Expression of MHC I Isoforms in Bovine Placentomes: Impact of Cloning
Rodrigo da Silva Nunes Barreto, Ana Carolina Furlanetto Mançanares, Maria Angelica Miglino, Flávio Vieira Meirelles, Lilian de Jesus Oliveira

TL;DR
This study shows that cloned cow pregnancies have altered immune-related protein expression in the placenta, which may explain higher rates of pregnancy loss.
Contribution
The study reveals differences in MHC-Ib protein and gene expression patterns between cloned and non-cloned bovine pregnancies.
Findings
Qa-2 protein expression in cloned pregnancies was intense at both early and late stages, unlike in controls.
ILA-88 labeling was weak in early cloned pregnancies but increased near term, contrasting with controls.
BoLA-NC3 gene was significantly upregulated in cloned pregnancies during early and near-term gestation.
Abstract
The immune system protects the body from infections and distinguishes between self and foreign components. During pregnancy, the mother’s immune system recognizes and tolerates the foreign paternal proteins present in the fetus. When there is a maladjustment of the maternal immune system during pregnancy, it may result in maternal rejection and pregnancy loss. The fetus also helps regulate the maternal immune system by producing placenta-specific proteins called non-classical major histocompatibility complexes (MHC-Ib). This study examined how MHC-Ib proteins differ between non-cloned (control) and cloned cow pregnancies since cloned pregnancies have more complications and losses. Using tissue staining and gene expression analyses, we found that in control pregnancies, early expression of a specific MHC-Ib (Qa-2) was barely present but increased near calving, whereas Qa-2 had an…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
