KMT2C/D mutations in newly diagnosed acute myeloid leukaemia: Clinical features, genetic co‐occurrences and prognostic significance
Wenting Wang, Miao Yang, Xue Zhang, Jiayuan Chen, Shaowei Qiu, Bingcheng Liu, Yingchang Mi, Jianxiang Wang, Hui Wei

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
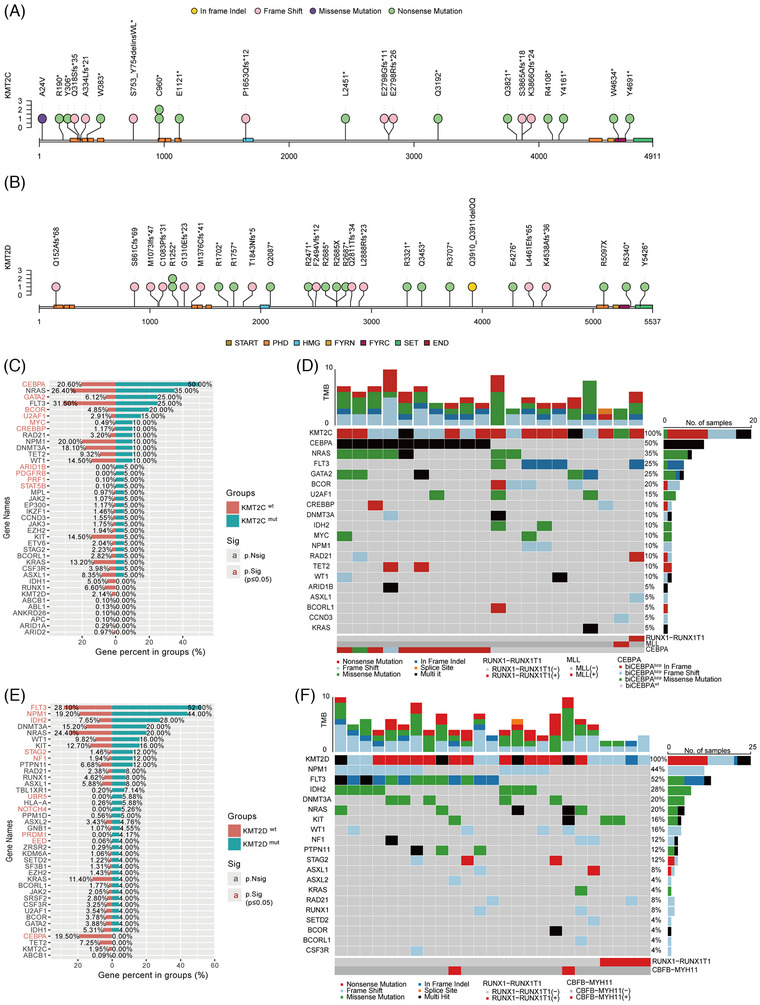 FIGURE 1
FIGURE 1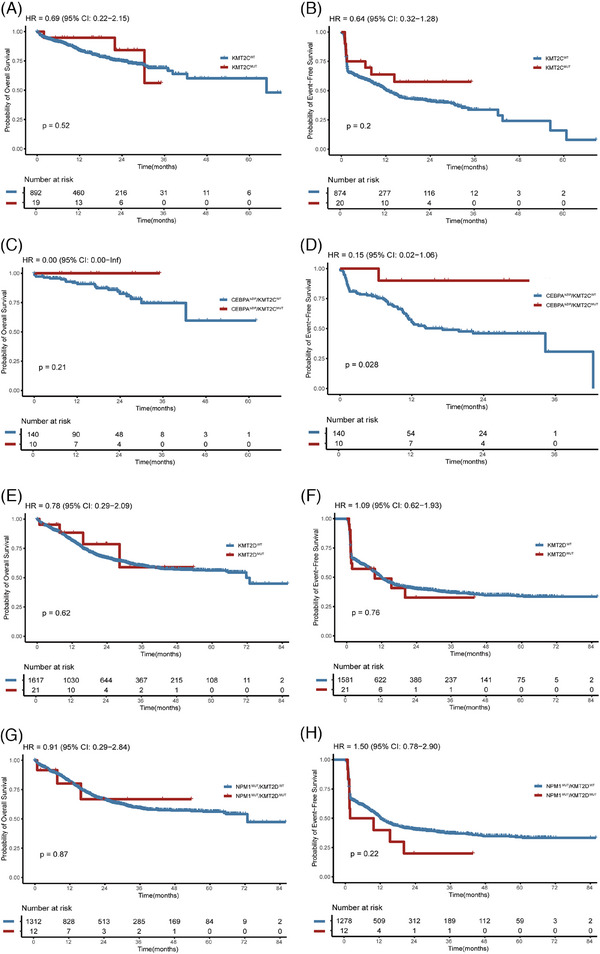 FIGURE 2
FIGURE 2|
( |
( |
( |
| |||
|---|---|---|---|---|---|---|
| Characteristic |
| ( |
| |||
| Age (years) | 43.9 (16–63) | 41.9 (14–79) | .598 | 42.0 (15–71) | 41.9 (13–82) | .976 |
| Gender | .518 | .428 | ||||
| Male ( | 9 (45.0%) | 551 (53.5%) | 11 (44.0%) | 946 (54.0%) | ||
| Female ( | 11 (55.0%) | 479 (46.5%) | 14 (56.0%) | 806 (46.0%) | ||
| Peripheral blood | ||||||
| WBC (109/L) | 23.4 (1.0–106.2) | 30.8 (.6–317.1) | .312 | 21.4 (1.1–132.9) | 33.0 (.5–404.1) | .093 |
| Haemoglobin (g/L) | 87.8 (57.0–135.0) | 84.4 (40.0–144.0) | .552 | 70.0 (47.0–97.0) | 86.1 (34.0–158.0) | <.001 |
| Platelets (109/L) | 58.0 (8.0–244.0) | 54.8 (2.0–376.0) | .811 | 60.0 (16.0–176.0) | 61.8 (2.0–1852.0) | .817 |
| AML classification | .127 | .022 | ||||
| AML with | 1 | 154 | 4 | 284 | ||
| AML with | 0 | 93 | 2 | 132 | ||
| AML with | 0 | 2 | 0 | 8 | ||
| AML with | 0 | 3 | 0 | 8 | ||
| AML with | 1 | 63 | 0 | 104 | ||
| AML with | 2 | 192 | 11 | 317 | ||
| AML with | 10 | 211 | 0 | 340 | ||
| AML with myelodysplasia related | 4 | 148 | 5 | 248 | ||
| AML defined by differentiation | 2 | 164 | 3 | 311 | ||
| Chromosome karyotype | .011 | .410 | ||||
| Normal karyotype | 10 (50.0%) | 460 (44.7%) | 18 (72.0%) | 795 (45.4%) | ||
| T(8;21) | 1 (5.0%) | 154 (15.0%) | 4 (16.0%) | 279 (15.9%) | ||
| Inv(16) | 0 (0%) | 84 (8.2%) | 1 (4.0%) | 122 (7.0%) | ||
| T(v;11q23.3) | 1 (5.0%) | 36 (3.5%) | 0 (0%) | 56 (3.1%) | ||
| Del(7)/‐7/abn(17p) | 2 (10.0%) | 14 (1.4%) | 0 (0%) | 30 (1.7%) | ||
| Del(5q) | 1 (5.0%) | 1 (.1%) | 0 (0%) | 5 (.3%) | ||
| Del(9q) | 1 (5.0%) | 19 (1.8%) | 0 (0%) | 30 (1.7%) | ||
| Trisomy | 1 (5.0%) | 57 (5.5%) | 1 (4.0%) | 112 (6.4%) | ||
| Other abnormal karyotype | 5 (25.0%) | 141 (13.7%) | 0 (0%) | 242 (13.8%) | ||
| ND | 0 (0%) | 64 (6.2%) | 1 (4.0%) | 81 (4.6%) |
- —Tian Jin Natural Science Foundation
