Genome-Wide Identification of GLK Family Genes in Phoebe bournei and Their Transcriptional Analysis Under Abiotic Stresses
Yiran Lian, Liang Peng, Xinying Shi, Qiumian Zheng, Dunjin Fan, Zhiyi Feng, Xiaomin Liu, Huanhuan Ma, Shijiang Cao, Weiyin Chang

TL;DR
This study identifies and analyzes GLK genes in Phoebe bournei, revealing their roles in stress responses and potential for genetic improvement.
Contribution
The first genome-wide identification and analysis of GLK genes in Phoebe bournei under abiotic stresses.
Findings
61 PbGLK genes were identified and classified into seven subfamilies.
PbGLK25 and PbGLK30 showed significant upregulation under drought, heat, light, and shade stresses.
Phylogenetic and structural analyses revealed diverse characteristics of PbGLK genes.
Abstract
GOLDEN2-LIKE (GLK) transcription factors are crucial regulators of chloroplast development and stress responses in plants. In this study, we investigated the GLK gene family in Phoebe bournei (Hemsl.) Yen C. Yang, a near-threatened species important for forestry and wood utilization in China. We identified 61 PbGLK genes which were classified into seven subfamilies. Our analyses of their phylogenetic relationships, gene structures, and chromosomal distribution revealed diverse characteristics. Expression profiling under different tissues and abiotic stresses showed that PbGLK25 and PbGLK30 were particularly responsive to drought, heat, light, and shade stresses, with significant upregulation. These findings highlight the potential role of PbGLK genes in stress adaptation and provide insights for the genetic improvement of P. bournei.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
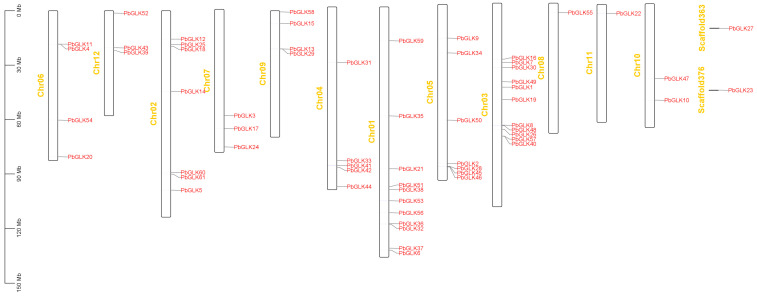 Figure 1
Figure 1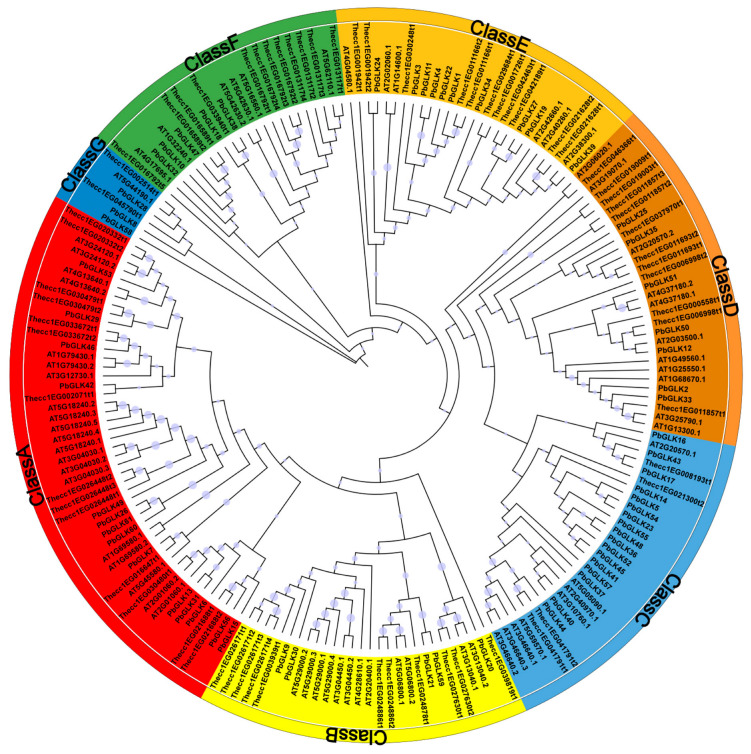 Figure 2
Figure 2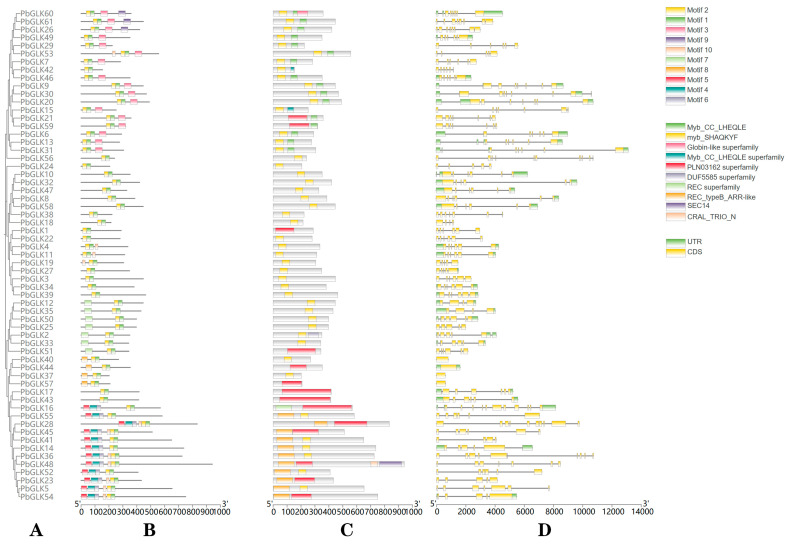 Figure 3
Figure 3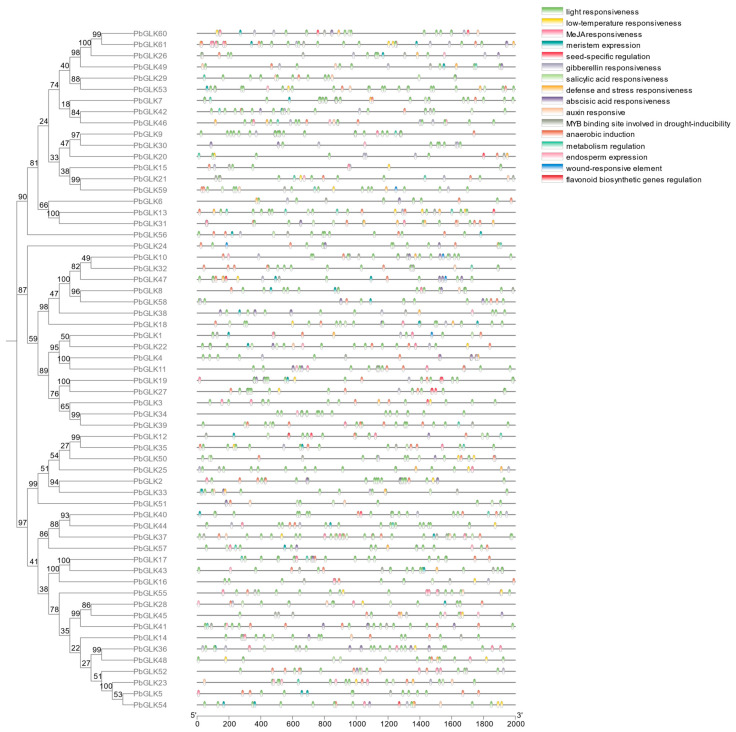 Figure 4
Figure 4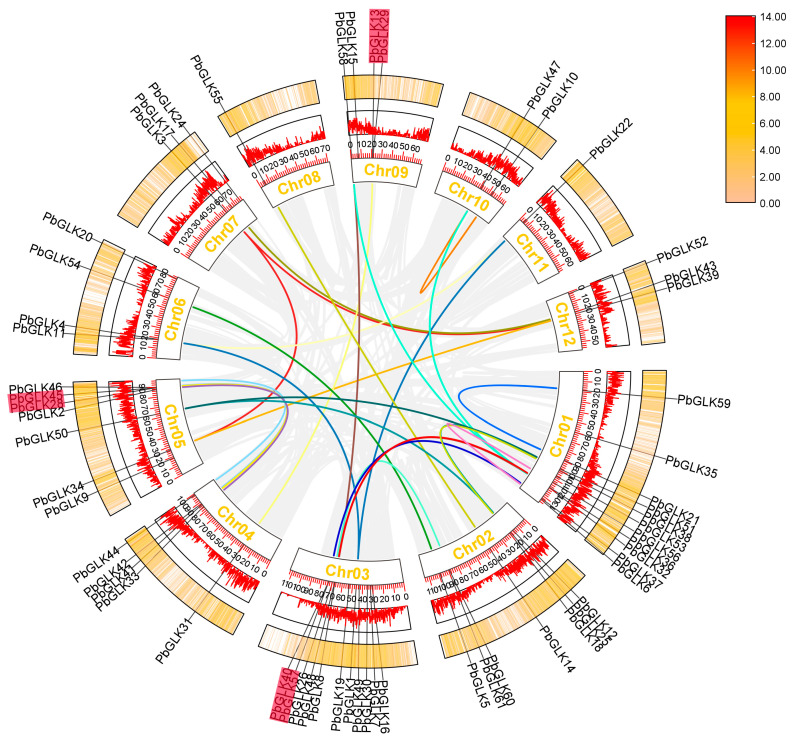 Figure 5
Figure 5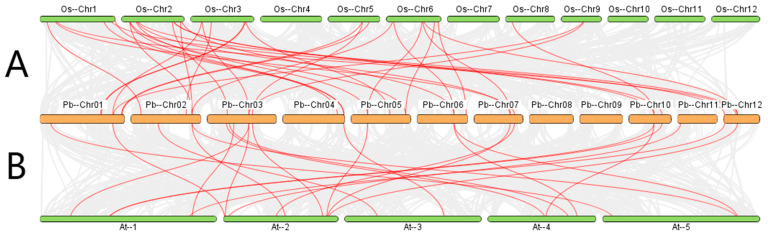 Figure 6
Figure 6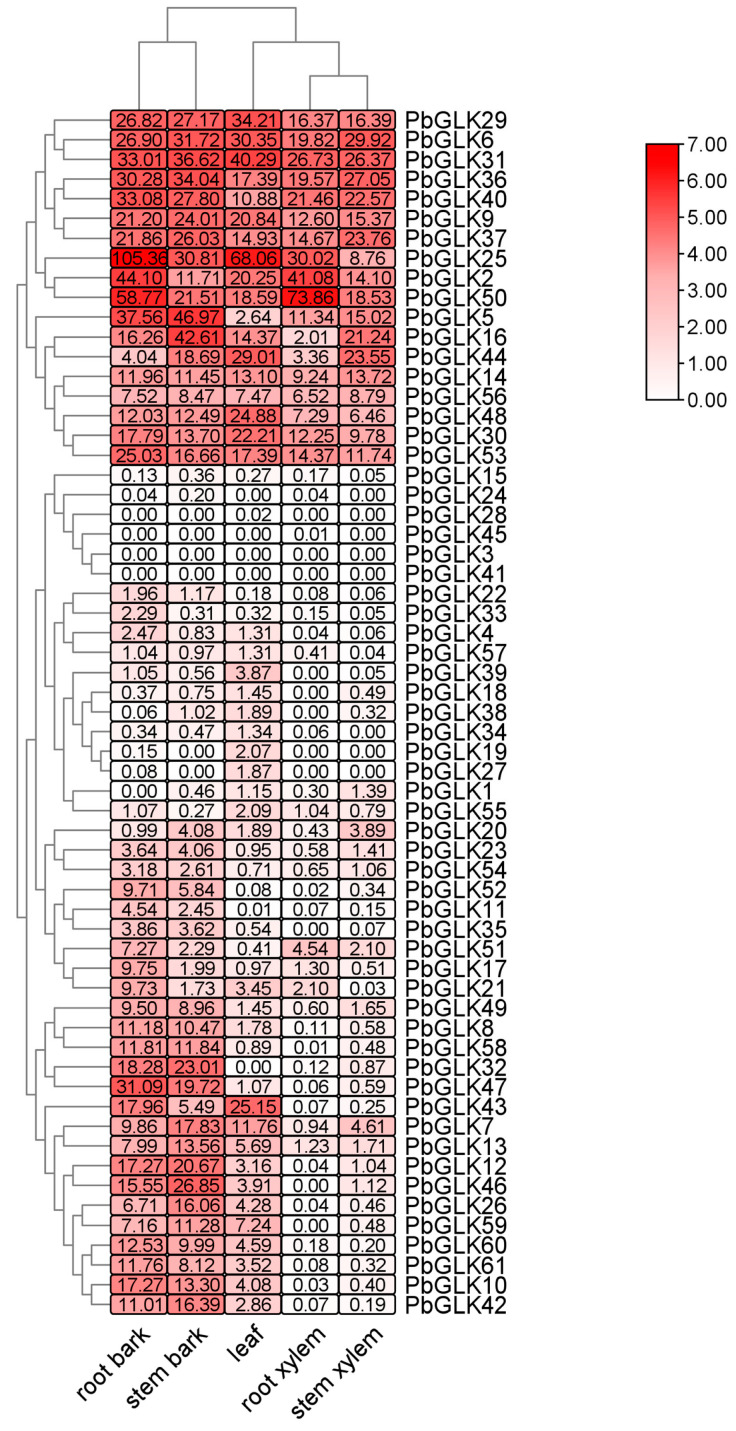 Figure 7
Figure 7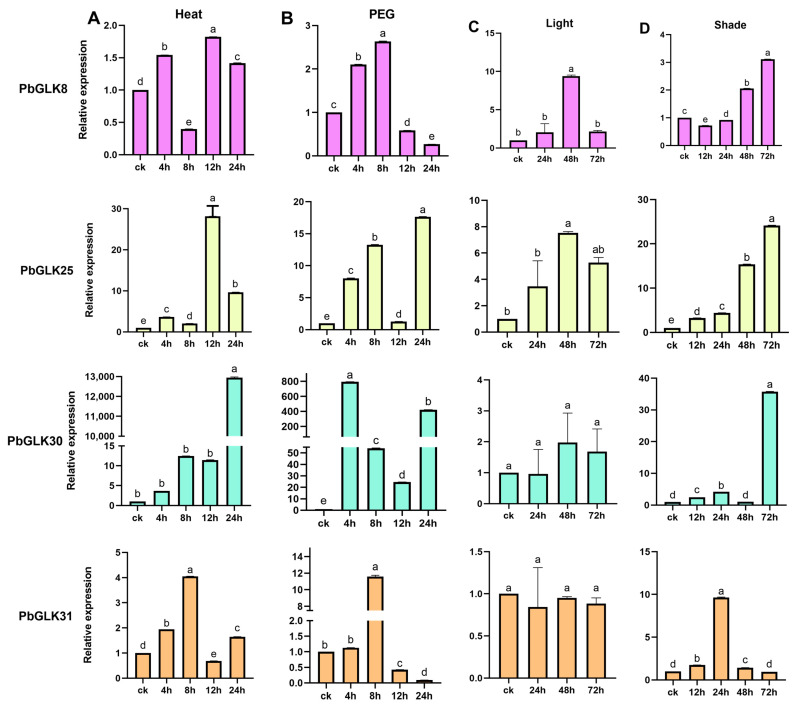 Figure 8
Figure 8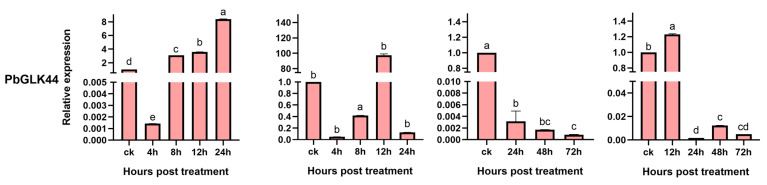 Figure 9
Figure 9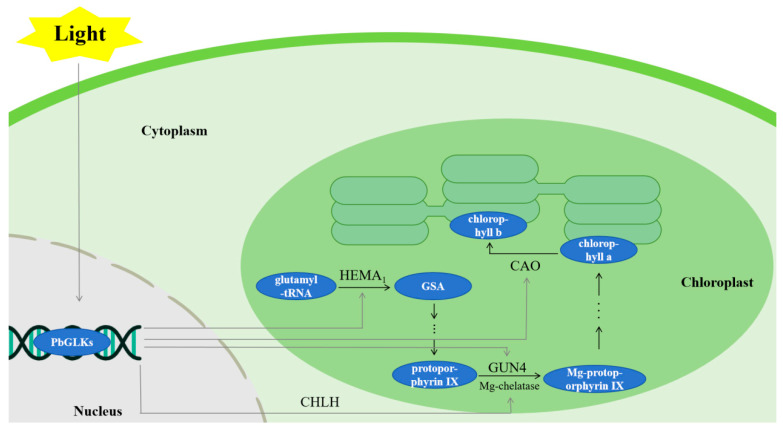 Figure 10
Figure 10- —Fujian Agriculture and Forestry University Forestry Peak Discipline Construction project
- —Fujian Agriculture and Forestry University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhotosynthetic Processes and Mechanisms · Plant Gene Expression Analysis · Plant Molecular Biology Research
