Development of an RT-qPCR Assay for the Detection of an Emerging Duck Egg-Reducing Syndrome
Zhifei Zhang, Xin Su, Dun Shuo, Dawei Yan, Xue Pan, Bangfeng Xu, Minghao Yan, Shuxuan Ren, Qinfang Liu, Chunxiu Yuan, Qiaoyang Teng, Zejun Li

TL;DR
Researchers developed a sensitive and specific RT-qPCR test to detect a virus causing reduced egg production in ducks, which could help control its spread and economic impact.
Contribution
A novel RT-qPCR assay was developed for the specific and sensitive detection of Duck Egg-Reducing Syndrome Virus (DERSV).
Findings
The RT-qPCR assay detected DERSV with a limit of 102 copies and a linear range from 102 to 109 copies per reaction.
The assay showed 92.59% efficiency and less than 2% coefficient of variation for both intra- and inter-assays.
Among 153 clinical samples, 47.06% tested positive for DERSV using the developed RT-qPCR method.
Abstract
Duck egg-reducing syndrome virus (DERSV), a novel Avihepatovirus, causes a gradual decline in duck egg production. Early detection of infected ducks is essential for preventing the spread of the virus and minimizing its economic impact. This study developed a quantitative reverse transcription PCR (RT-qPCR) assay for DERSV detection, which was validated for high sensitivity, specificity, and reliability. This method offers significant potential for use in epidemiological and pathogenesis studies. Duck egg-reducing syndrome virus (DERSV) is a novel Avihepatovirus and is responsible for a gradual decline in the laying rate of ducks, decreasing from a peak of 90% to 50%. The development of a rapid detection method for DERSV is crucial for the identification and control of virus infections. In this study, we developed a quantitative reverse transcription PCR (RT-qPCR) assay for detecting…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
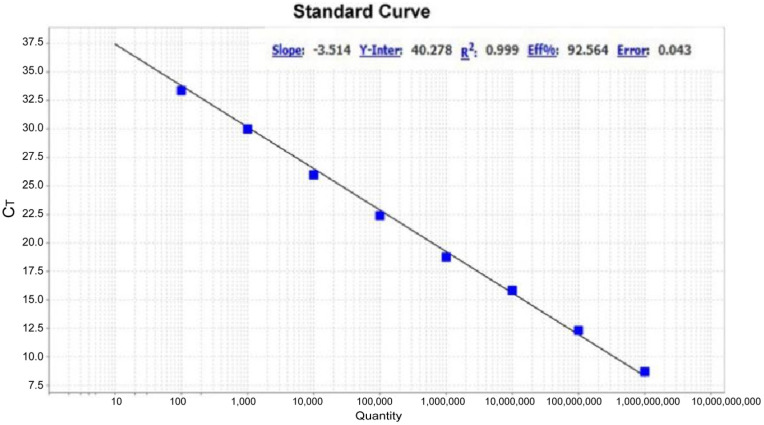 Figure 1
Figure 1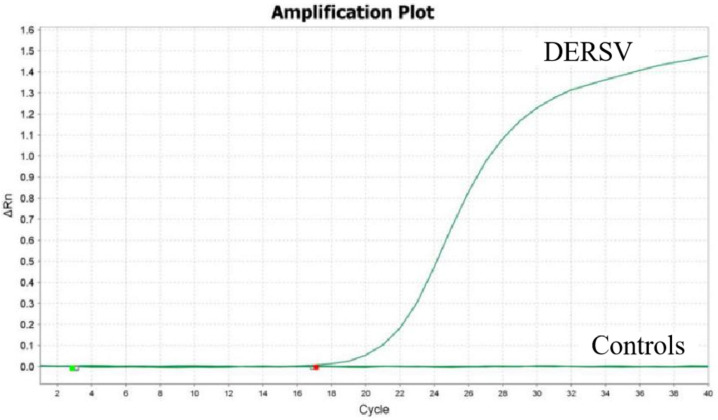 Figure 2
Figure 2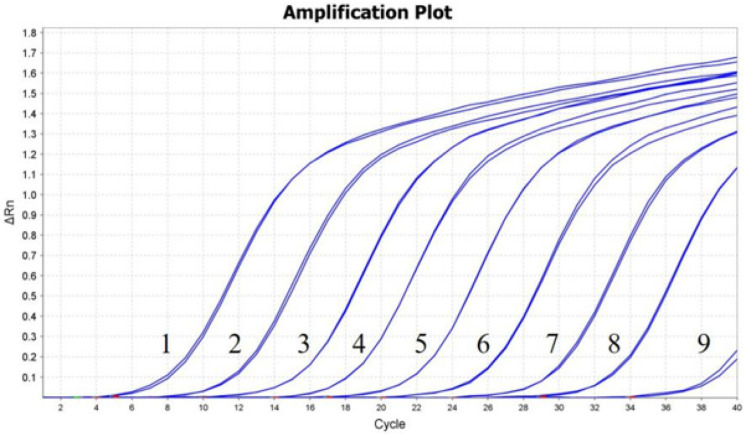 Figure 3
Figure 3- —Shanghai Agriculture Applied Technology Development Program
- —The National Key Research and Development Program of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsViral Infections and Immunology Research · Animal Virus Infections Studies · Animal Disease Management and Epidemiology
1. Introduction
