Spatiotemporal Diffusion, Colonization, and Antibody Responses in Susceptible C57BL/6J Mice Orally Infected with Toxoplasma gondii Cysts
Zhao Li, Qi-Shuai Liu, Jun-Jie Hu, Cai-Qin Deng, Tao Li, Wen-Bin Zheng, Xing-Quan Zhu, Feng-Cai Zou

TL;DR
This study tracks how Toxoplasma gondii spreads in mice, colonizes organs, and triggers immune responses after oral infection.
Contribution
The study provides a detailed spatiotemporal analysis of T. gondii infection dynamics and antibody responses in C57BL/6J mice.
Findings
T. gondii rapidly spreads to multiple organs and breaches the blood–brain barrier within 7 days.
Toxoplasma-specific IgG antibodies increase and remain stable for two months.
The brain is the primary predilection site for T. gondii cysts, leading to chronic meningitis.
Abstract
Toxoplasma gondii is an important zoonotic parasite infecting humans and the majority of other warm-blooded animals. The C57BL/6J mouse is susceptible to T. gondii infection and is considered an ideal model organism for T. gondii studies. This research explored the spatiotemporal dynamics of infection, colonization patterns, and antibody response fluctuations in C57BL/6J mice orally infected with T. gondii Type II strain cysts. The mice were orally infected with T. gondii cysts, and their clinical symptoms were monitored daily. The parasite load in various organs was assayed using qPCR targeting the T. gondii B1 gene. Serum antibody responses were assessed using ELISA. The cyst burden in the mouse brain was evaluated through histological analysis and immunofluorescence. T. gondii infection led to clinical manifestations in mice, such as fever and weight loss. The parasite rapidly…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
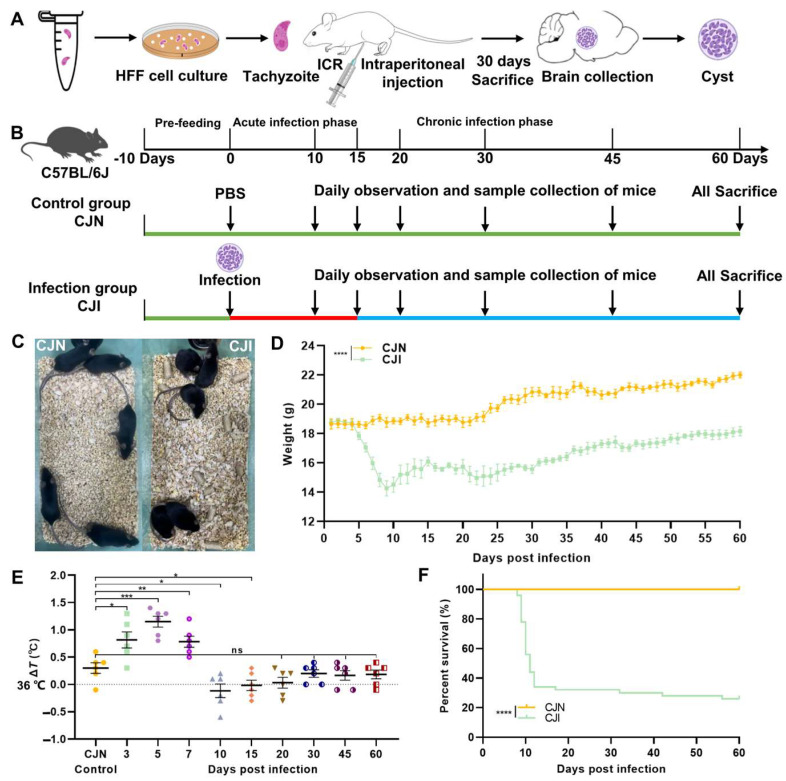 Figure 1
Figure 1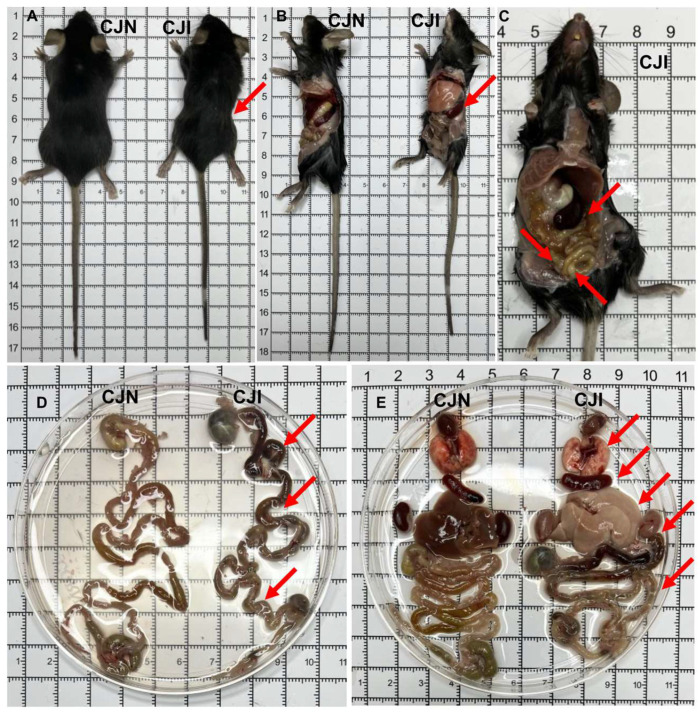 Figure 2
Figure 2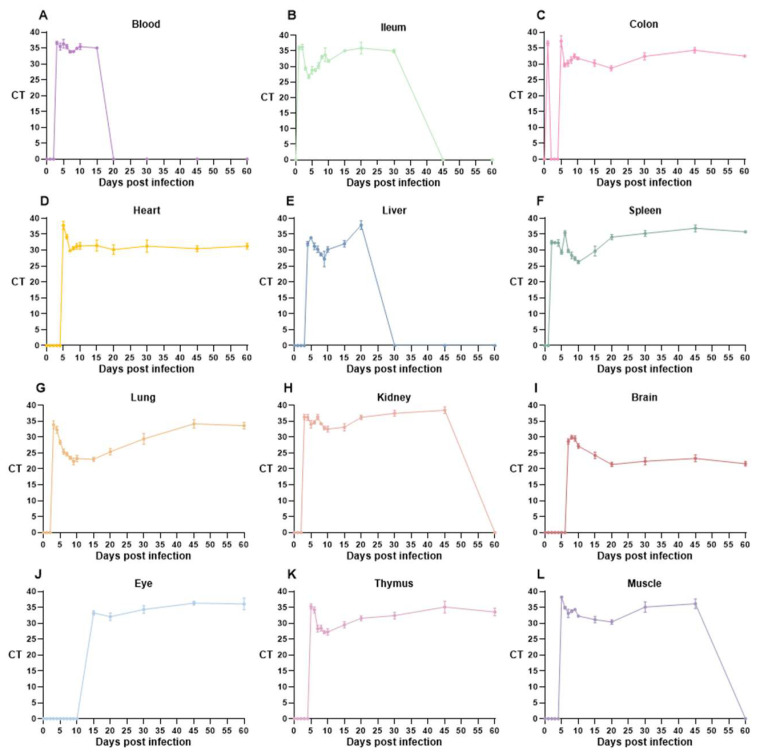 Figure 3
Figure 3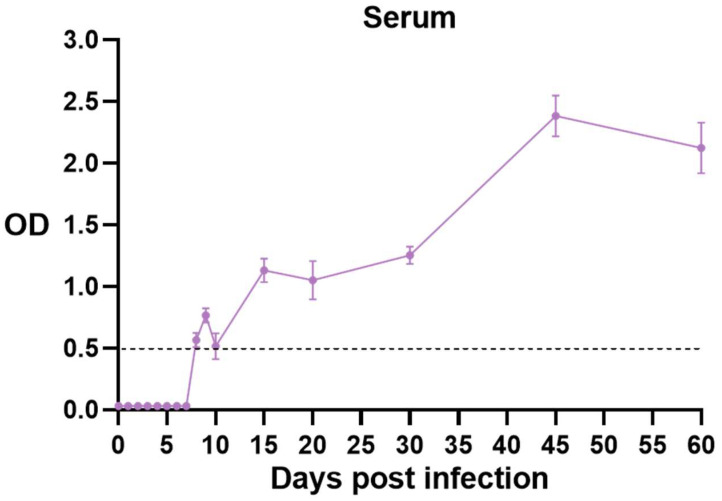 Figure 4
Figure 4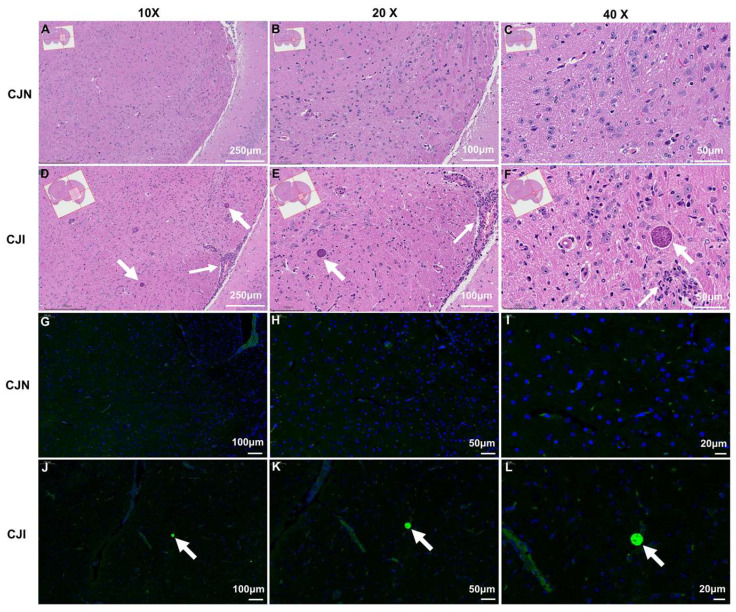 Figure 5
Figure 5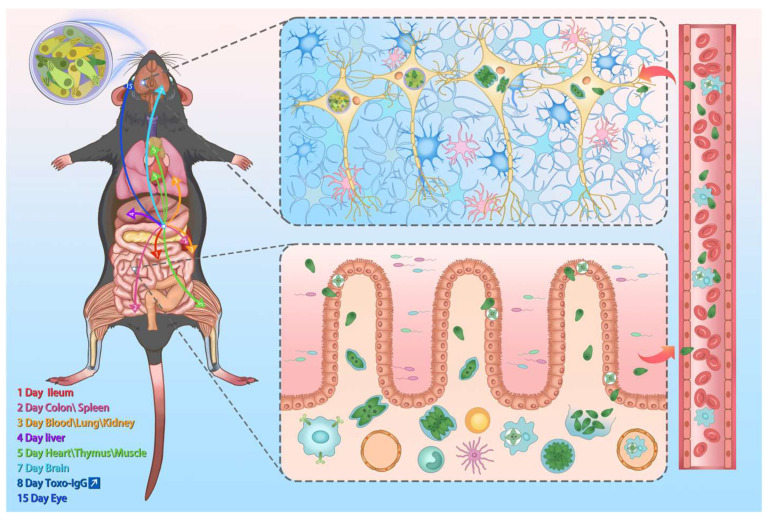 Figure 6
Figure 6- —the NSFC-Yunnan Joint Fund
- —the Yunnan Key Laboratory of Veterinary Etiological Biology