1. Introduction
Transcription factors (TFs), also known as trans-acting factors, are proteins that bind to cis-acting elements within the promoter regions of target genes to regulate their expression through activation or inhibition [1,2]. These proteins control gene expression by attaching to specific cis-regulatory sequences within the promoters of target genes [3]. In the plant genome, approximately seven percent of coding sequences belong to TFs [4], underscoring their prevalence and importance in genomic regulation. TFs are a highly diverse group of regulatory proteins that play critical roles in various biological processes. For instance, they regulate plant responses to abiotic stresses such as saline–alkaline environments, temperature changes, drought, UV radiation, and other challenging conditions [5,6,7]. By modulating gene expression, TFs intricately control cellular processes and enable plants to respond effectively to external stimuli [8]. Additionally, TFs facilitate the optimization of the trade-off between stress tolerance and growth in plants, thereby enhancing their resilience and adaptability to adverse environmental conditions [9,10]. Therefore, understanding the factors and mechanisms that influence the functionality of plant TFs is crucial for deciphering the complex regulatory networks governing plant development, growth, and environmental adaptation [1,11].
Throughout a plant’s lifetime, it frequently encounters unfavorable or stressful conditions, including abiotic stresses such as drought, cold, salinity, and nutrient deficiency, as well as biotic stresses such as pathogen infections and herbivore attacks [12]. The GARP (Golden2, ARR-B, and Psr1) superfamily comprises a group of transcription factors that play essential roles in various biological processes, including hormonal signaling, nutrient sensing, chloroplast biogenesis, plant development, circadian clock oscillation, and floral transition [13,14]. Among these, GLK (Golden2-Like) proteins are key members of the GARP superfamily [13,15] and typically contain two conserved domains: a DNA-binding Myb-related domain and a dimerizing GCT domain [16]. Numerous studies have investigated their functions in plant growth, development, and resistance to abiotic stresses. The GLK gene in cotton has been shown to be significantly responsive to drought, salt, temperature stress, and developmental processes. Overexpression of the GLK gene in cotton significantly improves its resistance to these stresses, resulting