Evidence for Auditory Stimulus‐Specific Adaptation But Not Deviance Detection in Larval Zebrafish Brains
Maya Wilde, Rebecca E. Poulsen, Wei Qin, Joshua Arnold, Itia A. Favre‐Bulle, Jason B. Mattingley, Ethan K. Scott, Sarah J. Stednitz

TL;DR
This study shows that larval zebrafish brains adapt to repeated sounds but do not detect unexpected ones, similar to how humans and other animals do.
Contribution
The first evidence of stimulus-specific adaptation in larval zebrafish auditory systems is presented.
Findings
Zebrafish brains show frequency-specific adaptation to repeated sounds.
No deviance detection was observed for unexpected sounds or omissions in sound sequences.
Whole-brain calcium imaging revealed variable response amplitudes to different sound frequencies.
Abstract
Animals receive a constant stream of sensory input, and detecting changes in this sensory landscape is critical to their survival. One signature of change detection in humans is the auditory mismatch negativity (MMN), a neural response to unexpected stimuli that deviate from a predictable sequence. This process requires the auditory system to adapt to specific repeated stimuli while remaining sensitive to novel input (stimulus‐specific adaptation [SSA]). MMN was originally described in humans, and equivalent responses have been found in other mammals and birds, but it is not known to what extent this deviance detection circuitry is evolutionarily conserved. Here we present the first evidence for SSA in the brain of a teleost fish, using whole‐brain calcium imaging of larval zebrafish at single‐neuron resolution with selective plane illumination microscopy. We found frequency‐specific…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
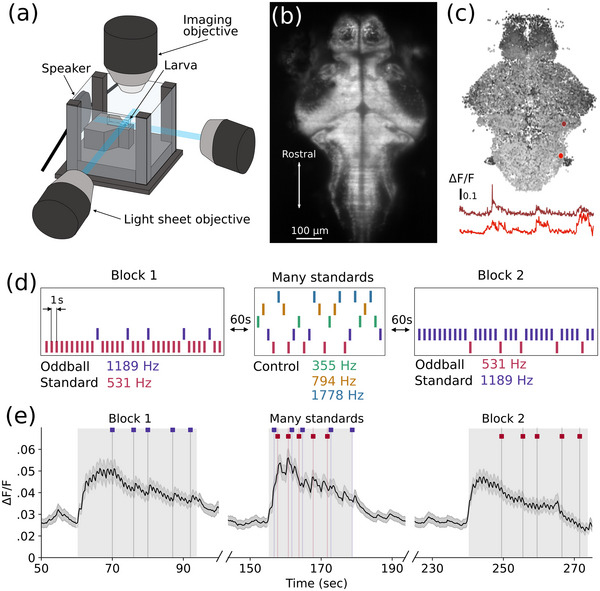 FIGURE 1
FIGURE 1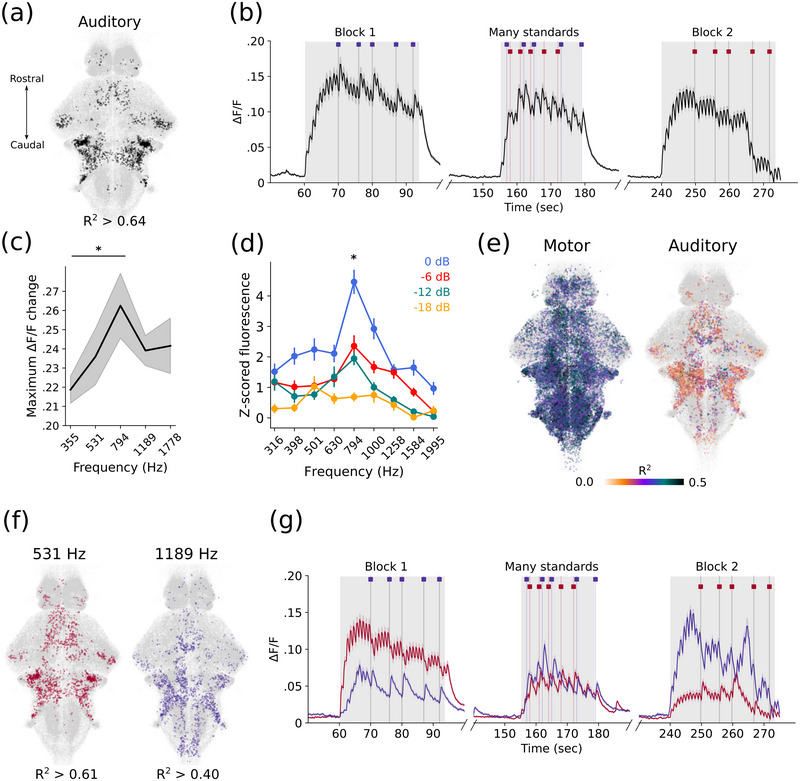 FIGURE 2
FIGURE 2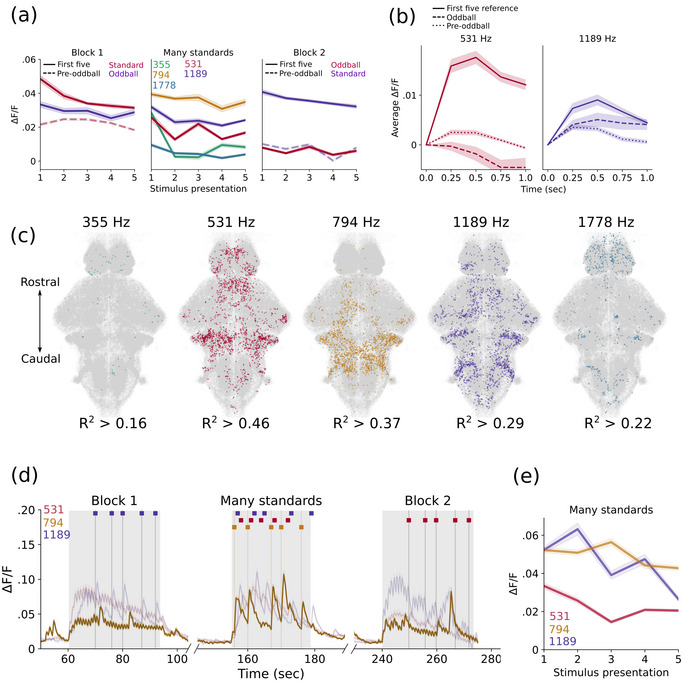 FIGURE 3
FIGURE 3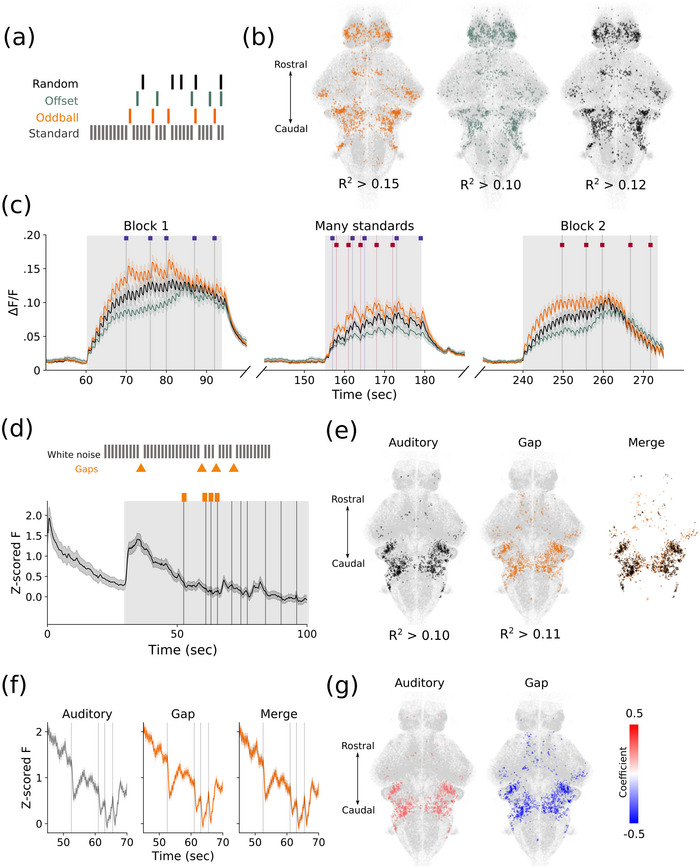 FIGURE 4
FIGURE 4- —National Health and Medical Research Council 10.13039/501100000925
- —Australian Research Council 10.13039/501100000923
- —Simons Foundation 10.13039/100000893
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeuroscience and Music Perception · Neural dynamics and brain function · Animal Vocal Communication and Behavior
Introduction
1
Animals receive a constant stream of sensory input, and detecting changes in this sensory landscape is critical to their survival. Our understanding of the neural circuits underlying this process across species is limited, especially at the level of individual neurons. In humans, one measure of brain activity following an unexpected change in sensory stimuli is known as mismatch negativity (MMN) (Näätänen et al. 2007; Schröger 1998; Fitzgerald and Todd 2020). The mismatch refers to the elevated response to a deviant, unexpected stimulus (such as a different frequency tone) compared to a standard, predictable stimulus (Näätänen et al. 2007). In the case of auditory stimuli, this signal is detected in the auditory cortex using electroencephalography or magnetoencephalography. Therefore, MMN in humans represents average activity in the auditory cortex, and the effect has not been recorded at the single‐neuron level. A key feature of single‐neuron activity underlying the MMN is stimulus‐specific adaptation (SSA), where a neuron will decrease its response to a specific repeated stimulus, but this decrement does not generalize to all sounds (Carbajal and Malmierca 2018). Although recognized as a