- —the Research Fund of Shanxi Province for Introduced High level Leading Talents
- —the Special Research Fund of Shanxi Agricultural University for High-level Talents
- —the National Natural Science Foundation of China
- —the Yunnan Fundamental Research Projects
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsToxoplasma gondii Research Studies · Cytomegalovirus and herpesvirus research · Herpesvirus Infections and Treatments
1. Introduction
Toxoplasma gondii is a single-celled zoonotic protozoan parasite that infects a wide range of warm-blooded animals, including humans, other mammals, and birds [1,2]. It is estimated to infect approximately one-third of the global population [3]. The parasite’s life cycle typically includes tachyzoite, bradyzoite, and oocyst stages, which vary across host types. In intermediate hosts such as humans, non-feline mammals, and birds, T. gondii reproduces asexually and forms tachyzoites during the acute phase of infection. When the infection progresses to a chronic stage, tachyzoites convert into bradyzoites, which persist within the host for its entire lifetime. In definitive hosts (e.g., cats and other felids), T. gondii undergoes sexual reproduction in the intestines, producing oocysts excreted in feces [4]. These oocysts are highly resistant to environmental conditions and can infect new hosts [5]. Additionally, definitive hosts may generate tachyzoites and bradyzoites. Dormant bradyzoites persist in host tissues, evade the immune system, and spread through the food chain to other intermediate and definitive hosts [6,7].
Humans acquire T. gondii infection primarily through the oral ingestion of undercooked meat containing cysts or by consuming food and water contaminated with oocysts. Mother-to-child transmission via the placenta and transmission via blood transfusion or organ transplant are also potential infection routes [8]. Infections among immunocompetent adults are often asymptomatic. Typical symptoms such as fever, fatigue, swollen lymph nodes, and muscle pain may manifest in some cases. Acute infections in healthy individuals usually resolve