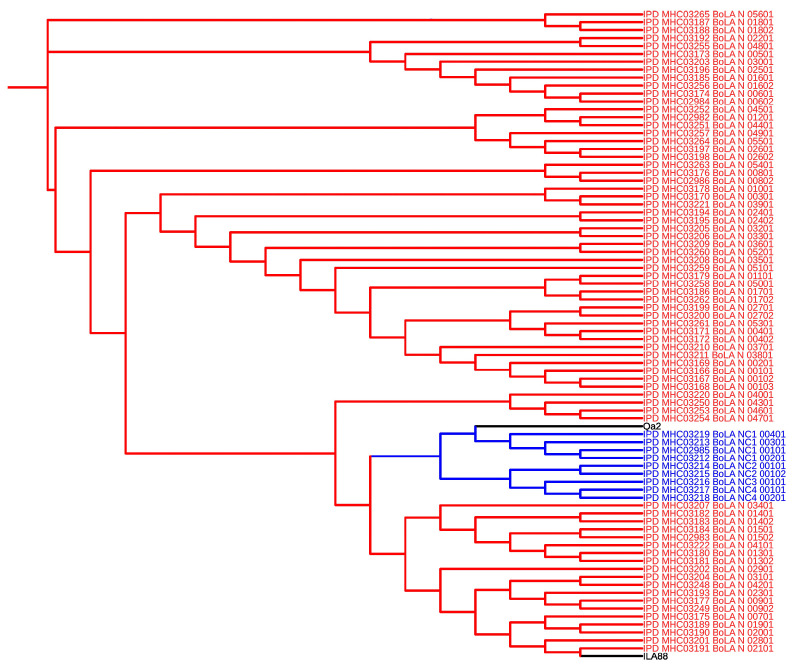 Figure 1
Figure 1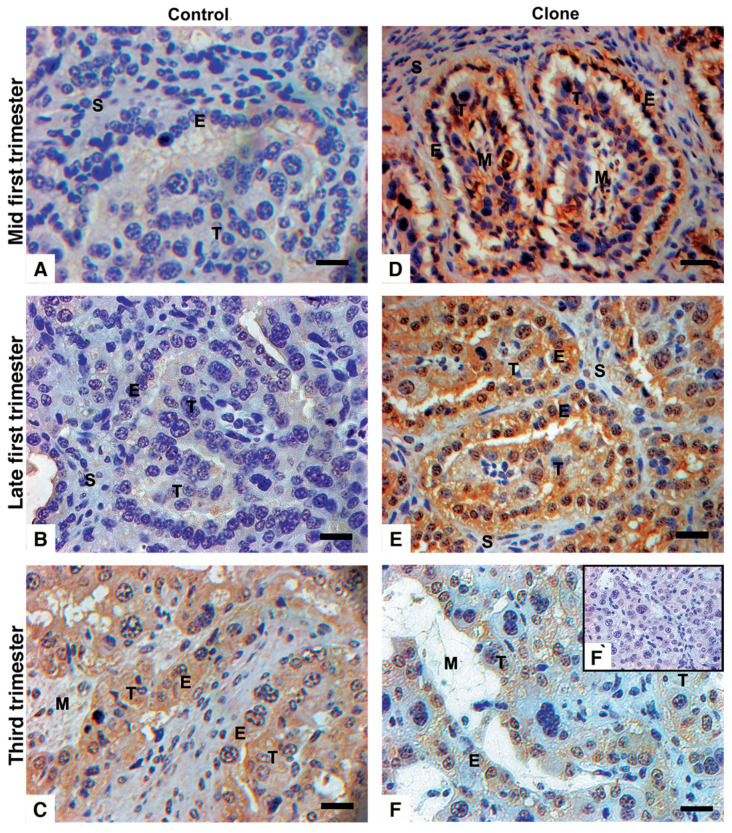 Figure 2
Figure 2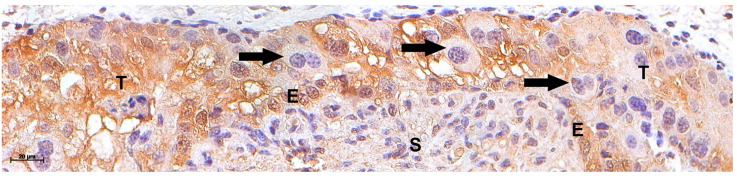 Figure 3
Figure 3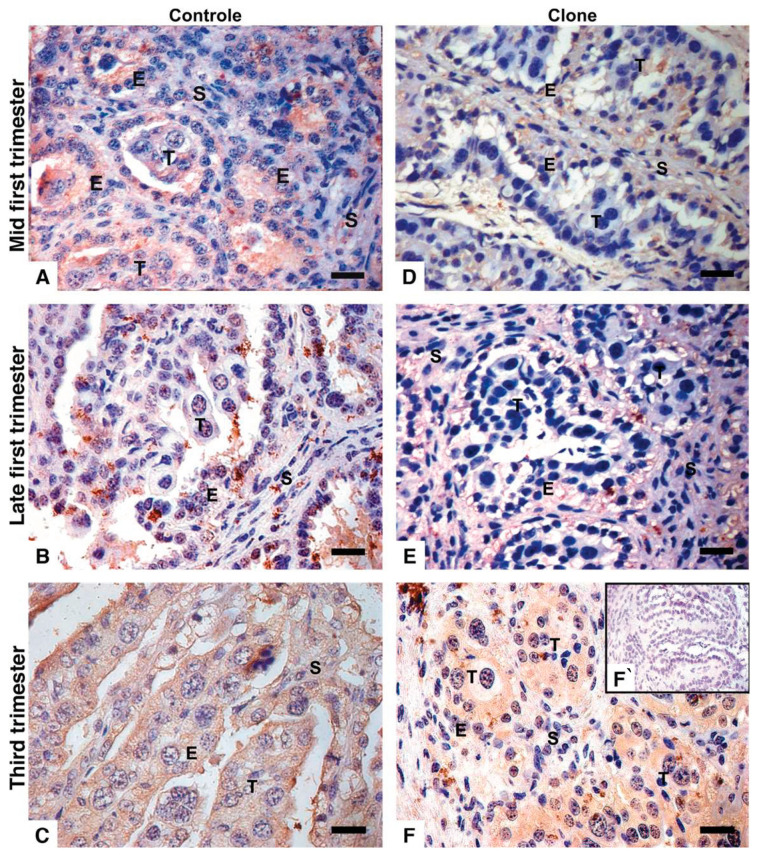 Figure 4
Figure 4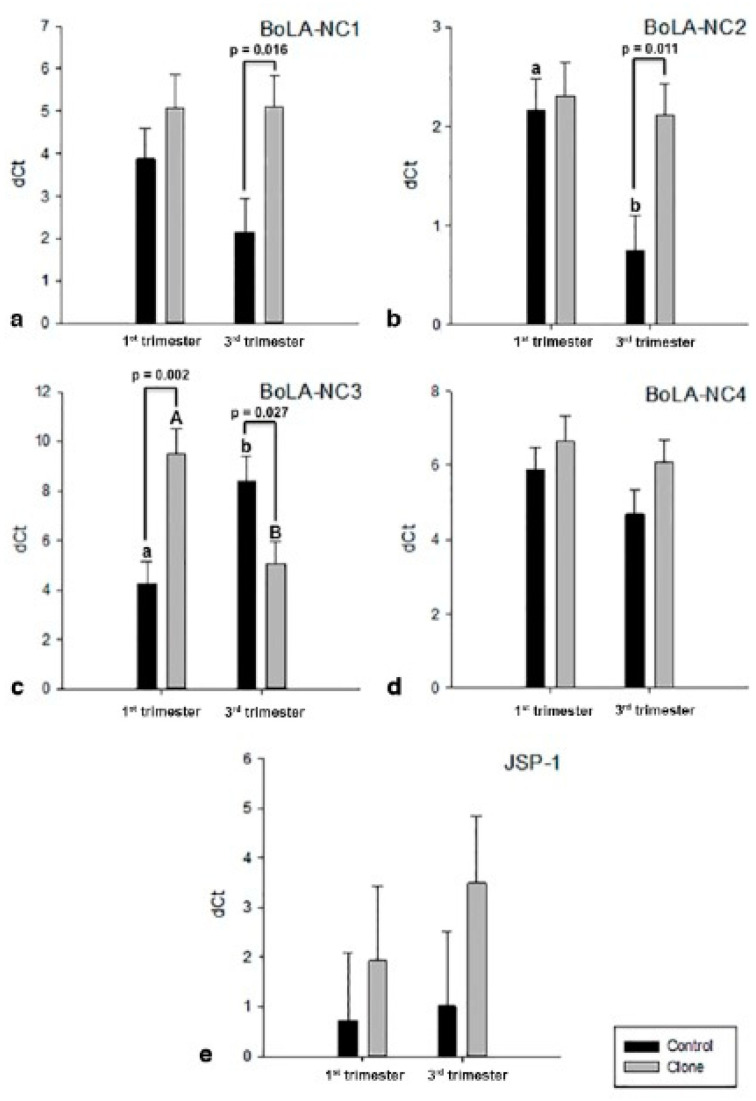 Figure 5
Figure 5- —CAPES
- —FAPESP
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsReproductive System and Pregnancy · Reproductive Physiology in Livestock · Reproductive Biology and Fertility
1. Introduction
During pregnancy, the communication between mother and fetus occurs through the expression and transfer of placental-specific factors [1], such as the fetal expression of major histocompatibility complex (MHC) proteins [2]. In hemochorial placental models, such as in mice and humans, semi-allogeneic trophoblastic cells (fetal cells) decrease the expression of MHC-I molecules early in gestation to avoid the recognition of paternal antigens by the maternal immune system [3]. With the low to no expression of MHC-I, the fetal cells may become a target of uterine natural killer (uNK) cells [4]. To prevent activation of uNK cells, the trophoblast cells express a unique form of MHC molecules: The non-classical (NC) MHC-I isoforms [5], described as human leukocyte antigen G (HLA-G) or murine preimplantation embryo development (PED) genes [2]. NC MHC-I genes are expressed in isoforms: membrane-bound or soluble proteins [6]. Soluble HLA-G is detected in the maternal blood in successful