crucial element of identifying deviant stimuli, it is debated whether SSA alone can represent true deviance detection or whether a “pure” mismatch effect requires SSA to be ruled out as a contributor (Carbajal and Malmierca 2018; Willmore and King 2023; May 2021). There is evidence for SSA in subcortical auditory structures in both humans (Cacciaglia et al. 2015) and rodents (Parras et al. 2017; Malmierca et al. 2019), notably the inferior colliculus and the medial geniculate nucleus of the thalamus (Carbajal and Malmierca 2018; Duque et al. 2015; Grimm and Escera 2012). Subcortical SSA is also considered crucial for the MMN but is typically weaker than cortical SSA (Willmore and King 2023).
MMN is reduced in some neurological conditions such as schizophrenia and autism, and it therefore holds promise as a biomarker to aid understanding of neural circuitry underlying differences in sensory processing (Schall 2016; Todd et al. 2013). Developing MMN paradigms in animal models enables detailed investigation into the phenomenon and perturbations that affect it, leveraging their smaller brains to use techniques that are not feasible in human studies. An equivalent of the auditory MMN response is well established in rodents (Todd et al. 2013; Harms et al. 2016; Harms et al. 2014). MMN‐like responses are also present in the equivalent of the auditory cortex in macaques (Lakatos et al. 2020), cats (Pincze et al. 2002), pigeons (Schall et al. 2015), and songbirds (Mori and Okanoya 2022), and there is preliminary evidence for auditory deviance responses in bats (Wetekam et al. 2024), dogs (Howell et al. 2012), owls (Bala and Takahashi 2000), and frogs (Yue et al. 2017), though interpretation of these findings is confounded by a lack of role reversal for the expected and unexpected sounds. MMN responses can be detected in human infants as young as 6 months (Trainor et al. 2003; Themas et al. 2023), but to the best of our knowledge, MMN in early developmental stages is untested in other animals. MMN can also be elicited in other sensory modalities (Näätänen et al. 2007), and indeed visual MMN has been characterized in mice using calcium imaging (Tang et al. 2023), and there is evidence for entrained responses to rhythmic visual stimuli in fishes (Bullock et al. 1990; Karamürsel and Bullock 1994; Sumbre et al. 2008).
Although the full complexity of the human MMN response may not be reflected in other species (Harms et al. 2016), it stands to reason that the ability to detect changes in the sensory landscape would have arisen early in evolutionary time. It is not known to what degree this auditory deviance response is conserved across species, and it is not yet described in teleost fishes. Filling this gap in the evolutionary tree and identifying which homologous brain structures are required could help clarify whether these circuits evolved early in a common ancestor or after the divergence of fish from mammals and birds.
Larval zebrafish are an appealing model for identifying whole‐brain circuits underlying auditory deviance responses (Friedrich et al. 2010). They are readily genetically modified, largely transparent, and have small brains, enabling the use of genetically encoded calcium indicators for whole‐brain imaging of activity at single‐neuron resolution (Vanwalleghem et al. 2018; Ahrens et al. 2012). Additionally, they share many evolutionarily conserved and functionally homologous brain regions with other vertebrates, including mammals (Wullimann and Mueller 2004).