spontaneously, whereas immunocompromised individuals can experience severe complications, such as pneumonia and encephalitis [9]. This condition can also increase the risk of miscarriage or congenital malformations in pregnant women [10,11,12]. Additionally, acutely infected immunosuppressed patients (e.g., those with cancer or HIV/AIDS) are increasingly vulnerable to reactivation of T. gondii cysts, causing severe retinitis [13] and central nervous system inflammation [14]. T. gondii and toxoplasmosis constitute a concern for individual and public health, food safety, and ecological stability, emphasizing comprehensive research on prevention, control, and treatment [5].
The C57BL/6J mouse is considered an important model organism due to its stable genetic background, complete telomere-to-telomere reference genomes [15,16], advanced gene editing technologies [17], ease of breeding, and cost-effectiveness attributes [18]. The C57BL/6J mouse is susceptible to T. gondii infection and has many advantages over other laboratory animals. These mice have been commonly employed in research focusing on the virulence of T. gondii, the development of vaccines, interactions between pathogens and hosts, and food safety [19]. Despite these applications, systematic studies on the infection dynamics of the C57BL/6J mice after oral infection with T. gondii cysts remain lacking. Mice are conventionally categorized into outbred and inbred strains. Outbred strains include ICR (CD-1), KM, and NIH, while typical inbred strains are C57BL/6, BALB/c, C3H, DBA/2, FVB, and SJL [20]. KM mice are most widely used in China, whereas ICR mice are more popular internationally [21]. Studies have reported that different strains exhibit varying levels of susceptibility and pathogenicity to T. gondii. Inbred strains typically possess higher susceptibility than outbred strains [22]. Specifically, C57BL/6 mice are more susceptible than BALB/c mice and are more likely to develop severe, lethal enteritis [23,24]. Additionally, mice with the C57BL/KsJ (H2(d) haplotype) and CB10-H2 (H2(b) haplotype) backgrounds are particularly prone to congenital toxoplasmosis [25].