pregnancies, suggesting a systemic modulation of the maternal immune system [7]. High expression of HLA-G is associated with an invasive phenotype of the trophoblast cells in vitro, the extravillous trophoblasts (EVTs) [8]. However, trophoblast invasion and adhesion to the decidua are regulated not only by HLA-G expression but also through interactions with inhibitory receptors on NK cells and macrophages, such as KIR2DL4/CD158d, LILRB1/2, and ILT2/4 [8,9]. HLA-G expression by the EVT modulates the local maternal immune system through the uNK cell system and its interactions with macrophages and other immune cells. Regulatory T (Treg) cells play an essential role in supporting the semi-allogeneic fetus in successful pregnancies [10], inhibiting T cell-mediated lysis of trophoblastic cells, and promoting a supporting immune environment [11]. Abnormal expression of HLA-G appears deleterious: its downregulation correlates with reduced numbers of CD56^bright^CD16^−^ uNK cells in the maternofetal interface, which can contribute to miscarriages [12]. An abnormal invasion of the EVTs and uNK cells can impair the development and remodeling of maternal spiral arteries, predisposing intrauterine to restrict growth and pre-eclampsia [13]. For example, when the EVTs from mid-gestation samples are cultured in conditioned uNK cell media, their ability to invade is significantly increased in vitro [14]. Moreover, the co-culture of the HLAG^+^ EVTs and CD4^+^ cells increases the expression of forkhead box P3 (FOXP3) by CD4^+^CD25^high^ T reg cells, supporting the HLA-G effects on the Th immune response at the maternofetal interface [10].