in higher survival and yield under adverse conditions. These genes play a crucial role in regulating plant growth and stress tolerance by regulating cell responses to environmental stress sources [17]. In Glycine max (L.) Merr., research on GLK genes has revealed their critical roles in crop development and metal ion stress [18]. Additionally, studies on UV-B stress have identified that GLK transcription factors are highly conserved during evolution and may be associated with abiotic stress responses [19]. To cope with and resist these adverse conditions, plants have evolved multiple specific regulatory networks. For example, studies have identified candidate upstream regulators that enhance GLK1’s adaptability to varying environmental conditions by modulating its baseline activity, thereby revealing changes in the activity of the GLK1-centered regulatory network [20]. This highlights the critical role of GLK1 within these networks, as it belongs to the GLK transcription factor family, a significant group of transcription factors in plants that orchestrates responses to environmental stressors.
The GLK transcription factor family represents a significant group of transcription factors in plants [21]. Notably, research indicates that certain components of the GLK regulatory network have independently evolved in diverse species [22]. Some studies have found that while the function of GLKs is conserved across species, their binding sites are predominantly species-specific, with conserved binding sites typically located near photosynthetic genes regulated by GLK expression and whose expression is highly sensitive to GLK mutations [23,24]. This suggests a close association between GLKs and photosynthesis. Additionally, GLK transcription factors have been studied in various plants, including Arabidopsis thaliana (L.) Heynh, Solanum lycopersicum L. (tomato), Oryza sativa L. (rice), Zea mays L. (maize), Hordeum vulgare L. (barley), Phyllostachys edulis (Carrière) J. Houzeau (Moso Bamboo), and Nicotiana tabacum L. [25,26,27,28,29,30,31]. Many studies have highlighted the critical role of GLKs in chloroplast development. For example, in A. thaliana, GLKs directly bind to the promoter regions of PhANGs, thereby promoting chlorophyll biosynthesis, the assembly of the photosynthetic apparatus, and, subsequently, chloroplast development [32]. In tomatoes, BPG4 regulates chloroplast development and homeostasis by repressing GLK transcription factors and is involved in light and brassinosteroid signaling [26].