At 6‐day post‐fertilization (dpf), zebrafish larvae show frequency‐specific neuronal responses to sounds across a hearing range of at least 2 kHz (Poulsen et al. 2021). Although there is not currently evidence for predictive responses in the auditory domain within the zebrafish visual system, there is evidence for the accumulation of sensory information and putative prediction error responses (Sumbre et al. 2008; Bahl and Engert 2020; Narayanan et al. 2024; Cheng et al. 2014). These studies use visual stimuli presented at a slower time course than is typically used for auditory MMN: many seconds, compared to auditory MMN stimuli, which are typically tens of milliseconds long, with hundreds of milliseconds interstimulus interval (ISI). Nevertheless, these studies provide evidence that larval zebrafish exhibit a degree of sensory predictive activity. This predictive activity, coupled with their ability to distinguish different sounds, is necessary precursor to auditory deviance responses. We therefore designed an auditory stimulus paradigm to elicit deviance responses in larval zebrafish and simultaneously imaged whole‐brain neuronal activity at single‐cell resolution.
Methods
2
Animals
2.1
We generated larvae from a stock of adult transgenic zebrafish (Danio rerio, Tg(elavl3:H2B‐GCaMP6f), ZFIN identifier: ZDB‐ALT‐150916‐4, RRID: Addgene_67159) with targeted expression of the fluorescent calcium indicator GCaMP6f in the nuclei of neurons (Chen et al. 2013), maintained on a TLN background at a density of 10–15 fish per liter. Fish were obtained from the Zebrafish International Resource Center and housed locally in a dedicated zebrafish facility at the University of Queensland. Adults were crossed in group mating tanks to produce mitfa ^−/−^ larvae that lack melanophores to facilitate imaging. Experimental animals were obtained from at least four group matings. Larvae were then maintained in an incubator (Thermoline refrigerated incubator 360L TRIL‐360‐DL) kept at 28.5°C, with a 14/10 h light/dark cycle (Westerfield 2000). Larvae were raised in 100 mm diameter petri dishes containing E3 media (distilled water with 10% Hanks solution, consisting of 137 mM NaCl, 5.4 mM KCl, 0.25 mM Na_2_HPO_4_, 0.44 mM KH_2_PO_4_, 1.3 mM CaCl_2_, 1.0 mM 654 MgSO_4_, and 4.2 mM NaHCO_3_ at pH 7.2) (Westerfield 2000), at a density of 50 fish per dish. The background noise in the incubator was approximately 61 dB SPL, as measured with a digital sound level meter (Benetech GS1357). Larvae were not fed prior to imaging to reduce developmental variability. We used 6 dpf larvae because we previously characterized the auditory system and experimental protocol for this age, and it is possible to capture the entire volume of the larval brain with the 20× objective used in our microscope. Further, a number of anatomical atlases are available at this stage, allowing us to compare our data to known structures. Zebrafish housing, breeding, larval maintenance, and experiments were performed with approval from the University of Queensland Animal Ethics Committee (IMB/237/16/BREED and SBS/341/19).
Calcium Imaging
2.2
Larval zebrafish aged 6 dpf were mounted upright on a raised opaque plastic platform in a custom 3‐dimensional (3D)‐printed imaging chamber (24 × 24 mm^2^) (Poulsen et al. 2021; Constantin et al. 2020), fully embedded in 2% low melting point agarose. The sides of the agarose were cut away to provide a perpendicular surface to the light sheet across all depths and reduce variability in agarose volume. Chambers consist of corner posts with 20 mm square coverslips affixed using waterproof glue (Liquid Fusion Clear Urethane Adhesive) to form the chamber walls. A speaker was affixed with adhesive to the glass coverslip wall of the chamber to create a water‐coupled speaker (Poulsen et al. 2021). The chamber was then filled with E3 media. Fish were left to settle in the chamber for at least 60 min to prevent z‐drift during acquisition.
Whole‐brain fluorescence calcium imaging was conducted with a custom‐built selective plane illumination microscope (SPIM), as described previously (Taylor et al. 2018; Panier et al. 2013; Favre‐Bulle et al. 2018; Marquez‐Legorreta et al. 2022). We used two light sheets, from the anterior and one side of the fish, to illuminate the full brain (Figure 1a). Sheets were scanned at 25 10 µm steps throughout the dorsoventral axis of the animal at a rate of 4 volumes per second. Both the image acquisition and delivery of acoustic stimuli were controlled using Micro Manager software (version 1.4) (Edelstein et al. 2010). An example image of a single plane is shown in Figure 1b.
Calcium imaging of responses to auditory stimuli. (a) Experimental setup. (b) Example single plane of a zebrafish larval brain expressing nuclear localized GCaMP6f under the control of the HuC promoter. (c) Visualization of all neurons segmented from a single larva colored by depth. (d) Stimulus train. (e) Average ΔF/F traces over all neurons (n = 186,675) from all fish (n = 12) for each block of the experiment. Note: (a) A larva mounted in low melting point agarose is illuminated by two perpendicular sheets of light. Acoustic stimuli are delivered by a speaker affixed to the back wall. Calcium activity is captured with a water‐immersion objective. (b) Image was averaged over the entire recording after motion correction. (c) Black is most ventral, white most dorsal. Example neurons are highlighted in red, with their respective calcium activity traces over time represented below. (d) Experimental tones of 531 Hz (pink) and 1189 Hz (purple) each act as the standard and deviant stimulus in different blocks. Control tones of 355 Hz, 794 Hz, and 1778 Hz enable each frequency to be presented at the same probability in the many standards control blocks. (e) Grey boxes indicate the timings of the stimulus blocks. Stimulus times for the experimental tones when not standard are illustrated above each graph.
Image Processing Pipeline
2.3
Individual regions of interest (correlating to neuronal nuclei) were segmented using Suite2p (Pachitariu et al. 2016), and their fluorescence traces over the course of the experiment were extracted (Figure 1c). The motion‐corrected 3D mean image of each fish was then registered to the ZBrain reference larval zebrafish brain atlas (Randlett et al. 2015), using Advanced Normalization Tools (ANTs) (Avants et al. 2008). These steps were completed within a custom data processing pipeline run on the high‐performance computing cluster at the University of Melbourne. These data were then extracted and sorted in MATLAB (MathWorks, 2022b). Masks from the ZBrain atlas were used to remove any regions of interest detected in the eyes or otherwise outside the brain.
Acoustic Stimuli
2.4
Sounds were delivered to the fish with a mini speaker (Dayton Audio DAEX‐9‐4SM Skinny Mini Exciter Audio, Haptic Item Number 295‐256), driven by an amplifier (Dayton Audio DA30 2 × 15 W Class D Bridgeable Mini Amplifier). The characteristics of the acoustic field within this setup have been described previously using accelerometer and hydrophone recordings (Poulsen et al. 2021). We used three paradigms to assess the presence of auditory deviance detection: an “oddball” paradigm where an unexpected frequency is played during an otherwise repetitive tone sequence, a loudness curve to control for any salience differences between frequencies used in the oddball paradigm, and an “oddball” paradigm where the unexpected stimulus is a gap in white noise to reveal deviance‐specific responses in the absence of auditory related activity.