The infection stage (tachyzoites, cysts, and oocysts), contamination route (e.g., oral gavage, intraperitoneal injection, tail vein injection, and subcutaneous injection), and phase (acute or chronic) largely impact pathogenicity and immune responses in mice [26,27]. Further investigation is necessary to clarify the underlying mechanisms of pathogenesis and the dynamic immune responses in C57BL/6 mice infected via natural oral routes.
T. gondii is classified as a primary pathogen that should be excluded from specific pathogen-free laboratory mice. C57BL/6 mice are commonly used as the genetic background for genetically modified mice [17]. A T. gondii infection model in C57BL/6 mice is required to investigate immune responses and interactions during T. gondii infection.
This study explored the spatiotemporal diffusion, colonization patterns, and antibody response fluctuations in C57BL/6J mice infected orally with Type II T. gondii ME49 strain cysts [23]. These animals’ clinical signs, body weights, temperatures, and survival rates were monitored daily for 60 days post-infection (dpi). Parasite burden in various organs was quantified using qPCR targeting the T. gondii B1 gene. Serum antibody responses were assessed with enzyme-linked immunosorbent assay (ELISA). We also evaluated the brain cyst burden during chronic infection using histology and immunofluorescence.
2. Materials and Methods
2.1. Parasite Strains, Cell Culture Conditions, and Purification
The T. gondii strain TgME49 was maintained at 37 °C with 5% CO_2_ in confluent monolayers of human foreskin fibroblast cells following the method previously described [28]. Tachyzoites were harvested from highly infected human foreskin fibroblasts by mechanical release with a 27-gauge needle, followed by filtration through a 5 µm polycarbonate membrane filter to obtain pure parasites for subsequent mouse infection.
2.2. Mice and T. gondii Infection
Female ICR and C57BL/6J mice aged 6–8 weeks were raised under specific pathogen-free conditions at the Animal Research and Resource Center of Yunnan University (Approval Nos. SCXK-Yunnan-K2021-0001, SYXK-Yunnan-K2021-0002, and CNAS LA0029). The housing conditions conformed to the national standards: an environmental temperature of 22 ± 1 °C, relative humidity of 50 ± 5%, and 12 h light/dark cycle. Mice were provided with an irradiated and sterilized diet (JXT Standard) and had ad libitum access to filtered sterile water. Each ICR mouse was intraperitoneally injected with 500 T. gondii tachyzoites, and their weight changes, diet, and water intake were monitored. T. gondii tachyzoites were expected to colonize the brain tissue and form cysts after 30 dpi. The mice were euthanized at this stage, and brain tissues were dissected and homogenized. The number of cysts was counted under a microscope for subsequent oral infection of C57BL/6J mice. The C57BL/6J mice were randomly divided into the blank control group (CJN, n = 10) and the T. gondii infected group (CJI, n = 50). All the animals were randomly grouped, labeled, and acclimatized for ten days before the experiment. Each mouse in the CJN group received an oral gavage of 200 µL of phosphate-buffered saline (PBS). In contrast, each CJI mouse was gavaged with 100 T. gondii cysts in a total volume of 100 µL. The grouping protocols, gavage infection, and sample collection are presented in Figure 1A.
2.3. Clinical Observation and Sample Collection of Mice
The mice’s diet, drinking habits, and activity levels were continuously monitored before and throughout the 60-day experiment. Body weight was measured using an electronic scale. The rectal temperature was recorded with a probe thermometer every three days. Commercial nutritional jelly (Ready Jelly^®^ Recovery, Shenzhen, Chian) was administered promptly when the weight loss was greater than 15%. Weight loss exceeding 25% was established as the humane endpoint for euthanasia to ensure animal welfare. Mice were euthanized using a carbon dioxide instrument (30–70% concentrations). A clinical examination was conducted on the dead body and fur surface after euthanasia. The animals’ abdominal and thoracic cavities were dissected macroscopically, focusing on the intestines, spleen, liver, lungs, and heart. The samples included whole blood, ileum, colon, heart, liver, spleen, lungs, kidneys, brain, eyes, thymus, and hind leg muscle. Mouse sera and tissues (including the brain) were collected for immunological and histopathological analyses under specified conditions. Infected mice were monitored for 60 consecutive dpi. During monitoring, tissue sampling and analysis were performed at various intervals to assess infection progression and immune responses.
