00024 • 1 ♂; Guangxi Province, Guilin City, Xiangshan District, Guilin National Forest Park in Guanxi; 25°13.93'N, 110°13.92'E; 148 m; 18 March 2024, Hao-fei Fan leg; ZSCTC-LI-0025 • 1 ♂; Guangxi Province, Guilin City, Diecai District, Baiyun Temple, 25°18.75'N, 110°22.38'E; 526 m; 28 March 2024, Hao-fei Fan leg; ZSCTC-LI-0026 • 1 ♀; same collection data as for preceding; 28 March 2024, Hao-fei Fan leg; ZSCTC-LI-0027
Diagnosis.
The absence of a specialized abdominal tergal gland places the new species into the simoni species group. This species is similar to N.baumi Lucañas, Blaha, Rahmadi & Patoka, 2021, N.bolivari Chopard, 1950, N.brooksi Roth, 1995, N.cockingi Trotter, McRae, Main & Finston, 2017, N.gonzalezi Lucañas & Lit, 2016, N.leleupi Chopard, 1966, N.quartermaieni Trotter, McRae, Main & Finston, 2017, and N.termitophila Silvestri, 1946 as all these species are apterous. It can be distinguished by its eyes reduced to a few ommatidia narrowly grouped near antennal sockets (Fig. 3D), while in N.baumi, N.bolivari, and N.cockingi eyes are absent. In addition, the tegmina of this species are distinctly longer than the end of the abdomen, while in N.brooksi, N.gonzalezi, N.leleupi, N.quartermaieni, and N.termitophila tegmina do not extend beyond the end of abdomen.
Nocticolabaiguensis sp. nov.: adult male A dorsal view B ventral view; adult female C dorsal view D ventral view E male pronotum F male head G forefemur H tegmen I hindwings J supra-anal plate, ventral view K subgenital plate, ventral view L tarsal claw N hind tarsus; male genitalia M phallomeres. Abbreviations: L1, L2, L3, L4G, L4N, L4M: sclerites of the left phallomere (L1 situated in the central dorsal wall; L2 arch-shaped sclerite situated in the ventral to L1; L3 situated in the left wall protrudes a large hook-process; L4G situated in the posterior ventral wall of the ventral lobe; L4N accessory hook-like phallomere; L4M situated in the ventral wall); R1, R2, R3, R1H: sclerites of the left phallomere (R1 situated in the dorsal or ventral walls, or in the posterior part of dorsal and ventral wall; R2 a ridge on the ventral margin; R3 plate-like situated in the anteriormost ventral wall; R1H a larger lobed situated in the dorsal wall, with extensions into the ventral wall).
Measurements (mm).
Male, pronotum: length × width: 0.80–1.01 × 1.09–1.30, tegmen: 2.29–2.32, overall length (including tegmen): 2.99–3.02, body length (the length from the tip of vertex up to the tip of abdomen): 2.85–3.35. Female, pronotum: length × width: 0.96–1.15 × 1.30–1.51, body length (the length from the tip of vertex up to the tip of abdomen): 3.16–3.67.
Description.
Small size. Nymphs whitish (Fig. 4F), adult yellowish. Male. (Figs 3A, B, 4E). Head: vertex of head exposed; eyes reduced to a few ommatidia narrowly grouped near antennal sockets (Fig. 3F); ocelli absent (Fig. 3F). Pronotum suboval, both sides of posterior margin with weak invagination, anterior margin and lateral margin with 12 setae (Fig. 3E). Tegmina and hind wings: tegmina extending beyond the end of abdomen (Fig. 3A, B), veins reduced in number, densely pubescent (Fig. 3H). Lateral portions of metanotum pubescent, somewhat produced, suggesting a wing surface, but this region not separated from metanotum. Legs: legs long and slender (Fig. 3I). Anteroventral margin of front femur Type C1 (Fig. 3G); the first tarsus of the hind leg longer than the sum of the remaining tarsi; tarsal claws symmetrical and unspecialized (Fig. 3L), arolium and pulvillus absent (Fig. 3N). Abdomen and genitalia: abdominal tergal gland unspecialized. Supra-anal plate symmetrical, middle of the hind margin triangular concave. Cerci with 11 segments; ventral surface of segments without spinous setae (Fig. 3J). Subgenital symmetrical, middle of the hind margin concave (Fig. 3K). Style absent. Male genitalia: genital hook (L3) elongate, ventral to hook with approximately 17 strong setae; accessory hook-like phallomere (L4N), apex short and with double-hook, heavily sclerotized; L2 narrow, L1 elongate, process long spine-like; R1 membranous, distal capitate-like; R2 sinuate protrusion of central part, surface with scale-like tubercles; R3 membranous; R1H reduced, rounded margin (Fig. 3M).