Since 2016, frequent outbreaks of egg-reducing syndromes, linked to an unidentified virus, have caused significant economic losses in duck farms across China. In 2022, Su et al. identified the virus as a new species within the Avihepatovirus genus of the picornavirus family, naming it the duck egg-reducing syndrome virus (DERSV) [1]. Picornaviruses are small, spherical, non-enveloped positive-stranded RNA viruses [2]. The genomes range from 7000 to 10,000 nucleotides in length. The RNA is linked to a small peptide at the termination of the 5′ untranslated region (UTR) and a poly (A) tail at the end of the 3′ UTR [3]. The genome contains a single long open reading frame (ORF) encoding a polyprotein, and viral proteases processed it into structural and nonstructural proteins [4]. As of early 2022, the Picornaviridae family consists of 63 officially recognized genera, with the family rapidly expanding [5]. Many picornaviruses are significant pathogens in both humans and animals, including hand-foot-and-mouth disease, poliomyelitis, and various respiratory infections, causing diseases that affect the central nervous system, respiratory and gastrointestinal tracts, liver, pancreas, heart, eyes, and skin [6,7]. Until now, the picornaviruses that have an important impact on birds include duck hepatitis A virus (DHAV) [8] and avian encephalomyelitis virus (AEV) [9].
As a novel picornavirus, DERSV infection is primarily characterized by a gradual decline in laying rate, dropping from a peak of 90% to 50%. The laying rate eventually recovers after some time, and infected ducks do not die. The virus is transmitted through direct contact among ducks. Infected individuals exhibit liver and kidney hemorrhage, follicular hemorrhage, and follicular rupture. Additionally, viral antigens are detectable in the liver, ovaries, and kidneys, where pathological changes are observed [1].
Real-time PCR is a well-established approach for quantification and detection of virus. To date, several real-time PCRs have been established, and the most commonly used real-time PCR chemistries are TaqMan probes and SYBR green [10]. TaqMan real-time PCR could template quantification and high-throughput screening with high sensitivity and precision [11], so it has been used in many clinical laboratories for virus pathogenesis studies and epidemiological investigations [12,13,14]. We have established conventional RT-PCR and nested RT-PCR of DERSV, but they are cumbersome and time-consuming, which also lacks quantitative assays. The aim of this study was to develop a TaqMan probe-based RT-qPCR assay targeting the 3D gene of DERSV to quantify viral load. The assay’s applicability was evaluated for detecting DERSV in clinical samples.