2.4. Detection of T. gondii with Real-Time q-PCR
Qualitative and quantitative analyses of the invasion, dissemination, proliferation, and colonization of T. gondii tachyzoites and cysts in different mouse tissues and organs were qualitatively and quantitatively analyzed using qPCR techniques. DNA was extracted from the tissue homogenate, and Toxoplasma nucleic acid was detected by targeting the T. gondii B1 gene using qPCR [29]. A TaqMan qPCR system targeting the T. gondii B1 locus (GenBank: AF179871) was applied with forward (TOXO-F: 5′-TCCCCTCTGCTGGCGAAAAGT-3′) and reverse (TOXO-R: 5′-AGCGTTCGTGGTCAACTATCGATTG-3′) primers paired with a dual-labeled (6FAM-TAMRA) hydrolysis probe. Genomic DNA was isolated from tissue homogenates using the Magnetic bead-based nucleic acid extraction kit (Sbeadex livestock kit, LGC, London, England) according to the manufacturer’s protocol. Amplification reactions (25 μL) comprising 12.5 μL of 2 × Premix Ex Taq (Probe qPCR) (TaKaRa, Dalian, China), 0.5 μL of each primer, 1 μL of probe, and 2 μL of template DNA were processed on a real-time PCR System (BioRad CFX96, Hercules, CA, USA). The thermal cycling parameters were set as follows: initial denaturation at 95 °C for 10 min, followed by 40 cycles of 95 °C for 15 s and 60 °C for 1 min. Real-time fluorescence monitoring enabled the automated determination of cycle threshold (Ct) values. C57BL/6J mice were orally inoculated with Toxoplasma cysts, with their tissues and organs being collected at specified periods post-infection (1–10 days, 15 days, 20 days, 30 days, 45 days, and 60 days). We analyzed the dynamics of T. gondii invasion, dissemination, proliferation, and colonization across different tissues and organs at each time point using the amplified CT values and the reference curve.
2.5. Serum T. gondii-Specific Antibodies
Serum was collected from mice at various durations following T. gondii infection to assess the levels of T. gondii-specific antibodies, specifically IgG. This measurement was performed using a commercially available ELISA kit (VRL Asia, Suzhou, China), following the manufacturer’s instructions. Each experiment included a positive control and a negative control to ensure validity. Specific criteria for a valid experimental design included an optical density (OD) of the negative control < 0.25 and a positive control OD ≥ 0.60. Invalid experiments were repeated until the defined criteria were met. The sample was considered positive when its OD value was ≥0.5 and negative when the corresponding OD was <0.5.
2.6. Histopathological Analysis
Brain tissue samples were collected from mice in the CJN and CJI groups 30 dpi and fixed in 4% (v/v) neutral-buffered formalin. These tissues were processed using standard histological techniques and embedded in paraffin to analyze the pathological lesions and distribution of T. gondii antigens. The paraffin sections were cut into 5-µm thick slices and stained with hematoxylin and eosin (H&E). Histopathological changes were evaluated using light microscopy (Olympus VS200, Tokyo, Japan). The methodology for pathological analysis was based on established protocols from previous research [30].
2.7. Immunofluorescence
Paraffin sections from the CJN and CJI mice were deparaffinized in water. The paraffin-free sections were subjected to antigen retrieval using an EDTA buffer (pH 8.0). These tissue sections were blocked with 10% bovine serum albumin at 37 °C for 30 min, and then the blocking buffer was removed. The resulting sections were incubated at 4 °C overnight with primary antibodies, specifically goat anti-T. gondii polyclonal antibody (1:1000, Invitrogen, Carlsbad, CA, USA). The incubated sections were washed three times with PBS and further incubated with secondary antibodies (1:500, Servicebio, Wuhan, China) at room temperature for 50 min. The 4′,6-diamidino-2-phenylindole was added and incubated at room temperature in the dark for 10 min to counterstain the cell nuclei. Finally, the sections were sealed with fluorescent mounting media for further analysis [31]. The stained sections were examined under a fluorescence microscope (Olympus VS200, Japan).
2.8. Statistical Analysis
GraphPad Prism 8.0.2 (GraphPad Software Inc., San Diego, CA, USA) was employed for analyses and graph creation. All the experiments were conducted at least three times, and the results are presented as the mean ± standard error (SEM). Body weight changes were determined based on two-way ANOVA. Cumulative mortality was plotted using Kaplan–Meier survival curves and analyzed with the Log-rank (Mantel–Cox) test. Differences between groups were assessed using Student’s t-test or the Mann–Whitney test, as indicated in figure legends. p < 0.05 was considered statistically significant. * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001 indicate a statistically significant difference, while ‘ns’ represents no significant differences.
3. Results
3.1. T. gondii-Infected Mouse Model and Analysis of Clinical Status
The cultivation of rapidly replicating T. gondii, the cyst preparation, and the experimental design for C57BL/6J mice are illustrated in Figure 1A, B. Approximately after 5 dpi with T. gondii cysts, notable changes in mice were observed, including reduced activity, rough fur, and a tendency to huddle in cage corners (Figure 1C). Daily clinical observations revealed a marked decrease in appetite and water intake at 3 dpi, accompanied by fever or an elevated body temperature, indicating the onset of the acute infection phase. The mice underwent an evident decline in body weight as of 5 dpi. In addition, the average body temperature decreased after peaking above 37 °C.