Oddball Tones
2.4.1
The five frequencies were chosen to be equally spaced on a logarithmic scale within the hearing range of larval zebrafish (Poulsen et al. 2021). Sound stimuli were 100 ms in length, with 2 ms on‐ and off‐ramps in volume. The volume was set to −6 dBFS (decibels digital full scale), equivalent to 96 dBSPL (decibels sound pressure level) when measured in an empty chamber with a digital sound level meter (Benetech GS1357) (Constantin et al. 2020). The stimulus paradigm for these experiments followed a “flip, many standards, flop” paradigm (Figure 1d) (Harms et al. 2014). Each block consisted of 10 repetitions of the standard stimulus, followed by a pseudorandom sequence of oddball tones representing 20% of the total stimuli for that block, for a total of 25 stimulus presentations per block. The many standards block consisted of 25 stimuli, with 5 frequencies occurring with 20% likelihood. Unless otherwise noted, the ISI was 1 s. The inter‐block interval was 60 s. All larvae (n = 12) received the same pseudorandom stimulus train, with the same order of stimulus blocks, for a total of 95 stimuli.
Loudness Curves
2.4.2
Nine frequencies were chosen to be equally spaced on a logarithmic scale within the hearing range of larval zebrafish, each played for 100 ms at four volumes: 0, −6, −12, and −18 dBFS. The order of these was randomized into two different sequences, and each fish (n = 18) received one of these.
Silent Gaps
2.4.3
Acoustic stimuli were 100 ms white noise bursts at −6 dBFS, with 2 ms on‐ and off‐ramps, and the deviant event was the absence of this stimulus. These were delivered with an ISI of 500 ms. Thirty white noise bursts were delivered to establish the sequence of sounds, then 400 stimulus timepoints, 40 (10%) of which were gaps (white noise was not delivered). At least two white noise bursts were delivered between each instance of a gap. All fish (n = 9) received the same pseudorandom stimulus sequence, totaling 390 sounds and 40 gaps.
Analysis
2.5
Auditory regressors were generated from stimulus onset times convolved to a synthetic calcium signal that mimics the rise and decay dynamics of our fluorescent probe. Regressors for motor activity in each fish were similarly generated using the motion correction output from Suite2p. Cells were considered significant for a regressor if they met two criteria: (1) a p value < 0.001 divided by the total number of cells in the experiment across animals and (2) an R ^2^ value greater than the 99th percentile of all cells, for that regressor.
Permutation clustering analysis was performed using k‐means clustering of ΔF/F for all cells that passed the threshold for any of our previously described regressors, including fictive controls (8306 cells total). Each iteration consisted of 30 clusters with randomly initialized centers, for a total of 33,000 unique clusters. The mean fluorescence of each cluster was correlated to both our oddball and offset control regressors. Clusters with an R ^2^ value to the oddball regressor greater than the 99th percentile (0.350, for total of 494 cells spanning 60 clusters) were then pooled to improve coverage across animals and used for subsequent calculations.
When applicable, paired t‐tests were used for multiple comparisons with an alpha cutoff of 0.05 adjusted by a Šidák correction for multiple comparisons. Error is reported throughout as standard error of the mean.
Analyses were performed using custom software written in Julia 1.7.2 using GLM.jl and MixedModels.jl (http://github.com/stednitzs/oddball/).
Oddball Tones
2.5.1
The change in fluorescence over the baseline fluorescence (ΔF/F) was calculated using a sliding window of 201 timepoints (50.25 s) and a smoothing window of 7 timepoints.
Stimulus‐Specific Adaptation
2.5.2
We used a linear mixed model to compare average response amplitudes as depicted in Figure 3a and Figure S2b,d, with each fish as a random effect to control for repeated measures. We used fixed effects of frequency, block, and presentation order (first and fifth presentation of the frequency within a given block). These stimulus timings were selected to ensure an equal number of repetitions for both standard and oddball tones. When applicable, paired t‐tests were used for post hoc comparisons with an alpha cutoff of 0.05 adjusted by a Šidák correction for multiple comparisons.
Loudness Curve
2.5.3
Auditory responsive neurons were identified using a regressor to the timing of each sound stimulus. The mean amplitude of the z‐scored responses of these neurons to each frequency at each volume was calculated for each fish to produce the loudness curve. Z‐scored responses were used to directly compare between neurons of different types of brightness to better capture the activity of dim cells that respond weakly to perceptually quiet stimuli. We used a linear mixed model to compare response amplitudes, with each fish as a random effect to control for repeated measures.
Silent Gaps
2.5.4
To better correct for sustained, elevated calcium signals induced by closely spaced white noise stimuli, z‐scored responses were used. Regressions were performed only on time periods during stimulus presentation to account for bleaching at the start of recording. We calculated the mean ΔF/F for gap responsive cells during the gap and performed a linear regression using the number of preceding white noise stimuli as a predictor.
Results
3
Auditory Responses
3.1
The experimental design followed a flip‐many standards‐flop paradigm, consisting of three stimulus blocks (Figure 1d) (Harms et al. 2014). This design enables us to compare responses to the same tone when it is the standard or deviant stimulus and controls for different sensitivity to each frequency. The many standards block is an additional control, where three other frequencies are included such that there is no expectation set by a predictable sequence, but the likelihood of each stimulus is equal to the deviant tone in the predictable sequences.
Using a regression‐based approach, we identified individual cells in animals that responded to specific aspects of the auditory stimuli. First, we classified cells as broadly auditory responsive (responding to all sounds) (Figure 2a,b). This approach permitted us to identify cell populations corresponding to previously identified brain regions in the zebrafish auditory processing pathway, including the octavolateralis nucleus, torus semicircularis, cerebellum, thalamus, and telencephalon (Figure 2a, Figure S1a,b) (Poulsen et al. 2021; Vanwalleghem et al. 2017). These broadly auditory cells respond to reference, many standards, and oddball tones, regardless of the preceding stimulus presentation (Figure 2b). The spatial distribution of these neurons resembles previously identified distributions of pure tone–responsive neurons in the larval zebrafish brain (Poulsen et al. 2021). Overall, auditory‐responsive cells decreased in activity by the last stimulus in a given block when compared to the first, but this recovered after breaks between blocks, indicating the decrease is not due to photobleaching (p < 0.001, linear mixed‐effect model).