- —CAMS Innovation Fund for Medical Sciences
- —National Natural Science Foundation of China 10.13039/501100001809
- —National Key Research and Development Program of China 10.13039/501100012166
- —Beijing Xisike Clinical Oncology Research Foundation 10.13039/100018904
- —Haihe Laboratory of Cell Ecosystem Innovation Fund
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAcute Myeloid Leukemia Research · Chromatin Remodeling and Cancer · Acute Lymphoblastic Leukemia research
1
To the Editor:
Acute myeloid leukaemia (AML) is a diverse and complex category of malignant disease with poor outcomes. Despite advancements in prognostication and treatment strategies, the molecular landscape of AML remains complex and is not fully understood. Epigenetic factors are acknowledged as crucial in tumour development and progression.1 The KMT2 (histone‐lysine N‐methyltransferase 2) family encompasses six key proteins: KMT2A/B, KMT2C/D, KMT2F and KMT2G, which is most notably associated with AML.2 KMT2A is associated with KMT2A‐rearranged leukaemia.3 KMT2B is identified as a hotspot for rearrangements.4 KMT2C/D mutations occurred frequently in various malignancies.5 However, the clinical characteristics of KMT2C/D mutations in AML remain poorly defined. We found that KMT2C and KMT2D mutations are relatively rare and mutually exclusive in newly diagnosed AML, with KMT2C mutations enriched in CEBPA‐mutated and KMT2D in NPM1‐mutated AML subtypes, respectively. In general, *KMT2C^MUT^
- and *KMT2C^WT^ *, as well as *KMT2D^MUT^
- and *KMT2D^WT^
- AML, exhibit distinct mutational spectrums, similar clinical characteristics and survival outcomes.
We reviewed 1935 AML patients who underwent next‐generation sequencing (NGS) analyses between 2015 and 2024. Of these, 1050 were eligible for KMT2C analysis and 1777 for KMT2D analysis. The KMT2C mutation rate was 1.90% (20/1050), consistent with previous studies.6, 7 The KMT2D mutation rate was 1.41% (25/1777), lower than that reported in a small‐sample study.8 No patient had concurrent KMT2C and KMT2D mutations. Characteristics of AML patients with and without KMT2C/D mutation are presented in Table 1. Clinical characteristics did not differ significantly between wild‐type and KMT2C/D‐mutated AML patients, except that *KMT2D^WT^