Few transcription factors have been identified to positively regulate chloroplast biogenesis, with the GLK gene family being an exception. Research indicates that members of the GLK family are essential for the formation and development of chloroplasts, thereby impacting plant photosynthesis and fruit quality [33,34,35]. It has been reported that GLK family transcription factors play a critical role in regulating chloroplast development and chlorophyll accumulation in the monocot maize and the eudicot A. thaliana, as well as in other plants, to adapt to varying environmental and developmental conditions [36,37,38]. Studies have shown that GLK genes assist in the coordinated regulation and synchronization of a certain set of nuclear photosynthesis genes, thereby optimizing photosynthetic capacity under various environmental and developmental conditions [24,39]. This enhanced photosynthetic efficiency subsequently promotes plant growth and development. In this study, an enrichment analysis of the GLK gene found that protein–chromophore linkage, photosynthesis, light harvesting, and the chlorophyll biosynthetic process were the most enriched biological processes. These results suggest that the GLK gene promotes chlorophyll biosynthesis and the expression of nuclear genes involved in the assembly of Lhcb (an important component of photosystem I (PSI) and photosystem II (PSII)) [39]. Although the important role of GLK genes in photosynthesis is well recognized, the vast majority of its biological functions remain unexplored and need further study.
In China, P. bournei is not only utilized as a preservative wood in wood art, shipbuilding, and architectural projects but is also an important tree species for use in artificial forestation [40,41]. However, its extensive use and afforestation effects have led to a decrease in its population [42]. Therefore, P. bournei has been classified as a near-threatened species, and continuous conservation efforts are imperative to protect it. It is particularly important to investigate the effects of various factors affecting the growth and development of P. bournei, hence restoring the amount of P. bournei in wild forests [43]. It has been suggested that the rapid growth of P. bournei under shaded conditions might be related to the increased expression of genes like GLK, which allows the plant to maintain high photosynthetic rates even in low-light environments [44]. However, the specific relationship between GLK genes and the growth of P. bournei remains to be further explored. In this study, we conducted a comprehensive study of the GLK gene in P. bournei, including genome-wide identification, phylogenetic analysis, gene structure, chromosome location, replication events, cis-acting element analysis, and expression levels under different tissues and stresses. It is anticipated that the findings from this study will not only enhance our current understanding of GLK genes in P. bournei and broader woody plant species but also provide a set of gene tools for their genetic improvement.
2. Results
2.1. Identification of PbGLK Genes in P. bournei
We identified a total of 61 GLK genes in P. bournei and renamed PbGLK01 to PbGLK61 (Table 1). The 22 GLK proteins encode amino acids ranging from 153aa to 941aa, with an average length of 404aa. The size of a protein is usually proportional to the length of its amino acid sequence. In the present study, the associated molecular weights of the 61 PbGLK proteins ranged from 17.56 kDa (PbGLK42) to 103.96 kDa (PbGLK48), with an average of 44.823 kDa, indicating a significant difference in the size of the GLK transcription factor proteins. A total of 34 PbGLK proteins were acidic (pI < 7.0), and the remaining 27 were basic, with a total value between 5.10 and 10.21. Meanwhile, six proteins (PbGLK7, PbGLK18, PbGLK23, PbGLK29, PbGLK42, and PbGLK52) were stable proteins, and the rest were unstable proteins (Instability Index > 40), ranging from 41.06 to 78.03. The data show that the aliphatic index ranged from 54.37 to 88.87, with a mean value of 70.97, reflecting the high thermostability of most PbGLK proteins. In addition, the mean value of the hydrophilicity of the PbGLK proteins was negative, which meant that all the PbGLK proteins were hydrophilic. According to the prediction of subcellular localization, there are 56 PbGLK proteins localized in the nucleus, followed by two each in the chloroplast and cytosol, and PbGLK1 in the mitochondrion.
2.2. Phylogenetic Analysis and PbGLK’s Chromosomal Locations
In this study, we determined that a total of 59 PbGLKs were distributed on 12 chromosomes (Chr), with PbGLK23 and PbGLK27 located in Scaffold376 and Scaffold363, respectively (Figure 1). We found that the distribution of GLK genes across the chromosomes was uneven. Most PbGLK genes were found in chromosomes 1 and 3, with both having 11 PbGLKs, whereas chromosomes 8 and 11 had the least PbGLK genes and carried only one GLK gene, named PbGLK55. Chr02, Chr03, Chr04, and Chr05 each had 3–5 PbGLK genes located in very close proximity.
To investigate the evolutionary characteristics and relationships among the GLK proteins, we constructed a phylogenetic tree (Figure 2) using 61 GLK proteins from P. bournei alongside 64 and 66 GLK proteins from Arabidopsis and cocoa (Theobroma cacao L.), respectively. A total of 191 GLK proteins were divided into seven subfamilies (A-G) (Figure 2). The phylogenetic analysis showed that 48 GLK family members were concentrated in subfamily A, while subfamily G had only seven. Among them, the subfamily with the most PbGLK family members was subfamily C, with 17 members, accounting for 27.87% of the total. The second was subfamily A, with 14. However, subfamily G had only three PbGLKs (PbGLK8, PbGLK28, and PbGLK58). In contrast, among the AtGLK family members, subfamily A had the most with 20, and subfamily G had the least with only one. In the TcGLK family, subfamily A had up to 14 members, whereas subfamily G had a minimum of one member. These results indicated that the number of GLK genes in each subfamily differed among species.
2.3. PbGLK Gene Structure, Conserved Motif, and Domain Analysis
We analyzed the exon/intron structure of 61 PbGLK genes and found that the exon number of GLK genes ranged from one (PbGLK37, PbGLK40, PbGLK44, and PbGLK57) to 11 (PbGLK16) (Figure 3D). Subfamily G had the highest average exons (seven) per gene, and subfamily C had the lowest (five). There are also differences in gene structure between different subfamilies. For example, the genes in subfamily B have seven exons, while those in subfamily D have four to six exons.
Ten conserved motifs (Motif 1–10) of 61 PbGLK proteins were analyzed. Similar conserved motif compositions are often present in most GLK proteins from the same subfamily. Motif 1 and Motif 2 are present in all 61 PbGLK proteins, while several motifs appear only in specific subfamilies. For instance, only three out of 61 PbGLK proteins contain Motif 9, and all belong to subfamily A. All proteins of subfamily D contain Motif 7, which is present only in this subfamily. All members of subfamily B have Motif 3, and two (PbGLK42, PbGLK56) of the 14 genes of subfamily A have no Motif 3. In addition, the myb_SHAQKYF conserved domain can be identified in almost every PbGLK gene. It can be inferred from the analysis that the Myb_CC_LHEQLE domain is identified in both subfamilies A and B, and the expression levels of these two subfamilies are relatively high in different plant tissues (see figure in Section 2.7). Thus the biological role of proteins with the Myb_CC_LHEQLE domain can be further analyzed.
Therefore, the classification of PbGLK subfamilies was further supported by the differences in gene structure, conserved motif arrangement, and phylogenetic trees between the same subfamily and different subfamilies mentioned above. The diversity of the number, arrangement, and distribution of different motifs in the different subfamilies may be what differentiates them from one another.