In contrast to humans and mice, bovines display a less invasive or epitheliochorial type of placentation in which all maternal tissue layers are maintained throughout pregnancy [14,15]. Unlike other species with epitheliochorial placenta, the ruminant placenta is unique and appears more permeable to fetal products [16]. In cows, the trophoblastic giant cells (TGCs) migrate toward the maternal epithelium and form a hybrid syncytium with the maternal epithelium [16] to deliver fetal products to the maternal system [17]. Despite maintaining all tissue layers in both the dam and fetus sides, the dam immune system displays similar local and systemic modulation as observed in humans and mice. Such include increased circulating CD4^+^CD25^high^ T reg cells in early pregnancy [18] and alternative activation of uterine macrophages near-term [19] The more intimate maternofetal communication in cows primes the maternal system to semi-allogeneic fetal antigens [20]. In contrast to humans and mice, MHC-I or bovine leukocyte antigen class I (BoLA-I) expression in early gestation is unknown. In cows, trophoblasts in the arcade zones and interplacentomal chorionic regions express BoLA-I around 6 months into the pregnancy; meanwhile, BoLA-I expression is absent deep in the chorionic villi [21,22,23,24]. This suggests that the suppression of BoLA-I expression in bovine placentomes occurs in areas of close contact between fetal cells and the maternal epithelium [21,22,23,25]. In clones, BoLA-I is expressed as early as day 36 in the chorioallantois membrane, which later differentiates into chorionic villi as the placentome development progresses [25]. This suggests that BoLA-I expression in early pregnancy may represent a non-classical isoform of BoLA class I (BoLA-NC), like murine PED and human HLA-G [26]. Furthermore, atemporal and increased expression of BoLA-I observed in cloned pregnancies may have contributed to abnormal pregnancy/placentation and losses in the early stages of pregnancy [8,27].
The expression patterns of classical and non-classical isoforms of BoLA class I molecules remain uncharacterized, and their role in local maternal–fetal immune modulation remains unclear. In this study, we describe the expression of classical and non-classical MHC class I molecules in bovine placentomes from both cloned and naturally conceived pregnancies, examined during early and near-term stages of gestation.
2. Materials and Methods
For immunohistochemistry (IHC), mouse Qa-2 (clone 69H1-9-9, #11-5996; eBioscience, San Diego, CA, USA) and mouse bovine monomorphic MHC (clone IL-A88, #MCA2444GA, AbDSerotec, Kidlington, Oxford, UK) were used as primary antibodies. For control, we used anti-mouse IgG (clone MOPC-21, #M5284; Sigma-Aldrich, Saint Luis, MO, USA). Reactions were tagged with Dako Advance tm HRP (#K4069) and stained using 3,3-diaminobenzidine (DAB, Liquid DAB+ Substrate Chromogen System, #K346811), both purchased from DAKO (Carpinteria, CA, USA).
For real-time quantitative PCR (RT-qPCR), an RNeasy Plus Mini kit (#74136, Qiagen—Valencia, CA, USA) was employed for RNA extraction, DNAse I amplification grade (#18068-015), alongside RNAseOut Recombinant Ribonuclease Inhibitor (#10777-019), High-Capacity cDNA Reverse Transcription kit with RNase Inhibitor (#4374967), and SyBr Green PCR master mix (#4344463), all purchased from Invitrogen (Foster City, CA, USA). Primers were purchased from Exxtend (Paulínia, SP, Brazil).
Monohydrate citric acid, sodium citrate tribasic dehydrate, sodium chloride (NaCL), hydrogen peroxide (H_2_O_2_), potassium chloride (KCl), Trizma base (C4H11NO3), and 3-(Aminopropyl) triethoxysilane for buffer solutions were purchased from Sigma-Aldrich.