Female: Apterous (Fig. 4C, D). Supra-anal plate triangular, transverse of hind margin, middle with U-shaped invagination. Subgenital lobate (Fig. 3C, D). Cerci with 11 segments; ventral surface of segments without spinous setae.
Etymology.
The specific name baiguensis is derived from the cave called Baigu Cave (Fig. 4A, B), which is the first collection site of this species.
Habitats of Nocticolabaiguensis sp. nov. from China A, B external environment of the Baigu Cave C, DN.baiguensis sp. nov. found on stone, surrounded by some animal feces EN.baiguensis sp. nov. found on rotting branch F nymphs of Nocticolabaiguensis sp. nov.
Distribution.
China (Guangxi).
Nocticola
cordiformis
sp. nov.
Taxon classificationAnimaliaBlattodeaNocticolidae
FCBA08CF-1C1B-59E3-B5E2-A8D5381EC11A
https://zoobank.org/867C3F19-3F52-4B41-80D9-8ECE341029E7
Type material.
Holotype: China • 1 ♂; Guangxi Province, Guilin City, Lingui District, Huixian Town, Edi Village, Yanzi Cave; 25°3.30'N, 110°11.27'E; 195 m; 13 December 2023; Hao-Fei Fan leg; ZSCTC-LI-0028**. Paratype**: China • 1 ♂; Guangxi Province, Guilin City, Lingui District, Lingjiaodi Village; 25°13.58'N, 110°14.50'E; 148 m; 30 December 2023; Hao-Fei Fan leg; ZSCTC-LI-0029 • 1 ♀; same collection data as for preceding; 30 December 2023; Hao-Fei Fan leg; ZSCTC-LI-0030 • 2 ♂; Guangxi Province, Guilin City, Lingui District, Huixian Town, Edi Village, 25°3.07'N, 110°10.25'E; 176 m; 29 April 2024; Hao-Fei Fan leg; ZSCTC-LI-0031 to 0032.
Diagnosis.
The absence of a specialized abdominal tergal gland places the new species into the simoni-species group. This species is similar to N.adebratti Roth, 1994, N.babindaensis Roth, 1994, N.clavate Andersen & Kjaerandsen, 1995, N.gerlachi Roth, 2003, N.pheromosa Lucañas & Maosheng, 2023, N.scytale Andersen & Kjaerandsen, 1995, N.wliensis Andersen & Kjaerandsen, 1995, and N.xiai Liu, Zhu, Dai & Wang, 2017, with tegmina and wings developed. It differs from N.adebratti and N.gerlachi by its ocelli absent (Fig. 5F), while in N.adebratti and N.gerlachi ocelli are present. The subgenital plate of the newly described species is symmetrical (Fig. 5L), while in N.adebratti, N.babindaensis, N.clavate, N.gerlachi, N.pheromosa, N.scytale, and N.wliensis, the subgenital plate is asymmetrical. It differs from N.xiai as follows: 1) tegmina and wings extending beyond the end of abdomen, body length is about half of the wing length, while in N.xiai tegmina and wings slightly extend beyond the end of abdomen, but not exceeding half of the body length (Fig. 1A); 2) eyes well developed, while in N.xiai eyes reduced (Fig. 1B); and 3) accessory hook-like phallomere (L4N) fin-shaped, whereas L4N is parabola-like in N.xiai (Fig. 1C).