- patients had higher haemoglobin levels than *KMT2D^MUT^
- patients (*p *< .001). Among KMT2C mutated patients, nine (45%) were male and their median age was 43.9 years. For AML classification, 10 of 20 patients were classified as AML with CEBPA mutation. Patients with KMT2D mutations included 11 (44%) males, with a median age of 42.0, and 11 (44%) were recognised as AML with NPM1 mutation. Patients with KMT2C and KMT2D mutations were enriched by AML with CEBPA and NPM1 mutations, respectively. Additionally, most patients had normal karyotypes, including 50% of *KMT2C^MUT^
- patients and 72% of *KMT2D^MUT^
- patients.
We identified 22 mutation sites in the KMT2C gene (Figure 1A) and 30 in the KMT2D gene (Figure 1B). In all 95.5% of KMT2C mutations were nonsense and frameshift, consistent with the COSMIC database analysis by Rao et al.5 The PHD domain was the most frequently mutated region in KMT2C, with 27.3% (6/22) of mutations located there, of which 83.3% (5/6) were nonsense. In contrast, KMT2D mutations were dispersed without clustering in specific exons or domains. We also analysed co‐mutations in KMT2C/D mutant patients. A total of 110 (median five mutations per patient) and 128 (median five mutations per patient) mutations had been found in KMT2C and KMT2D mutated AML, respectively. All KMT2C/D mutations were accompanied by additional gene mutations. CEBPA (n = 10, 50.0%) was the most frequent co‐mutation with KMT2C, followed by NRAS (n = 7, 35.0%), GATA2 (n = 5, 25.0%), FLT3 (n = 5, 25.0%). Garg et al. reported that KMT2C co‐mutates with FLT3.9 In addition, compared with *KMT2C^WT^
- AML, *KMT2C^MUT^
- AML had significantly higher frequencies of CEBPA (50.0% vs. 20.6%, p = .004), GATA2 (25.0% vs. 6.1%, *p *= .007) and BCOR (20.0% vs. 4.9%, p = .016) mutations (Figure 1C). Besides, all CEBPA mutations were *CEBPA^bZIP^
- (Figure 1D). Correspondingly, FLT3 (n = 13, 52.0%) was the most frequent co‐occurred mutation in *KMT2D^MUT^
- AML, followed by NPM1 (n = 11, 44.0%) and IDH2 (n = 7, 28.0%). Compared with *KMT2D^WT^
- AML, *KMT2D^MUT^
- patients were more likely to co‐occur with FLT3 (52.0% vs. 28.1%, p = .016) and NPM1 (44.0% vs. 19.2%, p = .004) mutations (Figure 1E,F).