*Auditory‐specific responses. (a) Spatial distribution of generally auditory responsive neurons (black). (b) Mean trace of all neurons passing the criteria for classification as generally auditory responsive. (c) Amplitude of the responses of auditory neurons to each frequency in the many standards control block. (d) Equal loudness contours for larval zebrafish, separate dataset. (e) Correlation to motor activity. (f) Spatial distribution of frequency‐specific neurons: Those preferentially responding to 531 Hz (pink) and those preferentially responding to 1189 Hz (purple), from across all animals. (g) Mean traces in each block of the experiment of all 531 Hz‐selective neurons (pink) and 1189 Hz‐selective neurons (purple). Note: (a) R 2 value is the threshold to attain 99th percentile of auditory neurons. (b) Timings of the three experimental blocks are indicated by gray shading behind the trace. (c) Gray shading represents the standard error of mean. *p < 0.01 for 794 versus 355 Hz. (d) Dots indicate means over 18 fish and bars indicate SEM. frequencies are evenly spaced within a logarithmic distribution. Significant effects of volume (p < 0.001) and frequency (p < 0.005), but no significant interaction (p = 0.236). p < 0.01. (e) Left indicates the distribution of motor responsive neurons; right indicates the motor correlation values of neurons that were classified as auditory responsive. Color of dots represents the R 2 value of the correlation to the motor activity. (f) Populations defined by responsiveness to all presentations of that frequency across blocks. R 2 values are the thresholds to attain 99th percentile of each of these populations. (g) Timings of the three experimental blocks are indicated by gray shading behind the trace. Timings of the deviant stimuli are indicated by the colored dashes above the trace.
However, we noticed that the response amplitude in these auditory neurons was lower for 531 Hz tones than 1189 Hz tones in the first block, despite being presented at the same intensity (Figure 2b). This could be a result of either adaptation or salience differences between these frequencies. We computed the maximum change in ΔF/F for each frequency in the many standards block only, finding that on average auditory cells exhibit larger calcium responses to frequencies between 531 and 1189 Hz (Figure 2c). This led us to question if larval zebrafish exhibit equal‐loudness contours similar to other animals, where frequencies in the middle of the hearing range are perceived as louder than those toward the edges, even when presented at equal intensity (Suzuki and Takeshima 2004). We therefore collected another dataset to establish the landscape of equal‐loudness contours, using nine logarithmically spaced frequencies across the hearing range of larval zebrafish (Poulsen et al. 2021). Indeed, the curve produced implies increased loudness for frequencies toward the center of the hearing range compared to the higher and lower ends (Figure 2d). We analyzed this loudness curve statistically and found main effects for both volume and frequency (p < 0.001 and 0.005, respectively) and no overall interaction effect (p = 0.236). At the volume used in the main experiment (−6 dBFS), responses to 794 Hz are significantly higher than frequencies on the periphery (p < 0.01). These curves are evidence that 1189 Hz tones have a greater effective loudness, and therefore salience, than 531 Hz tones across different volumes for larval zebrafish.
Although we recorded animals under a restrained preparation that did not permit the recording of tail movements, we inferred movement attempts from the motion correction derived from our processing pipeline (Figure S1c). Movement efforts are highly correlated to cells in the hindbrain, corresponding to motor command regions (Figure 2e) (Hale et al. 2016). A proportion of auditory‐responsive cells are also correlated with movement events, indicating movements often occur simultaneously to an auditory stimulus, but not consistently enough to suggest the requirement for motion correction arises from vibration of the chamber during stimulus presentation (Figure 2e). We find no consistent overall differences in the behavioral response rate from the two frequencies most relevant to our experimental paradigm (Figure S1d).
Given that some auditory cells may be frequency‐selective, we further defined two populations of neurons that preferred each of the experimental frequencies (Figure 2f,g). These populations are similarly spatially distributed throughout the brain (Figure 2f) and are present in all animals in our experiment (Figure S1e,f). The mean activity traces of each of these populations indicate they also respond to the non‐preferred frequency, but to a lower degree than to their chosen frequency (Figure 2g). In blocks where the non‐preferred stimulus is the standard tone, there is reduced calcium accumulation and lower overall activity, whereas the activity within the many standards control blocks is comparable. This effect is consistent with cells that respond preferentially to specific frequencies, as the number of stimulus presentations in the many standards blocks is equivalent for both tones.
Stimulus‐Specific Adaptation
3.2
We observed that by the second block, responses to the 531 Hz deviant tone were suppressed relative to the first block (Figure 2b,g), which could be explained by SSA. This adaptation is regarded as a key component underlying auditory deviance detection (Carbajal and Malmierca 2018). We measured the mean change in ΔF/F at each stimulus presentation for broadly auditory cells and compared this to the first five presentations of each stimulus in all three blocks (Figure 3a). We find that responses decay over repeated presentations, even when the tone is unexpected. We also included the five standard presentations that directly precede an oddball to account for the total number of preceding stimuli and found that these responses were decreased after many repetitions, consistent with adaptation. In contrast, the responses to the deviant frequency in the first block remain elevated during the many standards, suggesting the five preceding presentations of that frequency were insufficient to induce adaptation. Conversely, in the last block, responses to that frequency as the standard are initially elevated but then decrease to the same level as the adapted frequency. If the activity of these neurons also encoded the unexpected nature of a stimulus, the responses are expected to be greater when it is the oddball rather than the predictable standard preceding an oddball. However, this is only the case in the first block, where the oddball was not previously adapted (Figure 3b). We performed a mixed model analysis comparing the first and fifth responses to each frequency within a block, finding statistical evidence that SSA occurs rapidly in the standard frequency (p = 0.017 and p = 0.005 for the first and second blocks, respectively). In contrast, no such difference is found for the oddball frequency in Block 1 (p = 0.196) or the oddball frequency that served as standard in the first block (p = 0.124). Indeed, the first oddball tone in the second block is statistically indistinguishable from the preceding standard tone, indicating the persistence of SSA over the duration of the experiment (p = 0.127). During the many standards blocks, we found no significant effect for either frequency or repetition number in broadly auditory‐responsive cells, nor an interaction effect between them (p = 0.551, 0.219, and 0.716, respectively).