2.4. Cis-Acting Elements: Analysis of the PbGLK Gene Family
We identified cis-acting elements as DNA sequences located in the upstream promoter region of the genes, which can bind transcription factors to regulate gene function [45]. For example, it has been found that the HD-Zip transcription factor ArHDZ22 may regulate growth-related genes through its interaction with the hdzb cis-acting elements [46]. Additionally, experimental studies have identified a 150-bp cis-acting element within the promoter region of the AtNRT2.1 gene, which plays a role in regulating gene expression in response to nitrogen and carbon states in plants [47]. Beyond natural promoter regions, transposable elements (TEs) can also influence gene expression by introducing new cis-acting regulatory elements into unconnected genomic locations [48]. Collectively, these findings highlight the critical role of cis-acting elements in determining the level and specificity of gene expression, a concept that has gained increasing recognition in recent years [49]. By detecting the 2000 bp promoter sequence of the upstream PbGLK genes, it was found that there were 16 cis-acting elements in the promoter region of the PbGLK gene family, involving four growth and development response elements, seven stress response elements, and five hormone response elements (Figure 4). Among them, the largest number of functional elements was light responsiveness, with 688 (48.18%). Cis-elements related to light-response functions are widely distributed in the PbGLK gene promoter, suggesting that the GLK gene may play a role in plant light responses. In addition to the most light-responsive components, a total of 500 components related to hormone response were found, including responses to various hormones such as auxin, gibberellin (GA), salicylic acid (SA), and methyl jasmonate (MeJA), indicating that the PbGLK gene holds significant importance in hormone response. Relatively few components include but are not limited to wound-responsive elements (5), seed-specific regulation (13), and endosperm expression (21). While the high abundance of light-responsive elements suggests that PbGLK family genes may be particularly sensitive to light, the presence of these elements alone does not confirm a definitive role in light responses. Additionally, the significant number of hormone-responsive elements indicates that PbGLK genes may also play important roles in hormone signaling pathways. Further functional studies are required to elucidate the specific roles of PbGLK genes in these processes.
2.5. Intraspecific Collinearity Analysis of PbGLK Genes
Gene duplication events, including tandem repeat events and segmental duplication, play an important role in gene amplification and the generation of new functions, which are conducive to the development of species [50]. To better explore the evolutionary conservation of GLK genes in P. bournei, we examined genome-wide duplication events of PbGLK members in P. bournei. The results show a pattern of collinearity between duplicated gene pairs throughout the P. bournei genome (Figure 5). We found that there were 26 pairs of segmental duplications in the P. bournei genome, and the segment duplication events are mainly concentrated between Chr01 and Chr03 and between Chr04 and Chr05, which suggests that these genes may have arisen through gene duplication events, and that duplication events are the main drivers of the evolution of new functions for PbGLK genes.
2.6. Synteny Analysis Among PbGLK Genes
To explore the possible evolutionary patterns of the GLK gene family of P. bournei in different plant varieties, we performed synteny analyses of the GLK genes of P. bournei with GLK genes of two other typical plants (Figure 6), including one typical dicotyledonous plant, A. thaliana, and one representative monocotyledonous plant, Oryza sativa. Our analysis shows that 22 PbGLK genes were collinear with Oryza sativa, and 19 PbGLK genes were collinear with A. thaliana. There were 34 pairs of P. bournei genes collinear with rice and 23 pairs of P. bournei genes collinear with A. thaliana. Although our synteny analysis revealed that more PbGLK genes were collinear with Oryza sativa than with A. thaliana, the level of collinearity does not directly equate to phylogenetic relationships or gene evolutionary distance. The observed patterns may be influenced by multiple factors, including genome rearrangement, gene loss, and duplication events [51]. Therefore, while these results suggest a higher degree of conservation between PbGLK genes and Oryza sativa genes compared to those in A. thaliana, further studies are needed to elucidate the precise evolutionary relationships among these genes.
The results showed that the GLK gene of Oryza sativa was genetically closer to the GLK gene of P. bournei than that of A. thaliana.
The proportion of P. bournei and other species containing similar GLK genes suggests, to some extent, the evolutionary relationship of GLK genes among P. bournei, monocots, and dicots.
2.7. Expression Analysis of PbGLKs in P. bournei Tissues
We compared the expression levels of 61 PbGLKs across five tissues of P. bournei: root bark, stem bark, root xylem, stem xylem, and leaf (Figure 7). Apart from a few specific genes, most genes were generally expressed across the five selected plant tissues. There are only three genes in subfamily G, namely, PbGLK8, PbGLK28, and PbGLK58, among which PbGLK8 and PbGLK58 are expressed in all tissues and have the highest expression levels in root bark and stem bark, while PbGLK28 is only slightly expressed in leaves. In subfamily F, PbGLK32 was not expressed in leaves, PbGLK18 and PbGLK38 were not expressed in root xylem, and the other two genes were expressed in all tissues and were highly expressed in the epidermis. The average gene expression of subfamily E was the lowest among the seven subfamilies. In contrast to subfamily E, subfamily D had the highest average gene expression. PbGLK25 exhibited extremely high gene expression in the root bark and leaves, while PbGLK50 showed extremely high gene expression in the root bark and xylem. It is worth noting that the expression of PbGLK25 was the highest among 61 PbGLKs. Most genes in subfamilies A, B, and C were expressed in all tissues, and their expression levels were relatively high. These findings suggest that the expression levels of PbGLK vary within and between the subfamilies.
Overall, the expression levels were lowest in the xylem, whereas the epidermis showed higher expression compared to both the xylem and leaves, with the root epidermis exhibiting the highest expression levels. This may be due to the fact that PbGLK genes are largely involved in epidermal growth and development.
2.8. Expression of PbGLK Genes Under Abiotic Stress
Due to their immobility, plants are exposed to a more variable environment compared to other organisms, with abiotic stresses such as drought, heat, light, and shade stress significantly impacting their growth and development [52,53,54]. In order to study the role of PbGLK genes in plant responses to these stress, five PbGLK genes, PbGLK8, PbGLK25, PbGLK30, PbGLK31, and PbGLK44, were selected from seven subfamilies (A–G) for real-time quantitative polymerase chain reaction (qRT-PCR) based on the subfamily they belong to (less than two from one subfamily) (Figure 2), the number of cis-acting elements in each gene (Figure 4), and their expression levels in various tissues of P. bournei (Figure 8).
In response to elevated temperatures, the expression levels of various genes exhibited changes across distinct time intervals. Notably, following exposure to high temperatures, the expression levels of the genes PbGLK25 at 12 h and PbGLK44 at 24 h were observed to rise by approximately 28-fold and 8-fold, respectively, relative to their pre-treatment levels. Additionally, the expression of the PbGLK30 gene was markedly elevated at the 24 h mark, leading us to speculate that these genes are associated with plant adaptation to heat stress. In arid environments, except PbGLK25 and PbGLK30, the expression of most genes gradually decreases after 12 h. The expression level of PbGLK30 reached its peak after 4 h, and the maximum expression level of PbGLK30 was about 793 times that of the control group, indicating a potentially crucial role of the PbGLK30 gene in the plant’s response to drought conditions. When the genes were exposed to light stress, most of the genes showed a trend of low–high–low change. Compared to the control group, the expression levels of the genes PbGLK44 and PbGLK31 were reduced, with PbGLK44 showing the most pronounced suppression under light stress conditions. We therefore hypothesized that the PbGLK44 gene may not be beneficial for plants in effectively managing light stress. When examining the impact of shade stress on gene expression, it becomes evident that the expression of PbGLK44 and PbGLK31 diminished most notably over time. Conversely, the expression of the other three genes continued to rise even until the 72 h mark. Among them, PbGLK25 and PbGLK30 exhibited the highest levels of expression, increasing 24-fold and 36-fold, respectively, compared to the control group. These findings suggest that PbGLK25 and PbGLK30 may play important roles in the plant response to shade stress.