2.1. Clustering of BoLA Isoforms and Sequences Recognized by Qa-2 and IL-A88 Antibodies
Published classical and non-classical BoLA isoform protein sequences were retrieved from the IPD-MHC database (https://www.ebi.ac.uk/ipd/mhc/allele/list/?query=eq(organism.group,BoLA)#panelAdvanced, accessed on 8 January 2025) and peptide sequence recognized by the anti-Qa-2 (GenBank: X80933.1) and anti-IL-A88 (GenBank: NM_001201460.1) antibodies were retrieved in FASTA format for further analysis. Phylogenetic analyses were conducted using Clustal Omega (https://www.ebi.ac.uk/jdispatcher/msa/clustalo, accessed on 8 January 2025) [28] within a Singularity container to ensure reproducibility and control over computational resources. Sequences were aligned using the dealign option to eliminate pre-existing gaps, ensuring alignments based solely on sequence data. The software utilized MBED-like clustering for both guide-tree construction and iterative refinements to enhance alignment precision. The output sequences were maintained in their aligned order, and default settings were used for combined iterations, maximum guide tree, and HMM iterations, balancing efficiency, and detailed analysis.
2.2. In Silico Analysis of Bovine Classical and Non-Classical MHC Proteins Solubility
SignalP 4.1 (https://services.healthtech.dtu.dk/services/SignalP-5.0/, accessed on 10 January 2025) was used to predict secretion via the classical secretory pathway through the presence of signal peptides; SecretomeP 2.0 Server (https://services.healthtech.dtu.dk/services/SecretomeP-2.0/, accessed on 10 January 2025) was used to predict non-classical secretory pathways. Only the entries that were predicted to be secreted by the SignalP 4.1 or SecretomeP 2.0 Servers were used for further analyses.
The secreted predicted sequences were further analyzed by TMHMM Server v. 2.0 (https://services.healthtech.dtu.dk/service.php?TMHMM-2.0/, accessed on 10 January 2025) for the presence of transmembrane helices. Furthermore, to confirm the secretory nature of the analyzed sequences, two web tools were used to predict the subcellular location of predicted secreted proteins. Meanwhile, TargetP 1.1 Server (https://services.healthtech.dtu.dk/service.php?TargetP-2.0, accessed on 10 January 2025which utilizes neural networks to predict the subcellular location of a given protein sequence, was also used. A similar analysis was performed using WoLF PSORT (http://www.genscript.com/wolf-psort.html, accessed on 10 January 2025), which predicts the protein subcellular location using a dataset of 12,771 animal proteins obtained from UniProt (http://www.uniprot.org/, accessed on 10 January 2025) version 45 annotated by Gene Ontology (http://geneontology.org/, accessed on 10 January 2025).
2.3. Placentome Samples
Placentome samples from bovine embryos produced by somatic cell nuclear transfer (SCNT) pregnancies [29] were collected after surgical uteri recovery, and control placentomes were obtained from a local abattoir. For both groups, bovine uteri were incised in the pregnant horn antimesometrial line for placentae assessment. Then, membranes, endometrium, and fetuses were morphologically analyzed. For controls, we only used placentomes derived from pregnancies without gross abnormalities in vasculature, membranes, and placentome number and size. However, for clones, it was known that some placental abnormalities could be present [30,31,32], and only ones without exacerbated abnormalities were used. Placentomes were investigated from clones at early gestation (60 and 90 days; n = 5) and near-term (285 ± 0 days; n = 6), as well as from controls at early gestation (72.8 ± 10.6 days; n = 6) and near term (251 ± 21 days; n = 6); these were determined according to their crown–rump length [33]. For RT-qPCR, 16 samples were included, grouped into the following: early gestation (75 ± 17.3 days; n = 4) and near term (285 days; n = 4) for clones; early gestation (73 ± 16.2 days; n = 4) and near term (240 ± 7.4 days; n = 4) for controls. The research was approved by the Ethical Committee of the College of Veterinary Medicine and Animal Science of the University of Sao Paulo (protocol number: 9408110315).