Stimulus‐specific adaptation. (a) Mean response amplitudes of all auditory‐classified neurons to specific stimulus presentations. (b) Mean traces of frequency‐selective neurons to the first five reference tones of each frequency, the five oddball presentations of each frequency, and the five instances where each frequency directly precedes an oddball when it is the standard. (c) Spatial distributions of five populations of frequency‐specific neurons defined by regressions to their presentation during the many standards control blocks. (d) Mean traces throughout the whole experiment of the 794 Hz‐preferring population (dark yellow). (e) Average responses of each of the three middle frequency‐preferring populations to each of the five presentations of their preferred frequency during the many standards block. Note: (a) In Blocks 1 and 2, solid lines represent the first five presentations of each frequency (as either standard or oddball), and the dotted line represents the five presentations of the standard stimulus directly preceding an oddball. In the many standards block, the mean responses to five presentations of each frequency are shown. Shading represents the standard error of mean. (b) Shading represents standard error of mean. (c) R 2 values represent the threshold to attain the 99th percentile of responses. (d) Fainter lines for the 531 and 1189‐preferring populations are included for reference. Timings of the three experimental blocks are indicated by gray shading behind the trace. Timings of the two experimental frequencies when infrequent and the preferred 794 Hz tone are indicated by the colored dashes above the trace. (e) Shading represents standard error of the mean.
We then used the many standards control block alone to define populations of frequency‐selective cells to assess whether off‐target frequencies produced adaptation in each population and partially control for order effects (Figure 3c). We did not detect populations that correlate well with tones at the extreme ends of the hearing range, consistent with the weak responses detected in our analysis of auditory cells (Figure 2c) and previous studies (Poulsen et al. 2021). We therefore focused on the three middle frequencies and found that each population also responds to the other frequencies, despite preferring its assigned frequency (Figure 3d). However, the populations that prefer the initial standard frequency (531 Hz) are more adapted by the many standard blocks than other populations (Figure 3e). Interestingly, the responses of the 794 Hz neurons are similar in Blocks 1 and 2, suggesting the number of preceding off‐target stimuli doesn't affect future responses to off‐target stimuli; therefore, adaptation is specific to the preferred frequency (Figure 3d). As expected, 1189 Hz neurons maintain high response amplitudes until late in Block 2, despite many preceding off‐target frequency stimuli (Figure 3d). Despite a slight decline in responses of 1189 Hz cells during the many standards block, the difference between the first and fifth stimuli is not significant (p = 0.540).
Additionally, to determine the proportion of cells tuned to specific frequencies and investigate the possibility that a single frequency drives the average response, we tested an absolute threshold based on the R ^2^ value of the lowest percentile‐based cutoff in the many standards block (R ^2^ = 0.228 or greater). We calculated the percentage of auditory responsive cells that overlap with frequency‐tuned cells and found 63.7% of auditory cells (n = 27,989 over 12 fish) do not overlap with frequency‐specific cells.
If these activity patterns are explained by SSA, we would expect the rate and extent of adaptation to decrease with longer ISIs (Willmore and King 2023; Marquez‐Legorreta et al. 2022). To test this, we used an identical protocol with 2 and 3 s ISIs and found longer ISIs produce less pronounced SSA (Figure S2a–d). To quantify this effect, we repeated our mixed model analysis for the standard frequencies, finding only weak evidence for SSA at 2 s ISI when comparing the first and fifth standard presentation in Blocks 1 and 2 (p = 0.066 and 0.014, respectively). However, we observe a significant decrease by the second block (p < 0.001 for both frequencies). In contrast, we found no evidence of SSA within individual blocks in the 3 s ISI dataset regardless of frequency and a significant decrease across both blocks in 531 Hz but not 1189 (p < 0.001 and 0.146, respectively). These findings support the idea that shorter ISIs induce SSA more rapidly.
Oddball Tone
3.3
Given the response amplitude of frequency‐selective neurons was not reliably higher to oddball stimuli than standard stimuli (Figure 3b), we instead looked for a population of neurons that responds specifically to unexpected sounds. Using regressors to the timing of deviant sounds in Blocks 1 and 2, we then identified cells that selectively respond to oddball stimuli in both blocks (Figure 4a–c). As a control, we included two additional variables, consisting of the same number of fictive stimuli locked to the presentation of actual auditory pulses: (1) random, where fictive stimuli can occur at any time during the stimulus train, including during oddball presentations, and (2) offset, where fictive stimuli are constrained to occur only when oddball stimuli are not presented (Figure 4a). These controls allowed us to rule out cells that would be randomly correlated simply by the periodic nature of their firing rate.
No evidence for oddball‐specific responses. (a) Example of regressor timings to the oddball (orange) and the offset (cannot be aligned with oddball, green) and random (can occur at any time, black) controls. (b) Distribution of oddball‐responsive (orange), offset control (green), and random control (black) neurons. (c) Mean traces of neurons classified as high fidelity to the oddball, offset, and random stimulus timings. (d–g) Silent gap experiments. (d) Example of the early part of the stimulus train. (e) Neurons that regress significantly to the white noise bursts versus the gaps and their overlap. (f) Average Z‐scored fluorescence for cells that regress significantly to the white noise burst, the gap, and their overlap. (g) Regression coefficients for auditory and gap‐responsive cells, indicating positive and negative coefficients, respectively. Note: (b) R 2 values are the threshold cutoff for the 99th percentile of responses. (d) The standard stimulus is a white noise burst, and gaps in this sequence are the deviant stimuli (orange). Below is the Z‐scored fluorescence for all cells in the early part of the experiment, average across all fish (n = 9). Vertical lines indicate gaps in the stimulus train. Shaded region indicates the portion of the stimulus train illustrated above. (e) R 2 values are the threshold cutoff for the 99th percentile of responses. (f) The timepoints for these examples match those of the inset in (d).
The cells in each of these three populations had similar spatial distributions (Figure 4b) and were all broadly auditory responsive (Figure 4c). Although we could identify a population of neurons that correlated to the specific timing of the oddball tones, we could similarly find populations with an equivalent correlation to random subsets of stimulus timings. A higher proportion of cells in the telencephalon respond preferentially to oddball tones (33.4% ± 1.4%) than in the offset (4.38% ± 0.9%, p < 0.002) and random conditions (22.2% ± 1.3%, p < 0.001). This effect may be an artifact of high rates of spontaneous activity in the telencephalon and cannot be conclusively identified as a deviance‐selective population using a regression approach.
We considered the strictness of our regression criteria could hinder our ability to detect deviance‐responsive cells. Therefore, we performed a permutation clustering analysis with 100 iterations of k‐means using randomly initialized centers, which should capture even rare patterns of activity. We subsequently correlated the cluster average to our oddball regressor to identify any clusters characterized by deviance responses. Although we were able to identify clusters that were more highly correlated to unexpected stimuli than others (60 clusters with R ^2^ > 99th percentile compared to a mean of 0.158 across all clusters), these consisted of either few cells or were poorly distributed across animals (Figure S2e). We pooled all clusters with R ^2^ > 99th percentile and computed the average response to oddball and reference tones in both blocks, finding no significant differences between them, in contrast to the expected behavior of deviance‐detecting cells (Figure S2f). We further note similar correlations to the fictive offset control and, therefore, conclude that these correlations are spurious.