To summarize, the expression levels of almost all genes reached their peaks during specific intervals under drought, heat, light, and shade stress conditions. The expression of the PbGLK genes increased across most treatment periods compared to the beginning, suggesting that PbGLK could be vital for plants in managing these four types of stress. Notably, PbGLK25 and PbGLK30 displayed pronounced responses to drought, heat, light, and shade stress, indicating that P. bournei may be primarily regulated by some specific PbGLK genes when facing these stresses. While these findings provide insights into the potential functions of PbGLK genes under different stress conditions, further experiments are necessary to validate their exact roles and functional significance.
3. Discussion
GLK transcription factors have been demonstrated to be crucial for chloroplast development [23,25,55], photosynthesis [27,56], stress resistance [30,57], and hormone responses [22,30]. Due to the importance of the GLK gene family in abiotic stress and plant growth and development, it has been studied in plants such as A. thaliana [25] and maize [58]. To date, the whole-genome analysis of GLK has been conducted in plants such as Moso Bamboo, Gossypium hirsutum L., and Camellia sinensis (L.) Kuntze (Camellia) [12,30,45], but a whole-genome analysis of P. bournei has still not been carried out. Therefore, the genome-wide characterization and expression analysis of the GLK gene family in P. bournei will enhance our understanding of GLK genes within the same plant and the GLK gene family across different species, providing deeper insights into the functions and roles of these genes.
In this study, we used bioinformatics to identify the PbGLK gene family and discovered 61 PbGLK genes separated into seven subfamilies (Figure 2). The GLK genes in Camellia and Eucalyptus grandis W. Hill ex Maiden were also divided into seven subfamilies for analysis [22,59]. In comparison, the number of GLK genes in the G/I subfamily of tea tree and P. bournei was the lowest, while those distributed in the A/VII subfamily of Eucalyptus grandis and P. bournei were relatively high, accounting for the second highest and highest, respectively, in their respective comparisons. In addition, we found that P. bournei had three and 13 fewer GLK members than A. thaliana and cocoa, respectively (Table S1). The GLK family members from the three groups were intermingled, with no distinct branches observed (Figure 2). These results suggest that the GLK gene family shows a similar evolutionary pattern in P. bournei, Camellia, and Eucalyptus grandis, with members distributed in multiple subfamilies. Additionally, the GLK gene family in dicots exhibits limited evolutionary divergence and remains highly conserved across species. However, the differential expansion or contraction of specific subfamilies (e.g., A/VII and G/I) among species may reflect lineage-specific adaptations to varying ecological niches and environmental pressures. For instance, the expansion of the A/VII subfamily in P. bournei and Eucalyptus grandis could be linked to their adaptation to woody growth habits and stress tolerance mechanisms, as these genes may play roles in regulating light-responsive pathways and enhancing photosynthetic efficiency under adverse conditions. Conversely, the reduced size of the G/I subfamily in P. bournei and tea tree might indicate functional redundancy or specialization within this group, potentially driven by niche-specific selective pressures. Although our data suggest that the GLK gene family is conserved to some extent in these species, to fully understand the evolutionary dynamics and conservation of the GLK gene family in dicotyledonous plants, further comprehensive analyses in a wider range of dicotyledonous species are needed in subsequent studies.
GLK often works synergistically with other genes or with proteins and substances that are the products of their expression. For example, the interaction between GLK and TCP15 may help coordinate the expression of cell expansion genes and genes involved in the development of photosynthetic organs [55]; transgenic GLK facilitates tumor metastasis and cellular migration by leveraging the IQ motif within the scaffold protein that contains GTPase-activating protein 1 (IQGAP1) [60]; and BPG4 interacts with GLK, affecting the amount of chlorophyll and the size of the light collection complex [32]. In addition, AcGLK1 and AtGLK transcription factors are also thought to affect chloroplast development or chlorophyll accumulation [61,62,63]. Additionally, GLK is involved in modulating the various activities of chlorophyll synthases, including δ-aminoacetobulinate dehydratase (ALAD), porphyrinogen deaminase (PGBD), and protoporphyrinogen oxidase (PROTOX) [64]. These studies suggest that GLK often acts in concert with other factors and plays an important role in chloroplast development and accumulation. It is worth noting that our analysis of the subcellular localization prediction of PbGLKs showed that most PbGLKs were located in the nucleus. Only PbGLK6 and PbGLK21 were located in chloroplasts (Table 1), a finding similar to the results of a whole-genome analysis of GLK in Eucalyptus grandis, Citrus Cultivar Kanpei, and so on [12,65], again suggesting that the evolution of the GLK gene family is conservative. In addition, PbGLK21 was highly expressed in the root epidermis, root xylem, stem epidermis, stem xylem, and leaves (Figure 7). We speculate that the function and action of this gene are related to the physiological activity of chloroplasts, which needs to be verified by further experiments.
As has been shown, gene duplication events are not only an important mechanism for gene diversification but also provide opportunities for the acquisition of new gene functions [62]. In this study, we analyzed variants in conserved motifs in the GLK protein family and discussed the role of these variants in the diversification of gene functions. The myb_SHAQKYF transcriptional repressor, found in A. thaliana, can regulate leaf wax biosynthesis through the transcriptional repression of DEWAX [64]. In our analysis, myb_SHAQKYF was identified as a key conserved structural domain of the PbGLK gene family in almost all PbGLK genes (Figure 3C). It is suggested that this structural domain may have an important regulatory role in gene function and that the GLK genes may be regulating non-stomatal transpiration and, thus, responding to stress by regulating leaf wax.
Gene duplication and conserved motif variation are bridges connecting different subfamilies [66]. In P. bournei, all PbGLKs contain Motif 1, which varies between subfamilies but is conserved in each subfamily. This conservatism may reflect the importance of specific genes in specific biological functions [45]. However, Motifs 17 and 18 were only observed in subfamilies C and D, suggesting that these specific motifs may be associated with subfamily-specific functions. In Moso Bamboo, Motif 5 is present in subfamilies 1, 2, 3, 4, 5, 6, and 7, while Motifs 5, 6, and 8 occur in subfamily 10. It can be inferred that the amino acid variation in Motif 5 may be an important cause of its phylogenetic differentiation [30]. In the conserved motif analysis of this experiment, we found that all members of subfamily D contain Motif 7, while none of the genes of subfamily C contain this motif, suggesting that the loss of Motif 7 may be related to the evolutionary division of the subfamily (Figure 3B). All members of subfamily B and some members of subfamily A may have arisen from tandem duplication gene events, whereas PbGLK42 and PbGLK56 do not contain Motif 3. This loss of Motif 3 may have led to the divergence of the PbGLK gene family through tandem gene duplication events. In this context, the different evolutionary pathways of PbGLK42 and PbGLK56 may represent the starting point for functional diversification. Duplication events not only promote the expansion of gene family size but may also trigger the loss or gain of specific motifs, which, in turn, leads to the diversification of gene functions.