2.4. Immunohistochemistry Using Qa-2 and IL-A88 Antibodies
Samples were fixed in 4% buffered paraformaldehyde, dehydrated, and paraffin-embedded. Sections of 5 µm were mounted on precoated slides, deparaffinized, and rehydrated. Antigen retrieval was performed by microwaving the slides 4 times in high potency citrate buffer (1.83 mM of monohydrate citric acid and 8.9 mM of sodium citrate tribasic dehydrate, pH 6.0) for 5 min. Endogenous peroxidase was blocked using 3% hydrogen peroxide in distilled water in the dark for 1 h. Nonspecific protein interactions were blocked using 10% of normal goat serum in Tris-buffered saline (TBS: 2.0 mM of Trizma base and 1.36 mM of sodium chloride, pH 7.5) solution for 30 min, followed by incubation with specific primary antibodies Qa-2 (1:50), IL-A88 (1:50) or irrelevant anti-mouse IgG in a humid chamber at 4 °C overnight. The reaction was detected using Dako Advance tm HRP, and the color was developed after applying DAB. The slides were counter-stained with hematoxylin. Between each step, slides were rinsed in TBS 3 times for 5 min, and after antibody incubation, the slides were rinsed in TBS containing 1% normal goat serum. The slides were analyzed using an upright Zeiss Axioplan 2 microscope and Axiocan HRc camera (Zeiss, Jenna, DE).
2.5. Detection of MHC Gene Expressions by RT-qPCR
The RNA from all samples was extracted using the RNeasy Plus Mini kit; DNA digestion was performed using amplification grade DNAse I, and cDNA conversion was conducted using the High-Capacity cDNA Reverse Transcription kit, all following the manufacturer’s instructions. For each real-time qPCR reaction, 7.5 μL of SyBr Green PCR master mix, 1.0 mM of each primer (Table 1) [34,35,36,37,38], 1.0 μL of diluted cDNA (63.0 μL of ultrapure water and 1.0 μL of cDNA), and ultrapure water were used to a total volume of 15.0 μL. The default cycle program from the 7500 fast real-time PCR system (Invitrogen) was used. Data were analyzed according to the dCt method from Livak et al. [39], and statistical analysis was performed using least square means analysis of variance with the GLM procedure of SAS (version 5.1). The model included main effects (group and animal) and standard errors. Graphs were plotted on SigmaPlot software (version 12.0).
3. Results
3.1. Qa-2 and IL-A88 Antibodies Recognize Putative Soluble MHC-I Proteins
The sequence alignment of BoLA isoforms and specific peptides recognized by Qa-2 and IL-A88 antibodies revealed that the Qa-2 antibody predominantly clustered with non-classical bovine MHC I isoforms, while the IL-A88 antibody clustered with classical isoforms, as illustrated in the generated guide tree (Figure 1). In addition in silico prediction of soluble motifs of bovine MHC I isotypes can be accessed in Table S1.
3.2. Immunohistochemistry with Distinct Patterns Between Qa-2 and IL-A88 Antibodies
In early gestation, controls showed no labeling for the Qa-2 antibody in all tissues of the placentomes (Figure 2A,B). Comparatively, strong staining against Qa-2 antibody stain was observed in near-term uninucleate trophoblasts and the maternal uterine epithelium. In contrast, the bi-nucleated trophoblast was weakly stained, and the mesenchyme and endometrial stroma were largely negative (Figure 2C). In clones, trophoblasts and the maternal epithelial cells were strongly labeled against Qa-2 antibody in early gestation (Figure 2D,E) but only weakly stained near term (Figure 2F). The Qa-2 antibody labeling was stronger in the arcade zones than in the villous areas in all positive cases. In addition, TGCs that were closely apposed to or fused with maternal epithelial cells showed weak to no labeling for Qa-2 (Figure 3).
In controls, at 60 days of gestation, both trophoblast and maternal epithelial cells exhibited moderate labeling for IL-A88 antigen (Figure 4A). By day 90 of gestation, while the labeling intensity of IL-A88 in maternal cells remained consistent, it was markedly decreased in the trophoblast cells (Figure 4B). Near-term, all samples displayed faint to weak staining for IL-A88 (Figure 4C). In clone samples, the IL-A88 antibody showed mild to moderate labeling, which was observed only in maternal cells during early gestation (Figure 4D,E). Moreover, in the near term, the IL-A88 antibody labeling intensity was evident in both the trophoblast and maternal epithelium (Figure 4F). Additionally, the TGCs that were closely apposed or fused with the maternal epithelium exhibited markedly weak to no labeling for IL-A88, similar to the staining patterns observed with the Qa-2 antibody.