KMT2C/D mutations in AML. The distribution of KMT2C (A) and KMT2D (B) mutations, which were identified in our cohort, on the protein. Bar chart of additional gene mutation distribution in KMT2CWT and KMT2CMUT groups (C), and additional gene mutation distribution in KMT2DWT and KMT2DMUT groups (E), sorted according to the mutation rate in a mutated group, and the percentage of each gene mutation is shown. (D) Mutation oncoplot of the 20 AML patients with KMT2C mutation. (F) Mutation oncoplot of the 25 AML patients with KMT2D mutation.
Further, we compared the characteristics of different mutation statuses of KMT2C/D in the *CEBPA^bZIP^
- (Table S1) and NPM1 (Table S2) subgroups, respectively. In both subgroups, clinical characteristics were similar between wild‐type and KMT2C/D‐mutated AML patients, except for the higher haemoglobin in *KMT2D^WT^
- patients (p = .035). In the *CEBPA^bZIP^
- AML cohort, *KMT2C^MUT^
- AML were more likely to have co‐mutations of NRAS (50.0% vs. 21.1%, p = .05) than *KMT2C^WT^ *. In the NPM1 ^MUT^ subgroup, the prevalence of IDH2 co‐mutations was significantly higher in *KMT2D^MUT^
- AML compared with *KMT2D^WT^
- AML (63.6% vs. 18.7%, p = .002).
Finally, we examined whether KMT2C/D mutations influence the prognosis of AML. There was no difference in overall survival (OS) or event‐free survival (EFS) between *KMT2C^WT^
- and *KMT2C^MUT^
- AML patients. The 1‐year OS rates were 83.8% and 94.7% for *KMT2C^WT^
- and *KMT2C^MUT^
- groups (HR:. 69, 95% confidence interval [CI]:. 22–2.15, p = .52; Figure 2A), respectively. The 1‐year EFS rate in *KMT2C^WT^
- and *KMT2C^MUT^
- groups were 50.5% and 63.8% (HR:. 64, 95% CI:. 32–1.28, p = .2; Figure 2B), respectively. Although no difference in OS was observed between *CEBPA^bZIP^ */*KMT2C^WT^ *and *CEBPA^bZIP^ */*KMT2C^MUT^
- patients (Figure 2C), the *CEBPA^bZIP^ */*KMT2C^MUT^
- patients exhibited superior EFS, achieving 1‐year EFS rate of 90.0% in contrast to 55.4% in the *CEBPA^bZIP^ */*KMT2C^WT^
- patients (HR:. 15; 95% CI:. 02–1.06, p = .028; Figure 2D). Multivariate analysis also demonstrated that KMT2C mutation in *CEBPA^bZIP^
- patients was associated with better EFS (HR:. 102, 95% CI:. 013–.766, *p *= .026) but not OS (Table S3). *KMT2D^MUT^
- and *KMT2D^WT^
- groups exhibited similar outcomes with 1‐year OS rates at 81.5% and 88.4% (HR:. 78, 95% CI:. 29–2.09, p = .62; Figure 2E), 1‐year EFS rate at 49.5% and 49.0% (HR: 1.09, 95% CI:. 62–1.93, p = .76; Figure 2F), respectively. Moreover, for *NPM1^MUT^
- patients, KMT2D mutation did not affect EFS or OS, as shown by Kaplan–Meier survival analysis (Figure 2G,H) and multivariate analysis (Table S4). Additionally, we investigated the impact of KMT2C/D mutations on survival outcomes across different NCCN risk groups, yet found no significant effects in the favourable, intermediate or adverse risk groups (Figures S1 and S2). However, the effect of KMT2C/D on the prognosis of AML needs to be further explored and verified by more studies with larger samples.