Mortality occurred roughly one week post-infection. The minimum body weight and temperature were observed by 10 dpi, leading to significant mortality rates (Figure 1D–F). The survivors transitioned from the acute phase to the chronic phase between ten and 15 dpi. Their conditions had improved by approximately two weeks post-infection. This transition was characterized by slight weight gain despite persistently lower body temperatures than normal levels.
The body weight of mice in the CJI group slowly increased and stabilized within 30–60 dpi. Despite this improvement, the temperature remained significantly lower than in the CJN group. Although the body temperature returned to normal levels, occasional deaths were still recorded among the infected mice (Figure 1D–F).
3.2. Oral Infection of Mice with T. gondii Cysts Induces Inflammatory Bowel Disease and Multi-Organ System Damage
Acutely infected mice that reached humane endpoints were immediately euthanized, followed by gross anatomical and pathological examination. Infected mice exhibited noticeable signs of emaciation and rough fur (Figure 2A). Gross anatomical analysis revealed significant splenomegaly in the abdominal cavity, accompanied by enlarged intestinal lymph nodes (Figure 2B,C). The surface areas of the jejunum and ileum displayed black discoloration during gastrointestinal tract dissection, indicating severe hemorrhage, tissue damage, and necrosis. Additionally, the cecum and colon were markedly reduced and shortened. Extensive damage and necrosis were observed throughout the intestinal tissue, leading to lethal enteritis (Figure 2D).
Comparative dissection further revealed that the hearts of T. gondii-infected mice in the acute infection phase were slightly shrunken, while their spleens were markedly enlarged. The lungs, liver, and kidneys exhibited a pale appearance, and petechial hemorrhages were observed on the surfaces of the lungs and kidneys (Figure 2E). Based on the above observations, T. gondii infection led to significant pathological damage and inflammation across multiple organs and systems in mice, resulting in multi-organ failure and death.
3.3. T. gondii Loads Vary Across Multiple Time Points and Various Organs
As mentioned, brain tissues of mice infected with T. gondii were homogenized, and qPCR analysis targeting the Toxoplasma B1 gene demonstrated an inverse relationship between dilution concentration and CT values. This analysis confirmed our method’s effectiveness in detecting the presence of T. gondii cysts. The qPCR method facilitates the qualitative and quantitative analysis of T. gondii load in infected mouse tissues with high specificity and sensitivity, laying a groundwork for follow-up detection applications.
Subsequently, we evaluated changes in CT values and tissue samples (blood, duodenum, ileum, heart, liver, spleen, lungs, kidneys, brain, eyes, thymus, muscle) across 15 intervals (1–10, 15, 20, 30, 45, and 60 dpi) using the qualitative and quantitative qPCR method targeting the Toxoplasma B1 gene (Figure 3). Qualitative analysis was initially performed using CT values, followed by quantitative analysis. The detection result was considered positive when 18 ≤ CT ≤ 38, with a lower CT value indicating a higher T. gondii load. Infection intensity is denoted as a ‘+’ symbol based on CT levels. The dynamic distribution of T. gondii across various tissues over time is detailed in Table 1.
Oral gavage of T. gondii cysts triggered the infection. The introduced cysts were broken down by gastric acid in the host stomach, releasing parasites into the gastrointestinal environment. During the acute phase of infection (1–2 dpi), parasites migrated to the duodenum and colonic tissues and rapidly spread to the spleen via the bloodstream. The proliferating T. gondii forms disseminated to the lungs, kidneys, and liver through the bloodstream between 3 and 4 dpi. We detected the presence of T. gondii in the heart, thymus, and leg muscles at 5 dpi. By 7 dpi, parasites breached the blood–brain barrier (BBB) and reached the brain tissues. T. gondii rapidly disseminated to most organs and tissues within one week following infection. The lungs of acutely infected mice exhibited significant parasite proliferation (Figure 3G and Table 1). The infection subsequently progressed into the chronic phase. T. gondii breached the blood–retinal barrier and invaded the eye tissues at 15 dpi. This parasite progressively colonized the brain and proliferated considerably (Figure 3I and Table 1). Key tissues, including the heart, brain, and leg muscles, shifted from rapidly proliferating forms (tachyzoites) to bradyzoites, initiating a dormant state as cysts. These cysts remain isolated from the host’s immune system and can persist for the host’s lifetime, facilitating transmission to intermediate and definitive hosts.