2. Materials and Methods
2.1. Viral Strains and Clinical Samples
DERSV (strain AH204), duck Tembusu virus (DTMUV), H9N2 influenza virus (H9N2 AIV), duck reovirus (DRV), and duck hepatitis viruses type 1 and 3 (DHAV-1 and DHAV-3) were isolated and stored in our laboratory. A total of 153 archived tissue homogenates (from brain, pharynx, lung, heart, proventriculus, liver, spleen, pancreas, oviduct, follicles, and colon) were collected from sick ducks in Shandong province between 2016 and 2023.
2.2. Primers and Probe Design
The gene sequences of DERSV strains were referenced from GenBank and aligned using DNASTAR software (DNASTAR, Madison, WI, USA). Since the 3D gene is highly conserved among DERSV strains, it was chosen as the target for RT-qPCR analysis. Primers and a probe were designed using the IDT online server (Table 1) and were synthesized by GENEWIZ (Suzhou, China).
2.3. RNA Extraction and cDNA Synthesis
Viral RNA was extracted using the TIANamp Virus RNA Kit (Tiangen, Beijing, China) following the kit’s protocol, and viral RNA was reverse transcribed into cDNA using Vazyme M-MLV (H^−^) Reverse Transcriptase (Vazyme, Suzhou, China) according to the manufacturer’s guidelines. The RNA and cDNA samples were stored at −80 °C until further use.
2.4. Construction of Standard Plasmid
A part of the DERSV 3D gene was amplified using the primers mentioned before. The PCR product was embedded into the pMD19-T vector with the Ampicillin resistance gene and transformed into DH5α E. coli cells. Clones containing the 3D gene were sequenced using M13 F/R primers. The recombinant pMD19T-3D plasmid was purified using the TIANprep Mini Plasmid Kit (Tiangen, Beijing, China) according to the kit’s protocol, and the concentrations were quantified using an ND-2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). The recombinant plasmid’s copy numbers were calculated according to the methods described previously [15]. The pMD19T-3D plasmid was 10-fold serially diluted in TE buffer (10 mmol/L Tris-HCl, 1 mmol/L EDTA) to concentrations ranging from 10^9^ to 10^1^ copies/μL and stored at −80 °C until further use.
2.5. RT-qPCR Assay
The concentrations of the primers and probe were optimized using a matrix approach. RT-qPCR was performed in a 20 μL reaction mixture containing 10 μL of 2 × PCR buffer and 1 μL template. Primers (10 μmol/L), probe (10 μmol/L), and sterile water were added to the reaction with different volumes to optimize the assay. Amplification and detection were carried out using an ABI Q5 instrument (Applied Biosystems, Foster City, CA, USA) following these conditions: 95 °C for 2 min, followed by 40 cycles of amplification at 95 °C for 20 s and 54 °C for 1 min.
2.6. Specificity Analysis
The specificity analysis was performed using viral cDNA. The RT-qPCR assay was performed to amplify pathogens including DTMUV, H9N2 AIV, DRV, DHAV-1, and DHAV-3.
2.7. Sensitivity and Repeatability of the RT-qPCR Assays
The plasmid pMD19T-D3, with concentrations ranging from 10^9^ to 10^1^ copies/μL, was used for the sensitivity assay. To assess the repeatability of the RT-qPCR, 10^7^, 10^6^, and 10^5^ copies/μL of pMD19T-D3 were tested in quintuplicate on five separate occasions to calculate the coefficient of variation (CV). Both intra- and inter-assay CVs for Ct values were determined.
2.8. Detection of the Clinical Sample
A total of 153 clinical samples were collected from ducks suffering from duck egg-reducing syndrome. Viral RNA was extracted and detected using the developed RT-qPCR assay. The RT-PCR method we have established [16] was used to test these samples, and the RT-PCR products were sequenced (GENEWIZ, Suzhou, China). The clinical samples were also analyzed for the presence of DTMUV [17], AIV [18], DHAV [19], Duck enteritis virus (DEV) [20], and DRV [21].