3.3. Expression and Regulation of MHC I Gene Isoforms
Among the four non-classical MHC I isoforms investigated (Figure 5a–e), BoLA-NC3 exhibited significant differences in expression throughout pregnancy between the control and cloned groups (p < 0.01). In the controls compared to the clones, BoLA-NC3 expression was upregulated in early gestation (p < 0.01) and downregulated near term (p = 0.03) (Figure 5c). Expression of BoLA-NC1 differed only in the near term, showing downregulation in controls relative to clones (p < 0.02) (Figure 5a). Similarly, BoLA-NC2 expression differed only in the near term, being downregulated in controls compared to clones (p < 0.02). Additionally, controls exhibited a decrease in BoLA-NC2 expression near term compared to early gestation (p < 0.01), whereas its expression remained constant throughout pregnancy in clones (Figure 5b). In contrast, the expression of BoLA-NC4 and the classical isoform JSP-1 did not show significant differences at any analyzed stages of pregnancy or between groups (Figure 5d,e).
4. Discussion
In contrast to previously known, MHC-I isoforms were detected in both control and clone bovine placentomes throughout pregnancy at mRNA and protein levels. In our study, two different antibodies for immunohistochemistry were tested, and the antibody–antigen reaction was revealed with a biotin-free polymer-based secondary antibody system. Such second-generation antibody systems are more sensitive and specific [40] than the formerly used biotinylated ones [22,23,26,28,36,37]. We concluded that newer, more sensitive reagents are more suitable for detecting MHC I expression, particularly during early bovine gestation when the MHC class I protein expression is evident in our current work. It is possible that the antigen retrieval and blockage of nonspecific site methods previously required by older reagents may have disrupted the integrity of the antigen–antibody binding process, potentially altering epitope availability, reducing binding affinity, or increasing background signal, thereby affecting assay specificity and sensitivity.
We observed intense labeling in uninucleate trophoblasts compared to TGCs. All the TGCs composing the syncytium with the maternal epithelium exhibited a complete loss of labeling. A similar pattern of heterogeneous MHC-I expression in trophoblasts has been described in humans [41,42]. Only the migratory EVT cells express HLAG to suppress the activation of uterine NK cells and resident macrophages. The EVT interacts with maternal immune cells, hence HLAG expression. In contrast, syncytiotrophoblast near the maternal blood system has only light expression of soluble HLAG and does not express membrane-bound HLAG [43]. Thus, comparable to humans, we suppose that the fused cells at the fetal–maternal interface were also characterized in the bovine by suppressed MHC I expression. This may promote vulnerability regarding the maternal immune system, which may be involved in the process of apoptosis of the syncytia [44] and the release of their products into the maternal system [16]. The strong staining on the maternal side and putative soluble pattern of BoLA isoforms suggest the presence of (a) membrane-bound, classical MHC I as specific for epithelium and (b) soluble isoforms that must have been released by the trophoblast and perhaps “accumulated” in the uterine epithelium [45,46]. In humans, the maternal epithelium displays a similar “accumulation” of non-classical MHC I as observed in the bovine placenta. Even though the bovine placenta is generally less invasive, studies have proven the permeability of fetal products to the maternal system [17,47,48,49]. Recently, the cloned bovine endometrial immune response was shown to be regulated by trophoblast BoLA expression [50] and hypoxic conditions [51], which is a common feature in cloned pregnancies [52,53].
Our results highlight differences between the two antibodies used. In our in silico analysis, the Qa-2 sequence appeared closer to non-classical BoLA isoforms. Moreover, Qa-2 antigens were not detected in early gestation, with markedly increased labeling in the control samples in the near term. In contrast, the human placenta expresses HLA-G from the first trimester onwards [54], probably due to a more invasive placenta. In contrast, clones showed intense labeling in early gestation that decreased close to term, indicating that an altered overexpression of non-classical MHC I during early gestation may lead to pregnancy abnormalities, as evidenced by its expression by chorioallantoic membranes prior to placentome formation [25], which were even higher in MHC-I heterozygous incompatible clones [27]. In our in silico analysis, IL-A88 protein sequences appeared closer to classical MHC I proteins. In control samples, IHC for IL-A88 showed consistent labeling in the maternal epithelium at both early and later stages of gestation. In contrast, clones exhibited weak to no IL-A88 labeling in the maternal epithelium during early gestation but intense labeling in both maternal and fetal epithelium near term. These data suggest that IL-A88 may recognize the characteristics, i.e., classical antigens of epithelia, that become downregulated on the fetal side, similar to those previously observed for humans and mice [55,56]. However, in clones in the near term, the trophoblast is labeled for IL-A88 antigens. The anti-Qa-2 and anti-IL-A88 monoclonal antibodies differed in the pattern of labeling in the control and cloned samples; it could be a consequence of an abnormal chromatin remodeling due to abnormal patterns of methylation of the trophoblast during the preimplantation period that perpetuated during the placental development [57].