Abiotic stresses such as drought, heat, light, and shade significantly impact plant growth and development [67], and our findings indicate that GLK genes may play crucial roles in plant responses to these stresses by integrating developmental and stress-responsive pathways. Chloroplasts are essential for photosynthesis and act as environmental sensors [68], but their function can be compromised under stress conditions [69]. GLK genes, which regulate chloroplast development, might help maintain chloroplast integrity during stress; for instance, the substantial upregulation of PbGLK30 under drought stress (793-fold) suggests its potential role in protecting chloroplast function and optimizing photosynthetic efficiency when water is scarce. Furthermore, the abundance of light-responsive cis-acting elements in the promoters of GLK genes highlights their involvement in balancing light signaling and stress adaptation. Under light stress, the suppression of PbGLK44 indicates a possible negative regulatory role in preventing excessive photosynthetic activity that could lead to oxidative damage, while the upregulation of PbGLK25 and PbGLK30 under shade stress reflects their importance in compensating for reduced light availability. Additionally, the presence of numerous hormone-responsive elements in the PbGLK gene promoters implies that these genes participate in hormone-mediated stress responses, potentially interacting with pathways involving auxin, gibberellin, salicylic acid, and methyl jasmonate. The distinct expression patterns of PbGLK genes across different stress conditions further demonstrate their functional diversity, with PbGLK25 and PbGLK30 showing pronounced responses to multiple stresses, suggesting they are key regulators in plant stress adaptation. While these insights provide a foundation for understanding the roles of GLK genes in abiotic stress responses, further functional studies are necessary to clarify the underlying mechanisms and validate their significance in plant resilience.
Research has elucidated the impact of light and hormonal signals on chloroplast development and chlorophyll biosynthesis within tea tree (Camellia sinensis) [70]. The direct targeting of a common promoter cis-box (G-box) can optimize photosynthetic performance and growth through a series of processes [71], so we speculate that gene response to stress and the presence of cis-elements in plants jointly affect plant photosynthetic performance. In the above stress analysis, some GLK genes showed significant responses to light and shade, suggesting that these genes may be involved in responses to abiotic stresses (Figure 8). In addition, the above promoter analysis showed that, although cis-elements related to hormone, stress, and developmental regulation are also found in GLK promoters, those that respond to light are the most abundant. Under light conditions, the expression of PbGLK is closely related to four key enzymes of chlorophyll biosynthesis [72,73,74]. Based on the above findings, the role of PbGLK genes on chlorophyll synthesis under light conditions can be predicted (Figure 9).
Although this study provides some insights into the evolutionary patterns and functions of the GLK gene family in P. bournei, some limitations should be acknowledged. First, we did not perform subcellular localization experiments for the PbGLK proteins. This decision was based on the high conservation of GLK genes across species and consistent reports in the existing literature that indicate their nuclear localization. Given this strong evidence, we believe the predictions of subcellular localization are reliable. Second, while our data suggest a certain degree of conservation of the GLK gene family in these species (P. bournei, Camellia, and Eucalyptus grandis), a more comprehensive analysis of a broader range of dicotyledonous plants is required to fully understand the evolutionary dynamics and conservation of the GLK gene family in dicots. Additionally, although the presence of high-abundance, light-responsive, and hormone-responsive cis-acting elements implies potential roles of PbGLK genes in light- and hormone-signaling pathways, their exact functions remain unclear and require further experimental validation. The current study primarily relies on bioinformatics analyses, such as sequence comparisons and cis-element predictions, but lacks direct functional evidence from experiments like gene knockout, overexpression, or promoter activity assays. Finally, the roles of PbGLK genes in biological processes, including light and hormone signaling, need to be experimentally validated through functional studies to confirm their contributions to these pathways.
4. Materials and Methods
4.1. Genome Data and Plant Material Source
The genome sequence data and annotation information of P. bournei were downloaded from the Sequence Archive of China National GeneBank Database (CNSA) with accession number CNP0002030 [70]. Genome sequence files of A. thaliana and O. sativa were acquired from EnsemblPlents and Phytozome v13, respectively. The RNA-seq data from different tissues of P. bournei were downloaded from BioProject with accession number PRJNA628065. The plant materials were derived from one-year-old P. bournei seedlings cultured in an artificial climate box under different treatments. P. bournei seedlings with the same growth potential for one year were selected for treatment, and the materials were divided into the control group and stress treatment group, with 30 strains in the treatment group and three strains in the control group. Every two strains in the treatment group were used as a biological replicate, and three groups of biological replicates were set in each time period. During the stress treatment period, the seedlings were cultured in an artificial climate incubator with a temperature of 25 °C and a humidity of 75%. Then, the P. bournei seedlings were exposed to drought stress (10% PEG6000) and heat stress (40 °C). Samples of mature leaves were collected at 0 (CK), 4, 8, 12, and 24 h. Mature leaf samples under light stress (light 24 h/d) and shade stress (light 0 h/d) were picked at 0 (CK), 24, 48, and 72 h and 0 (CK), 12, 24, 48, and 72 h, respectively. The control group (CK) was treated with distilled water and normal growth conditions. After various treatments, P. bournei leaf samples were collected and stored in liquid nitrogen at −80 °C for RNA extraction.
4.2. Identification and Physical and Chemical Property Analysis
The conserved domain of the A. thaliana GLK gene family was downloaded from PlantTFDB. A local BLASTp search was used to compare the conserved domain between P. bournei and A. thaliana to screen the candidate GLK genes in P. bournei [71]. The repetitive results of the BLASTp search were removed. The identified protein sequences of GLK genes from P. bournei were submitted to the NCBI for BLASTp to perform further searches. To further identify GLK gene family members, the GLK conserved domain HMM model (PF00249) was downloaded from the Pfam database using HMMER-3.2.1 with an e-value <10^−5^ and other parameters by default. After the identification of GLK genes in P. bournei, the online website ExPASy was used to analyze the physical and chemical properties of the GLK proteins that were identified [72].
4.3. Chromosomal Distribution and Gene Duplication of PbGLK Genes
TBtools was used for grepping the location information of the PbGLK genes from the genome (FASTA) file and the annotation (GFF) file of P. bournei [72]. The syntenic relationships of PbGLK were determined using MCScanX (version 1.00) with default parameters and plotted using TBtools-II v2.105 [73].
4.4. Collinearity Analysis of PbGLK Genes
The syntenic relationships between PbGLK genes and GLK genes from A. thaliana and O. sativa were determined using MCScanX software. TBtools was used for visualization.
4.5. Phylogenetic Analysis
The sequences of GLK proteins of P. bournei, A. thaliana, and O. sativa were aligned using the Muscle program of MEGA11 with default settings for constructing maximum likelihood (Bootstrap replications: 1000) phylogenetic tree [74,75]. iTOL (https://itol.embl.de/, accessed on 13 November 2024) was used to improve and beautify the phylogenetic tree.
4.6. Analysis of Conserved Motifs and Gene Structures
The protein sequence of P. bournei was identified using the online software MEME (https://meme-suite.org/meme/tools/meme, accessed on 9 October 2024), and the predicted value of the motif number was 10. The multiple GLK protein sequence alignment was carried out using Jalview software (version 2_11_4_0). We used a batch CD-search with default parameters to detect the conserved domains of the PbGLK proteins.