3.4. Changes in Toxoplasma-Specific Antibodies in Mouse Serum
Serum samples were collected at various dpi intervals, and T. gondii-specific IgG antibodies were quantified using an ELISA kit. The results revealed progressive changes in antibody titers (Figure 4). T. gondii IgG antibodies were detectable in C57BL/6J mice by the 8th dpi. The OD value was nearly doubled by 15 dpi, indicating a consistent increase. This value peaked at 45 dpi, remained elevated within 60 dpi, then slightly diminished while remaining at a relatively high level. These findings demonstrated that T. gondii infection rapidly triggered IgG antibody production during the acute phase. In addition, the concentration of these antibodies increased when the infection shifted into the chronic phase and further elevated for more than two months. The detection method employed here can identify serum antibody positivity approximately one week into the acute phase of infection, showcasing good specificity and high sensitivity. These results underpin the development of immune serological testing technologies for T. gondii infections.
3.5. Histological and Immunofluorescence Analysis of Mouse Brain
Mice from the CJN and CJI groups were humanely euthanized at 30 dpi, and their brain tissues were harvested for histological examination using H&E staining and immunofluorescence analysis. T. gondii breached the BBB, infiltrated, and proliferated within the brain tissue by 7 dpi. Pathological examination revealed that T. gondii invasion led to significant monocyte infiltration, inducing meningitis (Figure 5A–F). This parasite transitioned from the rapidly replicating tachyzoites to the slowly growing bradyzoites when the infection progressed from the acute to the chronic phase, forming cysts (Figure 5F). Immunofluorescence analysis using recombinant T. gondii proteins effectively detected the presence of cysts within the hippocampal region of mouse brain tissue (Figure 5J–L). This method facilitates qualitative and quantitative assessments of parasite loads in brain tissue and supports research on the three-dimensional distribution of T. gondii cysts within the host brain.
4. Discussion
T. gondii is an important zoonotic parasite that can infect virtually all nucleated cells in warm-blooded animals, primarily transmitted through contaminated food, soil, and direct contact [1]. However, systematic research on the invasion, dissemination, proliferation, and colonization dynamics of orally ingested T. gondii cysts in mice by integrating clinical diagnostics, molecular biology, histopathology, and immunology remains limited [32,33]. Despite measurable humoral immunity in murine Toxoplasma models, therapeutic progress is hindered by unresolved mechanisms regulating tachyzoite–bradyzoite conversion, seroconversion thresholds, IgG neutralization kinetics, and immune memory durability. These critical gaps present challenges in elucidating immune–parasitic oscillatory dynamics that govern chronic, persistent infection [34,35]. Humans are typically infected with T. gondii by consuming undercooked meat contaminated with cysts, particularly pork and lamb [36,37]. Thus, clarifying the transmission pathways of food-borne cysts is essential for developing effective prevention, control, and treatment strategies [38]. Accordingly, we constructed a C57BL/6J mouse model, a strain susceptible to T. gondii, to investigate the spatiotemporal diffusion and colonization of T. gondii cysts following oral infection and the antibody responses (Figure 6).
During acute T. gondii infection, C57BL/6J mice may manifest various clinical signs, including respiratory distress, multi-organ failure, and death [24,39]. Our findings are consistent with previous results showing that T. gondii infection can induce lethal enteritis and abnormal proliferation in intestinal lymph nodes and the spleen [24,40,41]. Oral infection often triggers a “cytokine storm” in the small intestine, driven by Th1-type cytokines. Notably, IFN-γ mediates Paneth cell death by inhibiting mTOR [42], a mechanism resembling that in human inflammatory bowel diseases such as Crohn’s disease. Thus, oral T. gondii infection can cause immunopathology in various animals, in addition to C57BL/6J mice [38]. Chronic T. gondii infection is also associated with increased susceptibility to colitis [40].
Susceptibility to T. gondii varies among mouse strains and is greater in C57BL/6 mice than BALB/c mice [22]. IL-10 plays a key role in reducing mortality and preventing intestinal necrosis in BALB/c and C57BL/6 mice after oral infection [43]. Additionally, oral T. gondii infection has been demonstrated to cause gut microbiota dysbiosis, impair intestinal barrier integrity, and induce colonic inflammation [39,44]. Gram-negative bacteria can exacerbate small intestinal Th1-type immunopathology [45]. However, interventions such as fecal microbiota transplant using probiotics (e.g., Lactobacillus and Bacteroides) have proved to alleviate intestinal inflammation in infected mice [46].
This study showed that T. gondii preferentially targeted the lung tissue during acute infection, thereby disseminating through the bloodstream and causing severe pneumonia. These results align with previous research reporting that a T. gondii pneumonia model exhibited weight loss, ruffled fur, and respiratory rales, complicated by severe lymphocytic infiltration and pulmonary edema [47]. The lung’s role in pulmonary and systemic circulation facilitates efficient T. gondii dissemination, making it a preferred site for infection. This pattern may provide insights into the higher seropositivity rate (60.94%) in lung cancer patients than the general population [48].
This study demonstrated that T. gondii crossed the BBB and colonized the brain approximately 7 dpi. This finding corroborates previous work showing that T. gondii invaded vascular endothelial cells, proliferated, and crossed the BBB into the central nervous system (CNS) [49]. Researchers reported increased inflammatory monocytes in the blood 4 dpi [50]. T. gondii was detected in brain tissues by 7 dpi, indicating its successful passage through the BBB, possibly even earlier than this period [51].