Nocticolacordiformis sp. nov.: A male, dorsal view B male, ventral view C female, dorsal view D female, ventral view E pronotum F head G front femur H tegmen I wings J supra-anal plate, ventral view K tarsal claw L subgenital plate, ventral view N hind tarsus; male genitalia, M phallomeres. Abbreviations: L1, L2, L3, L4G, L4N, L4M sclerites of the left phallomere (L1 situated in the central dorsal wall; L2 arch-shaped sclerite situated in the ventral to L1; L3 situated in the left wall protrudes a large hook-process; L4G situated in the posterior ventral wall of the ventral lobe; L4N accessory hook-like phallomere; L4M situated in the ventral wall); R1, R2, R3, R1H sclerites of the left phallomere (R1 situated in the dorsal or ventral walls, or in the posterior part of dorsal and ventral wall; R2 a ridge on the ventral margin; R3 plate-like situated in the anteriormost ventral wall; R1H a larger lobed situated in the dorsal wall, with extensions into the ventral wall).
Measurements (mm).
Male, pronotum: length × width: 0.81–1.02 × 0.99–1.21, tegmen: 4.04–4.25, overall length (including tegmen): 4.83–5.24, body length (the length from the tip of vertex up to the tip of abdomen): 3.19–3.27. Female, pronotum: length × width: 1.13–1.22 × 1.35–1.42, body length (the length from the tip of vertex up to the tip of abdomen): 3.45–3.66.
Description.
Small size. Body tawny. Male. (Figs 5A, B, 6C). Head: vertex of head exposed; Eyes well developed, with heptagon concave; ocelli absent (Fig. 5F). Pronotum subtrapezoidal, densely pubescent, anterior margin and lateral margin with 12 setae (Fig. 5E). Tegmina and hind wings: tegmina and wings well developed, extending beyond the end of abdomen, body length is about half of the wing length (Fig. 5A, B), veins reduced in number, densely pubescent (Fig. 5H, I). Legs: legs long and slender. Anteroventral margin of front femur Type C1 (Fig. 5G); the first tarsus of the hind leg longer than the sum of the remaining tarsi; tarsal claws symmetrical and unspecialized (Fig. 5K), arolium and pulvillus absent (Fig. 5N). Abdomen and genitalia: abdominal tergal gland unspecialized. Supra-anal plate symmetrical, middle of the hind margin concave (Fig. 5J). Subgenital plate symmetrical, middle of the hind margin weakly concave (Fig. 5L). Style absent. Male genitalia: L3 elongate, ventral to hook with 7 strong setae; L4N fin-shaped; L2 narrow; left and right process of L1 towards curved, heart-shape; R1 long handle-like, covered with some setae; R2 sinuate protrusion, with scale-like tubercles; R3 membranous; R1H reduced, rounded margin, with scale-like tubercles (Fig. 5M).
Habitats of Nocticolacordiformis sp. nov. from China A, B habitats of holotype specimen of N.cordiformis sp. nov. C adult of N.cordiformis sp. nov. on wood.
Female: Apterous. Supra-anal plate triangular, transverse of hind margin, middle with triangular invagination. Subgenital lobate (Fig. 5C, D). Cerci 11 segments; ventral surface of segments not spinous setae.
Etymology.
The specific name is derived from the Latin word cordiformis, in reference to the L1 heart-shaped structure of left aedeagus.
Distribution.
China (Guangxi).
Nocticola
appendiculata
sp. nov.
Taxon classificationAnimaliaBlattodeaNocticolidae
FEB017E3-866B-546B-AD90-DB3D801BE14C
https://zoobank.org/C49BF683-1E94-4EFF-8E5A-E49684049B8C
Type material.