The overall survival (OS) (A) and event‐free survival (EFS) (B) of KMT2CWT and KMT2CMUT groups in our cohort, and OS (C) and EFS (D) of KMT2CWT and KMT2CMUT groups in the CEBPAbZIP subgroup. The OS (E) and EFS (F) of KMT2DWT and KMT2DMUT groups in our cohort, and OS (G) and EFS (H) of KMT2DWT and KMT2DMUT groups in the NPM1MUT subgroup.
AUTHOR CONTRIBUTIONS
HW and JW participated in concept design. MY, XZ, and WW were involved in data collection and analysis, drafting and revising the manuscript. JC, SQ, BL, and YM were responsible for interpreting the results. All authors have read and approved the final manuscript.
FUNDING INFORMATION
National Key Research and Development Program of China, Grant Number: 2023YFC2508900; National Natural Science Foundation of China, Grant Number: 82370183; CAMS Innovation Fund for Medical Sciences, Grant Number: 2023‐I2M‐2‐007; Tian Jin Natural Science Foundation, Grant Number: 23JCZXJC00310; Haihe Laboratory of Cell Ecosystem Innovation Fund, Grant Number: 22HHXBSS00040; Beijing Xisike Clinical Oncology Research Foundation, Grant Number: Y‐SYBLD2022ZD‐0031
ETHICS STATEMENT
This research was approved by the ethical committee in the Institute of Hematology and Blood Diseases Hospital, and all procedures were in accordance with the Helsinki Declaration. Written informed consent was obtained from each participant.
Supporting information
Supporting Information
Supporting Information
Supporting Information
Supporting Information
Supporting Information
Supporting Information
Supporting Information
Supporting Information
Supporting Information
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Baylin SB , Jones PA . A decade of exploring the cancer epigenome—biological and translational implications. Nat Rev Cancer. 2011;11(10):726‐734.21941284 10.1038/nrc 3130 PMC 3307543 · doi ↗ · pubmed ↗
- 2Van HT , Xie G , Dong P , Liu Z , Ge K . KMT 2 family of H 3K 4 methyltransferases: enzymatic activity‐dependent and ‐independent functions. J Mol Biol. 2024;436(7):168453.38266981 10.1016/j.jmb.2024.168453 PMC 10957308 · doi ↗ · pubmed ↗
- 3Ziemin‐van der Poel S , Mc Cabe NR , Gill HJ , et al. Identification of a gene, MLL, that spans the breakpoint in 11q 23 translocations associated with human leukemias. Proc Natl Acad Sci U S A. 1991;88(23):10735‐10739.1720549 10.1073/pnas.88.23.10735 PMC 53005 · doi ↗ · pubmed ↗
- 4Huntsman DG , Chin SF , Muleris M , et al. MLL 2, the second human homolog of the Drosophila trithorax gene, maps to 19q 13.1 and is amplified in solid tumor cell lines. Oncogene. 1999;18(56):7975‐7984.10637508 10.1038/sj.onc.1203291 · doi ↗ · pubmed ↗
- 5Rao RC , Dou Y . Hijacked in cancer: the KMT 2 (MLL) family of methyltransferases. Nat Rev Cancer. 2015;15(6):334‐346.25998713 10.1038/nrc 3929 PMC 4493861 · doi ↗ · pubmed ↗
- 6Dolnik A , Engelmann JC , Scharfenberger‐Schmeer M et al. Commonly altered genomic regions in acute myeloid leukemia are enriched for somatic mutations involved in chromatin remodeling and splicing. Blood. 2012;120(18):E 83‐E 92.22976956 10.1182/blood-2011-12-401471 · doi ↗ · pubmed ↗
- 7Ley TJ , Miller C , Ding L , et al. Genomic and epigenomic landscapes of adult de novo acute myeloid leukemia. N Engl J Med. 2013;368(22):2059‐2074.23634996 10.1056/NEJ Moa 1301689 PMC 3767041 · doi ↗ · pubmed ↗
- 8Meena JP , Pathak N , Gupta AK , et al. Molecular evaluation of gene mutation profiles and copy number variations in pediatric acute myeloid leukemia. Leuk Res. 2022;122:106954.36162216 10.1016/j.leukres.2022.106954 · doi ↗ · pubmed ↗