Mice enter the chronic infection stage by 14 dpi. In this case, the animals show partial improvements in locomotor activity, feeding patterns, and body mass indices. However, these parameters remain subnormal compared to uninfected controls throughout persistent infection. T. gondii persists as bradyzoites in low-immunity tissues such as the brain, skeletal muscles, eyes, and heart, leading to lifelong chronic infection [52]. It crosses the BBB through various pathways, including paracellular [53], transcellular [49], and the “Trojan horse” mechanism [50]. The resulting CNS damage can trigger neuropsychiatric symptoms [54]. The reason is that T. gondii induces inflammation in various CNS cells and stimulates the production of inflammatory cytokines and chemokines [55,56].
This study first detected T. gondii-specific IgG antibodies in mouse serum at 8 dpi, with levels gradually increasing and remaining elevated for two months. During chronic infection, its transformation from tachyzoites to bradyzoites facilitated immune detection evasion by forming a highly glycosylated cyst wall [57]. Chronic infection may manifest no overt symptoms under normal immune function; in contrast, declined host immunity can trigger cyst activation, leading to Toxoplasma encephalitis [58,59]. This reactivation is linked to various neurological symptoms, including cognitive impairments [60], suicide [61], and schizophrenia [62]. Researchers demonstrated that chronic T. gondii infection could induce anxiety-like behavior in mice via the “gut–brain axis”, possibly modulated through gut microbiota changes [63].
Initial infection triggers an innate immune response, recruiting monocytes from the bone marrow to the site of infection. These monocytes release cytokines and chemokines, differentiating into macrophages and dendritic cells, contributing to infection control [64]. GTPases induced by IFN-γ, such as IRG and GBP [65,66], are critical for host resistance [32]. Both the current and previous studies highlight that the brain is a primary target for T. gondii infection in mammals, with neurons being the preferred target of colonization and proliferation [67,68,69,70].
5. Conclusions
This research reveals that T. gondii cysts ingested by susceptible C57BL/6J mice are broken down by stomach acid, facilitating the rapid spread of sporozoites to the gastrointestinal tract and spleen. T. gondii enters the bloodstream within 3 dpi, causing fever, weight loss, and even fatal enteritis. T. gondii infiltrates most tissues, crosses the BBB, and colonizes the brain within 7 dpi, inducing inflammation, tissue damage, and an antibody response that persists for months. The lungs are the primary target during acute infection, and the brain is a predilection site in the chronic infection stage. The infection transitions to the chronic phase two weeks post-infection, with the parasite transforming into bradyzoites. These bradyzoites trigger lifelong inflammation and form cysts that persist within the host. These findings provide valuable insights into the underlying mechanisms of T. gondii pathogenesis and host–T. gondii interaction.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Dubey J.P. Toxoplasmosis of Animals and Humans 3rd ed.CRC Press Boca Raton, FL, USA 20222244
- 2Li Z. Sun C.Y. Yang R.D. Xu C.J. Chen M.C. Zhang Y.J. Cong W. Zhu X.Q. Liu Y. Zou F.C. A large-scale serological survey of Toxoplasma gondii infection among persons participated in health screening in Yunnan province, Southwestern China Vector Borne Zoonotic Dis.20191944144510.1089/vbz.2018.233430801237 · doi ↗ · pubmed ↗
- 3Robert-Gangneux F. DardéM. Epidemiology of and diagnostic strategies for toxoplasmosis Clin. Microbiol. Rev.20122526429610.1128/CMR.05013-1122491772 PMC 3346298 · doi ↗ · pubmed ↗
- 4Frenkel J.K. Dubey J.P. Miller N.L. Toxoplasma gondii in cats: Fecal stages identified as coccidian oocysts Science 197016789389610.1126/science.167.3919.8934903651 · doi ↗ · pubmed ↗
- 5Torrey E.F. Yolken R.H. Toxoplasma oocysts as a public health problem Trends Parasitol.20132938038410.1016/j.pt.2013.06.00123849140 · doi ↗ · pubmed ↗
- 6Elsheikha H.M. Marra C.M. Zhu X. Epidemiology, pathophysiology, diagnosis, and management of cerebral toxoplasmosis Clin. Microbiol. Rev.202034 e 00115-1910.1128/CMR.00115-1933239310 PMC 7690944 · doi ↗ · pubmed ↗
- 7Dubey J.P. Lindsay D.S. Speer C.A. Structures of Toxoplasma gondii tachyzoites, bradyzoites, and sporozoites and biology and development of tissue cysts Clin. Microbiol. Rev.19981126729910.1128/CMR.11.2.2679564564 PMC 106833 · doi ↗ · pubmed ↗
- 8Zhou P. Chen N. Zhang R.L. Lin R.Q. Zhu X.Q. Food-borne parasitic zoonoses in China: Perspective for control Trends Parasitol.20082419019610.1016/j.pt.2008.01.00118314393 · doi ↗ · pubmed ↗