Holotype: China • 1 ♂; Guangxi Province, Guilin City, Lingchuan County, Haiyang Town, Xiaofu Village, 25°15.17'N, 110°35.33'E; 329 m; 26 April 2024; Hao-Fei Fan leg; ZSCTC-LI-0033. Paratype: China • 1 ♂, 1 ♀; same data as for holotype; Hao-Fei Fan leg; ZSCTC-LI-0034 to 0035 • 2 ♂; Guangxi Province, Guilin City, Lingui District, Huixian Town, Edi Village, 25°3.67'N, 110°10.25'E; 176 m; 29 April 2024; Hao-Fei Fan leg; ZSCTC-LI-0036 to 0037.
Diagnosis.
The fourth abdominal tergum specialized tergal gland places the new species into the uenoi-species group. It closely resembles Nocticolacurrani Trotter, McRae, Main & Finston, 2017 in terms of the well-developed tegmina and fourth abdominal tergum specialized. It differs from N.currani as follows: 1) wings curved, longer than tegmina and extending beyond the end of abdomen, while in N.currani wings reduced, not extending beyond the first abdomen; 2) ventral of L3 hook with ~ 10 strong setae scattered, while in N.currani with 11 long and strong setae clustered on distal end below the curve of L3 hook; and 3) accessory hook-like phallomere (L4N) inner margin curved and smooth at distal end, whereas L4N is spear-shaped, with longitudinal ribbing of distal end in N.currani. In addition, the tegmina of this species are distinctly longer than the end of the abdomen, with a small appendicular field, while the tegmina of N.australiensis Roth, 1988, N.uenoikikaiensis Asahina, 1974, N.uenoimiyakoensis Asahina, 1974, N.uenoiuenoi Asahina, 1974 and N.rohini (Fernando, 1962) are shorter than the end of the abdomen, and without appendicular field.
Measurements (mm).
Male, pronotum: length × width: 1.01–1.21 × 1.08–1.29, tegmen: 2.66–2.87, wings: 3.05–3.26, overall length (including tegmen): 4.08, body length (the length from the tip of vertex up to the tip of abdomen): 2.48–2.65. Female, pronotum: length × width: 1.10 × 1.33; body length (the length from the tip of vertex up to the tip of abdomen): 3.50.
Description.
Small size. Nymphs whitish (Fig. 8B). Body yellowish. Male (Figs 7A, B, 8B, C, D). Head: vertex of head exposed; eyes reduced; ocelli absent (Fig. 7D). Pronotum ovoid, densely pubescent, hind margin weakly concave, anterior and lateral margins with 12 setae (Fig. 7B). Tegmina and hind wings: tegmina extending beyond the end of abdomen, with a small appendicular at the apical (Fig. 7H). Wings inflexion, longer than tegmina and extending beyond the end of abdomen, veins indistinct, with some setae. Hind wings are placed on both sides of the abdomen and not covered by the tegmina (Fig. 7A, B, I). Legs: legs long and slender. Anteroventral margin of front femur Type C1 (Fig. 7G); the first tarsus of the hind leg longer than the sum of the remaining tarsi; tarsal claws symmetrical and unspecialized (Fig. 7L), arolium and pulvillus absent (Fig. 7O). Abdomen and genitalia: second and third abdominal tergum deeply concave on hind margin, exposing the gland of the fourth abdominal tergum. Fourth abdominal tergum specialized, median area has dense setae convex and a large and deep invagination; Posterior margin has three convexities, with the middle one being smaller than the two sides (Fig. 7J, K). Supra-anal plate symmetrical, middle of the hind margin concave (Fig. 7N). Subgenital plate symmetrical, middle of the hind margin weakly concave (Fig. 7M). Style absent. Male genitalia of left phallomere: L3 elongate, ventral to hook with ~ 10 strong setae scattered; L4N inner margin smooth; L2 narrow; L1 long, apex of left process slightly curved towards L4N. Male genitalia of right phallomere: R1 medial border with several long setae; R2 sinuate protrusion, with scale-like tubercles; R3 membranous; R1H reduced, rounded margin, with scale-like tubercles (Fig. 7P).