Silent Gap
3.4
We questioned whether the lack of clear deviance‐specific responses was due to the masking of salience and order effects. We therefore used a paradigm where the deviant stimulus was the absence of a predictable sound so that any response at that time could represent predictive activity (Figure 4d). We identified neuronal populations that responded specifically to the white noise sound stimuli or the gap and found nearly completely overlapping spatial distributions and activity (Figure 4e,f). Indeed, responses to gaps occurred only in cells that were also auditory sensitive and were negatively correlated with gaps (Figure 4g), indicating a drop in calcium consistent with auditory cells. The magnitude of this drop was not correlated to the number of preceding sounds, which would have indicated it was related to how unexpected the gap was (Figure S2g). Finally, we did not find a population of neurons whose activity correlated positively with the gaps, suggesting that zebrafish do not exhibit MMN‐like responses to the unexpected absence of auditory stimuli at this developmental stage.
Discussion
4
Stimulus‐Specific Adaptation in Larval Zebrafish
4.1
We found that the two frequencies of sound used in the experiment had varying salience for the fish (Figure 2c,d), and each recruited frequency‐specific populations of neurons (Figure 2f,g). We also demonstrated here an effective loudness curve for larval zebrafish (Figure 2d), which will aid the design of future research into auditory responses in larval zebrafish. These results confirmed that the larval nervous system has the capacity to distinguish between distinct frequencies across a range similar to that shown by previous studies (Poulsen et al. 2021; Vanwalleghem et al. 2017; Privat et al. 2019). We observed a decrease in the response amplitude across the course of the experiment, representing adaptation (Figure 2b). This decrease is unlikely to be due to photobleaching, as the responses recover after the brief break between blocks of stimuli.
Within the general population of auditory neurons, we found that the degree of adaptation to one frequency did not generalize to the other frequency (Figure 3a,b). Although many stimulus repetitions cause decreased responses to a given sound, the same reduction is not observed when the sound is of a different frequency (Figure 3a).
To understand if this is a property of neurons that respond to any tone or can also be observed in frequency‐selective cells, we measured SSA within populations of neurons that prefer a specific frequency (Figure 3c–e). As expected, we did not find neurons selective for frequencies at the far ends of the hearing range, consistent with the profiles of frequency‐specific populations in a previous study (Poulsen et al. 2021). These neurons were not entirely frequency selective; rather, they exhibit smaller responses to off‐target frequencies (Figure 3d). However, these weak responses to other frequencies do not impact the subsequent response to the preferred frequency. For example, the 1189 Hz cells do not decline in activity until the final block, when it is consecutively repeated. In contrast, the 531 Hz population is already adapted during the first block, when it is the standard tone, and their responses remain low throughout the subsequent blocks.
These results suggest SSA in the auditory domain in larval zebrafish, a critical requirement for the ability to detect auditory deviance responses (Carbajal and Malmierca 2018; Willmore and King 2023; May 2021). The neurons showing SSA are located throughout the fish auditory system: in the octavolateralis nucleus (homologous to the mammalian cochlear nucleus), the torus semicircularis (homologous to the mammalian inferior colliculus), the cerebellum, thalamus, and telencephalon. These auditory regions represent the zebrafish homologs of the subcortical auditory pathway in mammals (Vanwalleghem et al. 2017), which exhibit SSA as a building block underlying auditory deviance detection in higher order brain structures (Carbajal and Malmierca 2018; Willmore and King 2023). Our findings are therefore consistent with the idea that SSA evolved in a common ancestor and that its fundamental mechanisms may be conserved between zebrafish and other species. Although the presence of SSA in larval zebrafish suggests SSA persists into adulthood and would be present in other teleosts, further studies are necessary to confirm these expectations.
SSA, as described here, is a crucial element underlying the ability to detect auditory deviance (Carbajal and Malmierca 2018). Further, a deviance‐specific response can be established by the inclusion of both predictable blocks and the unpredictable many‐standards control, where the response to a given frequency is elevated only when it is unexpected (Harms et al. 2016). Studies in rodents and pigeons successfully identified responses meeting these criteria (Harms et al. 2014, 2016, 2014, 2016; Schall et al. 2015). However, efforts to identify an equivalent of MMN in songbirds and frogs failed to find similarly deviant‐specific responses that cannot be explained by stimulus salience or SSA (Mori and Okanoya 2022; Yue et al. 2017). From our findings, the larval zebrafish fit into the latter category. This does not exclude the possibility that deviance‐specific responses exist in these animals, but perhaps the optimal experimental paradigm has not yet been tested. In our case, larval zebrafish do not have fully developed nervous systems, and true deviance detection may arise later in development.
Future Directions for Identifying Deviance Responses
4.2
A challenge in designing an auditory deviance paradigm for larval zebrafish is the lack of an expected behavioral output. Indeed, although we detected some behavioral responses to auditory stimuli despite larvae being physically restrained, these did not correspond specifically to deviant stimuli. Although it is established that zebrafish possess distinct cell populations that are frequency‐selective (Poulsen et al. 2021), and we recapitulate those findings here, the ethological relevance of this is unclear. For example, some species of teleost fish employ auditory communication, but this is not reported in zebrafish (Schulze et al. 2018; Wysocki and Ladich 2003). Another possibility is that frequency selectivity is a nascent property of the larval brain that will become ethologically relevant as the animal develops further. Identifying the functional purpose of frequency selectivity could inform an experimental paradigm that induces more detectable and consistent deviance responses.
One aspect of deviance detection in mammals is a response to the lack of an expected stimulus. Such an experimental design also eliminates the salience and order effect confounds seen in the experiment with pure tone stimuli, but responses to missing stimuli are more difficult to characterize (Awwad et al. 2023). With this approach, any positive deflections in the activity at the time of the gap between predictable sounds are more likely to be a prediction error rather than a preference for a given frequency. However, the only neurons we found with significant correlation to our regressor to the timings of the gaps were negatively correlated, consistent with the expected behavior of auditory neurons (Figure 4g). The absence of positive responses to gaps suggests the omission of a stimulus in a predictable sequence is not represented in the brain of larval zebrafish. Although we did not find any relationship between the magnitude of these dips and the number of preceding stimuli (Figure S2g), it is possible that these events are encoded by suppression of neurons, and the overall elevated calcium levels in our experiment did not permit us to reliably capture suppression (Vanwalleghem et al. 2021).