3. Results
3.1. Optimization for the RT-qPCR
The optimal concentrations of primers and probe, which resulted in the highest fluorescence and the lowest threshold cycle, were determined as follows: 0.4 μmol/L for both primers and 0.2 μmol/L for the probe. The optimized reaction system was a 20 μL reaction mixture containing 10 μL of 2 × PCR buffer, 0.8 μL of each primer (10 μmol/L), 0.4 μL of probe (10 μmol/L), and 1 μL of template.
3.2. Standard Curve
The pMD19T-3D plasmid was 10-fold serially diluted in TE buffer, and a standard curve was generated from 10^2^ to 10^9^ copies (Figure 1). The assay demonstrated linearity over a 10^8^ dilution range of template DNA, with a reaction efficiency of 92.59% and an R^2^ value of 0.999.
3.3. Specificity of the RT-qPCR Assay
Using the viral cDNA of DTMUV, H9N2 AIV, DRV, DHAV-1, and DHAV-3 as a template, only DERSV was detected by RT-qPCR while other viruses were not detected (Figure 2), demonstrating the assay was high specifically.
3.4. Sensitivity and Repeatability of the RT-qPCR Assay
A dilution series of the pMD19T-3D plasmid, ranging from 10^9^ to 10^1^ copies/μL, was used as the template. The RT-qPCR assay demonstrated a detection limit of 10^2^ copies (Figure 3). Three independent replicates were conducted, all yielding consistent results. The intra-assay CVs of 10^7^, 10^6^, and 10^5^ copies of pMD19T-3D were 0.27%, 0.38%, and 0.44%, and the inter-assay CVs were 0.32%, 1.85%, and 0.88%, respectively (Table 2).
3.5. Clinical Sample Test Results
Of the 153 samples evaluated, 72 were positive for DERSV RNA, resulting in a positivity rate of 47.06% (72/153). Ten samples were positive for AIV and negative for DERSV. None of the samples exhibited detectable levels of DTMUV, DHAV, DEV, or DRV. Among the 72 DERSV positive samples, the liver, spleen, lungs, and proventriculus exhibited positive results in over 40% of the specimens. The Ct values for these samples ranged from 22.27 to 29.66. The samples positive for DERSV by RT-PCR were confirmed by sequencing of the amplicons.
4. Discussion
DERSV is a recently discovered virus belonging to the Avihepatovirus genus within the Picornaviridae family, first detected in ducks in 2016. Its pathogenic mechanisms and molecular epidemiology remain unexplored. Although we have developed RT-PCR and nested RT-PCR methods for detecting DERSV, these methods are time-consuming and do not allow for detailed quantitative analysis. In contrast, real-time PCR has become widely accepted due to its speed, simplicity, and reproducibility, while also minimizing the risk of carry-over contamination compared to conventional PCR [22].
Currently, a variety of qPCR methods have been developed for the detection of viruses belonging to the Picornaviridae family. Huang, Qiuxue et al. established a real-time quantitative PCR assay for detecting duck hepatitis A virus, and the limit of detection was 3.36 × 10^3^ copies [23]. Li, Yeqiu et al. developed a duplex RT-PCR assay for the detection of duck hepatitis A virus (DHAV-1) and duck astrovirus type 3 (DAstV-3), which is a valuable tool for the detection of coinfection [24]. Liu, Qingtian et al. developed a SYBR Green based RT-qPCR assay for avian encephalomyelitis virus detection, and the sensitivity was 100 times more compared with the conventional RT-PCR method [25]. All these studies show that real-time PCR is a valuable technique for clinical diagnosis and detection.