Nocticolaappendiculata sp. nov. A male, dorsal view B male, ventral view C female, dorsal view D female, ventral view E pronotum F head G front femur H tegmen I wings J, K male T4, dorsal view L tarsal claw N supra-anal plate, ventral view M subgenital plate, dorsal view O hind tarsus, male genitalia P phallomeres. Abbreviations: L1, L2, L3, L4G, L4N, L4M: sclerites of the left phallomere (L1 situated in the central dorsal wall; L2 arch-shaped sclerite situated in the ventral to L1; L3 situated in the left wall protrudes a large hook-process; L4G situated in the posterior ventral wall of the ventral lobe; L4N accessory hook-like phallomere; L4M situated in the ventral wall); R1, R2, R3, R1H: sclerites of the left phallomere (R1 situated in the dorsal or ventral walls, or in the posterior part of dorsal and ventral wall; R2 a ridge on the ventral margin; R3 plate-like situated in the anteriormost ventral wall; R1H a larger lobed situated in the dorsal wall, with extensions into the ventral wall).
Habitats of Nocticolaappendiculata sp. nov. from China A habitats of N.appendiculata sp. nov. B nymphs and an adult of N.appendiculata sp. nov. on wood CN.appendiculata sp. nov., side view D female and male of N.appendiculata sp. nov. mating.
Female: Apterous (Fig. 8D). Supra-anal plate triangular, transverse of hind margin, U-shaped invagination slightly to the right in the middle of hind margin. Subgenital lobate (Fig. 7C, D). Cerci with 11 segments; ventral surface of segments without spinous setae.
Etymology.
The specific name is derived from the Latin word appendiculatus, in reference to the extended appendicular field of tegmina.
Distribution.
China (Guangxi).
Molecular analysis based on COI
We collected the 14 COI sequences of the three new species and blasted them on GenBank. The resulting sequence alignment comprised 658 nucleotides. Sequence comparisons reveal sequence differences in nucleotide numbers (Table 2, above diagonal), as well as intra- and interspecific genetic distances (Table 2, below diagonal). The intraspecific genetic distance of these three new species is 0. The interspecific genetic distance between N.baiguensis sp. nov. and N.cordiformis sp. nov. is 19.40%, while that between N.baiguensis sp. nov. and N.appendiculata sp. nov. is 21.05%; The genetic distance between N.appendiculata sp. nov. and N.cordiformis sp. nov. is 4.26%. The COI sequence of these two species differ in 27 nucleotide sites.
The phylogenetic tree of the 20 COI sequences, derived from Table 1, is depicted in Fig. 9. The maximum likelihood (ML) tree shows that samples with the same morphology exhibit high bootstrap values in forming monophyletic groups, although most of the other nodes did not have high bootstrap values.
Maximum likelihood (ML) tree of N.baiguensis sp. nov., N.cordiformis sp. nov. and N.appendiculata sp. nov. based on a portion of the COI sequence. Bootstrap values are reported above each branch.
Discussion
In this study, we used morphological differential diagnosis to conclude that N.baiguensis, N.cordiformis, and N.appendiculata are new species of the genus Nocticola. The three species have significant differences in male genitalia. Molecular comparison of these three species show that the maximum genetic distance between them was 21.05% (N.baiguensis and N.cordiformis), while the minimum interspecies genetic distance was 4.26% (N.cordiformis, and N.appendiculata).
Nocticola is mainly distributed in Africa, Asia, and Australasia (Beccaloni 2014), with two species N.xiai Liu, Zhu, Dai & Wang, 2017 and N.sinensis Silvestri, 1946 recorded in China, both of which are termitophilous (Silvestri 1946; Liu et al. 2017); the other two termitophilous species are N.termitophila Silvestri, 1946 and N.jodarlingtonae Roth, 2003 (Silvestri 1946; Roth 2003a). The four species of Nocticola were found in termite nests, but at that time, there were no research records of a relationship between Nocticola and termites. In March 2023, we discovered a female Nocticola sp. with reduced eyes in Jiangjunshan Park, Zhuhai City, Guangdong Province. It was found in a relatively dry, mud-built, ant nest at the bottom of some decaying wood; the ant species was identified as Camponotusnicobarensis Mayr, 1865. Unfortunately, we only found one female Nocticola sp. Due to the scarcity of evidence, we cannot determine whether this Nocticola sp. accidentally entered the nest or had a relationship with C.nicobarensis.