MHC I mRNA expression was detected in all bovine placentome samples throughout pregnancy, consistent with the isoform evaluations by Santos et al. [58]. Our findings reveal significant variations in the non-classical isoforms, with BoLA-NC3 being the most regulated gene during pregnancy. The expression patterns of this gene closely correlated with the immunohistochemistry results for Qa-2 antigens observed in this study. The abnormal Qa-2 labeling was especially aligned with non-classical isoform expression in cloned samples. In addition, BoLA-NC1 and BoLA-NC2 were significantly upregulated in the near-term stages of clones; however, the functional meaning remained unclear.
5. Conclusions
Our data suggest that MHC-I protein and gene expressions in the bovine placenta are similar to those observed in more invasive placenta types, such as in humans and mice, suggesting bovine epitheliochorial placentas possess similar mechanisms of trophoblast evasion from the maternal immune system during gestation. Controls and clones had distinct protein expression profiles for Qa-2 and IL-A88 antigens, which was somehow observed via differences in the expression of non-classical MHC-I molecules. The disparity in MHC-I regulation in clones may lead to a failure in proper pregnancy signaling, potentially leading to gestational failures.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zumkeller W. Current Topic: The Role of Growth Hormone and Insulin-like Growth Factors for Placental Growth and Development Placenta 20002145146710.1053/plac.2000.050510940195 · doi ↗ · pubmed ↗
- 2Rizzo R. Melchiorri L. Stignani M. Baricordi O.R. HLA-G Expression Is a Fundamental Prerequisite to Pregnancy Hum. Immunol.20076824425010.1016/j.humimm.2006.10.01217400059 · doi ↗ · pubmed ↗
- 3Moffett-King A. Natural Killer Cells and Pregnancy Nat. Rev. Immunol.2002265666310.1038/nri 88612209134 · doi ↗ · pubmed ↗
- 4Heemskerk B. Lankester A.C. van Vreeswijk T. Beersma M.F.C. Claas E.C.J. Veltrop-Duits L.A. Kroes A.C.M. Vossen J.M.J.J. Schilham M.W. van Tol M.J.D. Immune Reconstitution and Clearance of Human Adenovirus Viremia in Pediatric Stem-Cell Recipients J. Infect. Dis.200519152053010.1086/42751315655775 · doi ↗ · pubmed ↗
- 5Bulmer J.N. Johnson P.M. Antigen Expression by Trophoblast Populations in the Human Placenta and Their Possible Immunobiological Relevance Placenta 1985612714010.1016/S 0143-4004(85)80063-13895220 · doi ↗ · pubmed ↗
- 6Hunt J.S. Petroff M.G. Mc Intire R.H. Ober C. HLA-G and Immune Tolerance in Pregnancy FASEB J.20051968169310.1096/fj.04-2078 rev 15857883 · doi ↗ · pubmed ↗
- 7Steinborn A. Varkonyi T. Scharf A. Bahlmann F. Klee A. Sohn C. Early Detection of Decreased Soluble HLA-G Levels in the Maternal Circulation Predicts the Occurrence of Preeclampsia and Intrauterine Growth Retardation during Further Course of Pregnancy Am. J. Reprod. Immunol.20075727728610.1111/j.1600-0897.2007.00475.x 17362389 · doi ↗ · pubmed ↗
- 8Li C. Houser B.L. Nicotra M.L. Strominger J.L. HLA-G Homodimer-Induced Cytokine Secretion through HLA-G Receptors on Human Decidual Macrophages and Natural Killer Cells Proc. Natl. Acad. Sci. USA 20091065767577210.1073/pnas.090117310619304799 PMC 2667005 · doi ↗ · pubmed ↗