On the basis of these results, future studies could be refined to improve the possibility of eliciting deviance‐specific responses in larval zebrafish. The selection of pure tones could be improved by using loudness curves to better match the salience of sounds, as well as introducing more stimuli per block. An initial block of alternating stimuli could also be used to equally adapt both experimental frequencies before the first experimental block. Reversing the order of Blocks 1 and 2 between animals could also be a useful approach, although it may introduce inter‐individual differences in order effects.
Limitations of Our Approach and Future Prospects for Addressing Them
4.3
An inherent limitation of calcium imaging is the speed of the probes (Papaioannou and Medini 2022). Although in this manuscript we utilize the fastest GCaMP6 variant, calcium accumulation in cells limits our ability to detect very small or fast changes, which can be detected with techniques such as electroencephalography. It is possible that smaller ISIs would reveal stronger oddball responses, but the slow rise and decay of our probe did not permit us to investigate this possibility. This limitation is especially evident in the gap experiments, where the large drop in calcium potentially masks subtle differences in activity during this time. Regardless of the speed of the probe, calcium accumulation in cells required us to use highly stringent criteria to exclude possibly confounding results. Voltage indicators or more recent generations of calcium indicators are a promising future avenue to investigate similar paradigms with greater subtlety and faster imaging rates.
The 6 dpf larvae tested in this preparation permit whole‐brain imaging at a single‐cell resolution; however, many higher cognitive functions such as associative learning and social interaction occur at later developmental stages (Yashina et al. 2019; Palumbo et al. 2020; Dreosti et al. 2015; Stednitz and Washbourne 2020; Valente et al. 2012). Although it is difficult to match developmental stages between mammals and fishes, the MMN response in humans is not present in early infancy, is detectable at around 6 months, and becomes more robust with age (Trainor et al. 2003; Themas et al. 2023). It is therefore possible that the SSA of responses detected here in larval zebrafish only become behaviorally relevant later in the life of the animal and that the associated functional circuits are not yet fully established. Similarly to the mammalian cortex, the telencephalon is associated with cognition in adult fish (von Trotha et al. 2014; Stednitz et al. 2018; Portavella et al. 2004), but during the larval period, these same regions are undergoing extensive growth, remodeling, and synaptogenesis (Furlan et al. 2017), and more complex circuitry may not yet be established. True deviance‐detection responses may therefore become present in the telencephalon at later developmental stages. Although our microscope's field of view would not capture the entire brain of zebrafish at older developmental stages, targeted studies of individual brain regions at single‐cell resolution are technically feasible (Jetti et al. 2014; Hollmann et al. 2015; Torigoe et al. 2021).
Conclusions
5
Altogether, we report the first evidence for auditory stimulus‐specific adaptation in teleost fishes. Although these responses are a requirement for deviance detection, we did not find definitive deviance‐specific responses in larval zebrafish, nor did we find responses to the omission of a stimulus in a predictable sequence. Further studies could expand on the experiments presented here to identify the ethological relevance of frequency selectivity in zebrafish and investigate whether true deviance‐detection responses develop with age.
Author Contributions
Experimental design: Maya Wilde, Rebecca E. Poulsen, Jason B. Mattingley, and Ethan K. Scott. Experimental methodology: Maya Wilde, Rebecca E. Poulsen, Itia A. Favre‐Bulle. Data collection: Maya Wilde. Processing of calcium imaging data: Joshua Arnold and Wei Qin. Data analysis: Maya Wilde and Sarah J. Stednitz. Writing: Maya Wilde, Sarah J. Stednitz,, and Ethan K. Scott. Supervision: Sarah J. Stednitz and Ethan K. Scott. All authors read and approved the final manuscript.
Ethics Statement
All work was performed according to research application SBS/341/19 and breeding application IMB/271/19/BREED, which was approved by the Anatomical Biosciences Animal Ethics Committee at the University of Queensland.
Peer Review
The peer review history for this article is available at https://publons.com/publon/10.1002/cne.70046.
Supporting information
Figure S1 Auditory supplemental information.Figure S2 Stimulus‐specific adaptation and oddball supplemental information.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ahrens, M. B. , J. M. Li , M. B. Orger , et al. 2012. “Brain‐Wide Neuronal Dynamics During Motor Adaptation in Zebrafish.” Nature 485: 471–477. 10.1038/nature 11057.22622571 PMC 3618960 · doi ↗ · pubmed ↗
- 2Avants, B. B. , C. L. Epstein , M. Grossman , and J. C. Gee . 2008. “Symmetric Diffeomorphic Image Registration With Cross‐Correlation: Evaluating Automated Labeling of Elderly and Neurodegenerative Brain.” Medical Image Analysis 12: 26–41. 10.1016/j.media.2007.06.004.17659998 PMC 2276735 · doi ↗ · pubmed ↗
- 3Awwad, B. , M. M. Jankowski , A. Polterovich , S. Bashari , and I. Nelken . 2023. “Extensive Representation of Sensory Deviance in the Responses to Auditory Gaps in Unanesthetized Rats.” Current Biology 33: 3024–3030.e 3. 10.1016/j.cub.2023.06.013.37385255 · doi ↗ · pubmed ↗
- 4Bahl, A. , and F. Engert . 2020. “Neural Circuits for Evidence Accumulation and Decision Making in Larval Zebrafish.” Nature Neuroscience 23: 94–102. 10.1038/s 41593-019-0534-9.31792464 PMC 7295007 · doi ↗ · pubmed ↗
- 5Bala, A. D. S. , and T. T. Takahashi . 2000. “Pupillary Dilation Response as an Indicator of Auditory Discrimination in the Barn Owl.” Journal of Comparative Physiology A 186: 425–434. 10.1007/s 003590050442.10879946 · doi ↗ · pubmed ↗
- 6Bullock, T. H. , M. H. Hofmann , F. K. Nahm , J. G. New , and J. C. Prechtl . 1990. “Event‐Related Potentials in the Retina and Optic Tectum of Fish.” Journal of Neurophysiology 64: 903–914. 10.1152/jn.1990.64.3.903.2230933 · doi ↗ · pubmed ↗
- 7Cacciaglia, R. , C. Escera , L. Slabu , et al. 2015. “Involvement of the Human Midbrain and Thalamus in Auditory Deviance Detection.” Neuropsychologia 68: 51–58. 10.1016/j.neuropsychologia.2015.01.001.25556848 · doi ↗ · pubmed ↗
- 8Carbajal, G. V. , and M. S. Malmierca . 2018. “The Neuronal Basis of Predictive Coding Along the Auditory Pathway: From the Subcortical Roots to Cortical Deviance Detection.” Trends in Hearing 22, no. 5: 233121651878482. 10.1177/2331216518784822.PMC 605386830022729 · doi ↗ · pubmed ↗