This article describes three new species: N.baiguensis sp. nov., N.cordiformis sp. nov., and N.appendiculata sp. nov. Among them, N.baiguensis sp. nov. was first discovered in the Baigu Cave, which is a natural karst cave. We found them on some decaying branches and stones next to the bat feces. We collected four pairs of N.baiguensis sp. nov. in the cave, and one of them was surrounded by many small nymphs. We brought them back to the laboratory for breeding. Kept under observation, we found that they feed on mycelium on wood at a slow rate. They prefer to hide under the bottom of wood and wet tissues and did not seem to be very active. We also found some ants identified as C.nicobarensis living together with N.baiguensis sp. nov.
China is a country with the largest area of karst caves, a total of approximately 3.4 million square kilometers and approximately 500,000 karst caves (Zhang et al. 2021). Most of the karst caves have constant temperature and humidity inside because of the underground space with dark environments and complex terrain (Ran 2013). Due to some social and historical reasons, the exploration of cave-dwelling organisms started relatively late in China (Zhang 1992), resulting in few reports on cave insects and the present research on cave cockroaches. Nocticolacordiformis sp. nov. and N.appendiculata sp. nov. were found under rocks at the roadside. The eyes and wings of N.cordiformis sp. nov. are well developed, while the eyes of N.baiguensis sp. nov. and N.appendiculata sp. nov. were smaller. Nocticolaappendiculata sp. nov. has well-developed front and hind wings, whereas tegmina are well developed and hind wings reduced in N.baiguensis sp. nov. During the sample collection process, it was common to find N.cordiformis sp. nov. and N.appendiculata sp. nov. living under the same stone, with overlapping areas, although they have certain differences in morphology and COI sequences. Further research on the living habits of Nocticola and exploration of their relationship with ants should be carried out in the future.
Supplementary Material
XML Treatment for Nocticola
XML Treatment for Nocticola sinensis
XML Treatment for Nocticola baiguensis
XML Treatment for Nocticola cordiformis
XML Treatment for Nocticola appendiculata
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Andersen T Kjaerandsen J (1995) Three new species of Nocticola Bolívar from Ghana, West Africa (Blattaria: Nocticolidae).Journal of African Zoology 109(4): 377–385.
- 2Beccaloni GW (2014) Cockroach Species File Online. Version 5.0/5.0.World Wide Web Electronic Publication. http://Cockroach.Species File.org [Accessed 1 April 2024]
- 3Bolívar I (1892) Voyage de M. E. Simon aux l’île Philippines (Mars et Avril 1890). Études sur les Arthropodes cavernicoles de l’île de Luzon.Annales de la Société Entomologique de France 61: 29–34. 10.1080/21686351.1892.12279282 · doi ↗
- 4Bruner L (1915) Preliminary catalogue of the Orthopteroid insects of the Philippine Islands.University Studies of the University of Nebraska 15: 195–281.
- 5Chopard L (1932) Un cas de microphtalmie liée à l’atrophie des ailes chez une blatte cavernicole. In: Société Entomologique de France (Ed.) Livre du Centenaire.Société Entomologique de France, Paris, 485–496.
- 6Chopard L (1946) Note sur quelques Orthoptères cavernicoles de Madagascar.Revue Francaise d’Entomologie 13: 146–155.
- 7Chopard L (1950) Les blattes cavernicoles du genre Nocticola Bol.Eos, Revista Española de Entomología 29: 301–310. http://hdl.handle.net/10261/153622
- 8Chopard L (1966) Une espèce nouvelle de Nocticola provenant d’une grotte du Transvaa (Dictyoptères, Nocticolidae).In: Bulletin de la Société entomologique de France 71: 307–310. 10.3406/bsef.1966.20908 · doi ↗