4.7. Promoter Cis-Element Analysis of PbGLK Genes
To explore the cis-acting elements in the sequence, we extracted the upstream 2000 bp sequences from the P. bournei genome. The online database PlantCARE was used to identify and analyze the cis-acting regulatory elements in the promoter region of the PbGLK genes. After selection and categorization, the data were visualized using TBtools software.
4.8. RNA Extraction and qRT-PCR Analysis
Total RNA extraction was carried out using an RNA Extraction Kit (Omega Bio TEK, Shanghai, China) for both the control and the stress-treated samples. Following the instructions of manufacturers, EasyScript One-step gDNA Removal and cDNA Synthesis SuperMix (Transgen, Beijing, China) were utilized to synthesize the cDNA. qRT-PCR was subsequently performed using TransStart top green qPCR SuperMix (Transgen, Beijing, China). The specific primers used in the qRT-PCR experiment were designed by TBtools and are listed in Table S2. The mixture solution of the qRT-PCR reaction is composed of 1 L of cDNA, 2 L of specific primers, 10 L of SYBR Premix Ex TaqTM II, and 7 L of ddH_2_O. The qRT-PCR reaction process was as follows: pre-degeneration at 95 °C for 30 s, then 40 cycles of denaturation at 95 °C for 5 s, 60 °C for 30 s, 95 °C for 5 s, 60 °C for 60 s, and 50 °C for 30 s. The internal reference gene was PbEF1α (GenBank No. KX682032) [76]. The relative expression of PbGLK genes was calculated using the 2^−∆∆CT^ method, and one-way ANOVA and Duncan multiple comparison tests were performed using the GraphPad Prism9.0 software (https://www.graphpad.com/) [77]. One-way ANOVA was conducted to determine whether there were significant differences among the means of the groups. Subsequently, Duncan’s multiple comparison test was performed to identify which specific groups exhibited significant differences. This test was chosen because it is suitable for comparing multiple groups with unequal sample sizes while effectively controlling the Type I error rate. After determining the experimental groups, differences among groups were annotated using the ABCD annotations. This method was selected because it is more appropriate for representing significance when dealing with multiple time points. To ensure robustness, all quantitative PCRs were conducted with three biological repeats and three technical replicates.
5. Conclusions
In this comprehensive study, we conducted a genome-wide analysis of the GLK gene family in P. bournei, elucidating its phylogenetic relationships, gene structures, and expression under various abiotic stresses. Our findings have significant implications for understanding the role of GLK genes in plant growth, chloroplast development, and stress adaptation, particularly in the context of woody plants.
Firstly, we identified 61 PbGLK genes in P. bournei, which were classified into seven subfamilies based on phylogenetic analysis. This classification was supported by differences in gene structure, conserved motif arrangement, and chromosomal distribution. Notably, the C subfamily contained the largest number of members, indicating its potential importance in the functional diversification of PbGLK genes. The uneven distribution of PbGLK genes across 12 chromosomes, along with evidence of segmental duplication events, suggests that gene duplication has played a crucial role in the expansion and functional evolution of this gene family.
Our analysis of cis-acting elements in the promoter regions of PbGLK genes showed a high prevalence of both hormone-related and light-responsive elements, with a particularly significant occurrence of light-responsive elements. This suggests that these genes are likely involved in regulating chloroplast development and photosynthesis under varying light conditions. This finding is further supported by the expression analysis, which showed that specific PbGLK genes, such as PbGLK25 and PbGLK30, exhibited significant upregulation under drought, heat, light, and shade stresses. These genes may serve as key regulators in enhancing stress tolerance and maintaining photosynthetic efficiency in P. bournei.
Although the implication of our findings is for fundamental research, given the near-threatened status of P. bournei and its ecological and economic importance in China, the identification of stress-responsive PbGLK genes provides a valuable genetic resource for the genetic improvement of this species. For instance, the overexpression of PbGLK25 and PbGLK30 could potentially be explored in breeding programs aimed at developing P. bournei varieties with enhanced tolerance to abiotic stresses, thereby contributing to forest conservation and sustainable forestry practices.
Moreover, our study highlights the conserved and divergent roles of GLK genes across different plant species. The synteny analysis with A. thaliana and O. sativa revealed both conserved and species-specific patterns, suggesting that the GLK gene family has undergone lineage-specific evolution while retaining core functions related to chloroplast development. This insight could guide comparative studies on GLK genes in other woody plants and crops, facilitating the identification of key regulatory genes involved in stress adaptation and growth optimization.
In conclusion, this study enhances our understanding of the GLK gene family in P. bournei and provides a foundation for future research on the molecular mechanisms underlying their functions. The identified stress-responsive PbGLK genes offer promising targets for genetic engineering and breeding efforts aimed at improving the resilience of P. bournei and other related species. Future work could focus on functional validation of these genes through transgenic approaches, as well as exploring their interactions with other regulatory networks involved in stress responses and chloroplast biogenesis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lu C.F. Liu X.Y. Tang Y.Q. Fu Y.Q. Zhang J.M. Yang L.T. Li P.H. Zhu Z.L. Dong P. A comprehensive review of TGA transcription factors in plant growth, stress responses, and beyond Int. J. Biol. Macromol.202425812888010.1016/j.ijbiomac.2023.12888038141713 · doi ↗ · pubmed ↗
- 2Yuan H.Y. Kagale S. Ferrie A.M.R. Multifaceted roles of transcription factors during plant embryogenesis Front. Plant Sci.202414132272810.3389/fpls.2023.132272838235196 PMC 10791896 · doi ↗ · pubmed ↗
- 3Bernal-Gallardo J.J. Zuñiga-Mayo V.M. Marsch-Martinez N. de Folter S. Novel Roles of SPATULA in the Control of Stomata and Trichome Number, and Anthocyanin Biosynthesis Plants 20231259610.3390/plants 1203059636771679 PMC 9919660 · doi ↗ · pubmed ↗
- 4Drewell R.A. Klonaros D. Dresch J.M. Transcription factor expression landscape in Drosophila embryonic cell lines BMC Genom.20242530710.1186/s 12864-024-10241-1PMC 1096099038521929 · doi ↗ · pubmed ↗
- 5Ma Z.M. Hu L.J. WRKY Transcription Factor Responses and Tolerance to Abiotic Stresses in Plants Int. J. Mol. Sci.202425684510.3390/ijms 2513684538999954 PMC 11241455 · doi ↗ · pubmed ↗
- 6Yan T.Y. Shu X.C. Ning C.L. Li Y.H. Wang Z. Wang T. Zhuang W.B. Functions and Regulatory Mechanisms of b HLH Transcription Factors during the Responses to Biotic and Abiotic Stresses in Woody Plants Plants 202413231510.3390/plants 1316231539204751 PMC 11360703 · doi ↗ · pubmed ↗
- 7Radani Y. Li R.X. Korboe H.M. Ma H.Y. Yang L.M. Transcriptional and Post-Translational Regulation of Plant b HLH Transcription Factors during the Response to Environmental Stresses Plants 202312211310.3390/plants 1211211337299095 PMC 10255293 · doi ↗ · pubmed ↗
- 8Dhatterwal P. Sharma N. Prasad M. Decoding the functionality of plant transcription factors J. Exp. Bot.2024754745475910.1093/jxb/erae 23138761104 · doi ↗ · pubmed ↗