In this study, we developed a TaqMan-based RT-qPCR assay for detecting DERSV infection, which demonstrated high sensitivity, specificity, and good repeatability. The results showed that the standard curve exhibited a linear correlation (R^2^) of 0.999 and an efficiency of 92.56%. The assay exhibited no cross-reactivity with 6 duck-derived pathogens. In future studies, additional pathogens, such as Circovirus and Paramyxovirus, should be examined to evaluate the specificity and establish a foundation for the development of multiplex RT-qPCR. The detection limit for DERSV was 10^2^ copies/μL, and the intra-assay and inter-assay coefficients of variation were 0.44% and 1.85%, respectively. DERSV was detected primarily in the lung, proventriculus, liver, and kidney, suggesting that these organs should be prioritized in clinical investigation. These findings demonstrate that this qPCR assay is a reliable and reproducible platform.
To date, we have isolated 9 strains of DERSV from different provinces, which exhibit high amino acid similarity and certain nucleotide variations [1]. These strains are capable of infecting different duck strains and show a trend of gradual spread. Additional sample collection is essential to monitor the spread and evolution of DERSV, providing critical data to support the development of effective strategies for its prevention and control.
5. Conclusions
In conclusion, the RT-qPCR assay developed for detecting DERSV RNA is sensitive, specific, and reliable. This assay could serve as a valuable tool for diagnosis, epidemiological, and pathogenesis studies.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Su X. Shuo D. Luo Y. Pan X. Yan D. Li X. Lin W. Huang D. Yang J. Yuan C. An Emerging Duck Egg-Reducing Syndrome Caused by a Novel Picornavirus Containing Seven Putative 2A Peptides Viruses 20221493210.3390/v 1405093235632674 PMC 9144743 · doi ↗ · pubmed ↗
- 2Lizcano-Perret B. Michiels T. Nucleocytoplasmic Trafficking Perturbation Induced by Picornaviruses Viruses 202113121010.3390/v 1307121034201715 PMC 8310216 · doi ↗ · pubmed ↗
- 3Zhang X. Paget M. Wang C. Zhu Z. Zheng H. Innate Immune Evasion by Picornaviruses Eur. J. Immunol.2020501268128210.1002/eji.20204878532767562 · doi ↗ · pubmed ↗
- 4Haji Zamani N. Hosseini H. Ziafati Kafi Z. Sadri N. Hojabr Rajeoni A. Esmaeelzadeh Dizaji R. Molouki A. Fallah Mehrabadi M.H. Abdoshah M. Ghalyanchilangeroudi A. Whole-Genome Characterization of Avian Picornaviruses from Diarrheic Broiler Chickens Co-Infected with Multiple Picornaviruses in Iran Virus Genes 202359799010.1007/s 11262-022-01938-036239871 · doi ↗ · pubmed ↗
- 5Zell R. Delwart E. Gorbalenya A.E. Hovi T. King A.M.Q. Knowles N.J. Lindberg A.M. Pallansch M.A. Palmenberg A.C. Reuter G. ICTV Virus Taxonomy Profile: Picornaviridae J. Gen. Virol.2017982421242210.1099/jgv.0.00091128884666 PMC 5725991 · doi ↗ · pubmed ↗
- 6Zell R. Picornaviridae—The Ever-Growing Virus Family Arch. Virol.201816329931710.1007/s 00705-017-3614-829058149 · doi ↗ · pubmed ↗
- 7Holmes A.C. Semler B.L. Picornaviruses and RNA Metabolism: Local and Global Effects of Infection J. Virol.201993 e 02088-1710.1128/JVI.02088-17PMC 680326231413128 · doi ↗ · pubmed ↗
- 8Kim M.-C. Kwon Y.-K. Joh S.-J. Lindberg A.M. Kwon J.-H. Kim J.-H. Kim S.-J. Molecular Analysis of Duck Hepatitis Virus Type 1 Reveals a Novel Lineage Close to the Genus Parechovirus in the Family Picornaviridae J. Gen. Virol.2006873307331610.1099/vir.0.81804-017030865 · doi ↗ · pubmed ↗