The Mitochondrial Genome of the Springtail Semicerura bryophila (Collembola): New Data Call into Question the Relevance of the Subfamilies of the Isotomidae
Zhijng Xie, Mingxin Zheng, Yueying Li, Shiyu Du, Ruslan Saifutdinov, Mikhail Potapov, Xin Sun, Donghui Wu

TL;DR
This paper analyzes the mitochondrial genome of a springtail species, revealing new insights into the evolutionary relationships within the Isotomidae family.
Contribution
The study provides the first mitochondrial genome of Semicerura bryophila and challenges the classification of Isotomidae subfamilies.
Findings
The mitochondrial genome of S. bryophila is 15,247 bp long with typical gene content.
Phylogenetic analysis suggests Anurophorinae is more closely related to Semicerura and Folsomotoma than Isotominae.
Isotominae is found to be paraphyletic, challenging current taxonomic groupings.
Abstract
Background: Semicerura bryophila Potapov & Sun, 2020 is a soil-dwelling springtail belonging to the family Isotomidae. The phylogenetic relationships among species of this group remain controversial due to a lack of molecular data. Therefore, in this study, we sequenced the mitochondrial genome of S. bryophila, analyzed the characterization of the mitochondrial genome, and investigated the phylogenetic relationships of Isotomidae. Methods: The mitochondrial genome of S. bryophila was sequenced and assembled. We analyzed the sequence length, nucleotide composition, and evolutionary relationships within the Isotomidae family, incorporating data from twelve previously published mitochondrial genomes. Results: The length of the S. bryophila mitogenome is 15,247 bp and comprises 13 protein-coding genes, 22 tRNAs, and two rRNAs, arranged in a typical order. Its base composition is as follows:…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1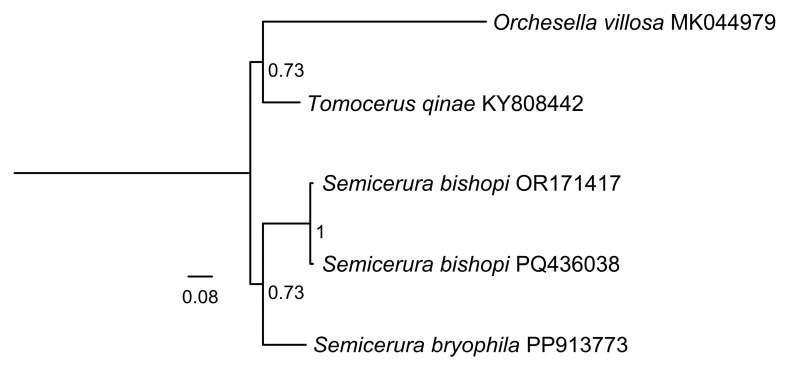 Figure 2
Figure 2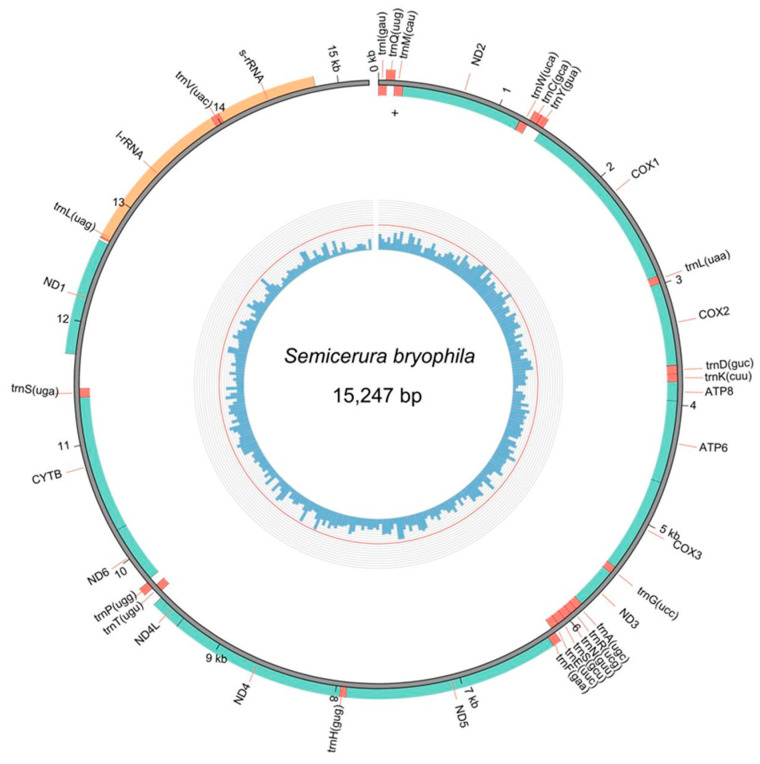 Figure 3
Figure 3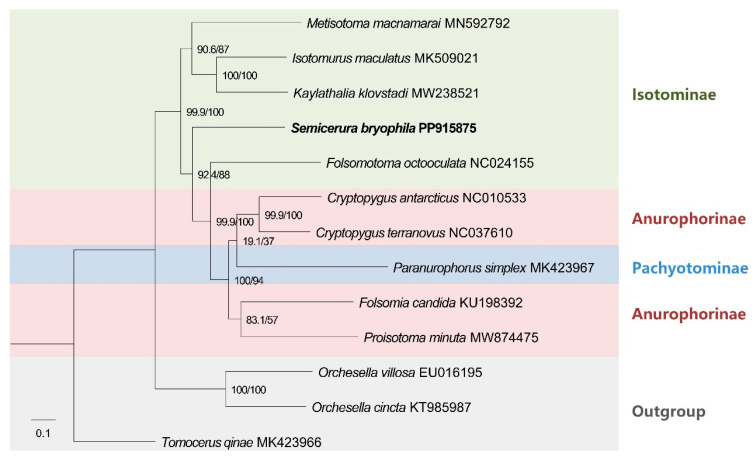 Figure 4
Figure 4- —National Natural Science Foundation for Young Scientists of China
- —Fundamental Research Funds for the Central Universities
- —National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCollembola Taxonomy and Ecology Studies · Study of Mite Species · Invertebrate Taxonomy and Ecology
1. Introduction
Collembola (springtails) are a diverse, abundant, and widespread group that plays a key role in terrestrial ecosystem processes, including carbon and nitrogen cycling, soil microstructure formation, and plant litter decomposition [1,2,3]. Over 9000 species have been described worldwide although it is difficult to give an exact figure as there are many species yet to be discovered [1]. Collembola have an extensive global distribution [4], they are found on every continent, including Antarctica, demonstrating their ability to inhabit a wide range of environments, from tropical forests to polar regions. Their presence in Antarctica is particularly notable, as it represents one of the most extreme and inhospitable environments for life.
The family Isotomidae Börner, 1913 [5], is one the most abundant and widespread in virtually any habitat of the Palaearctic region [6] with 1484 species recorded worldwide (as of 26 February 2025) [4]. Currently, three subfamilies are recognized within the family: Isotominae, Anurophorinae, and Pachyotominae. Among these, the genus Semicerura presumably belongs to Isotominae, although it displays several uncommon morphological features for the subfamily [7]. The genus is distributed in North America and East Asia and currently comprises five species [4]. The recently described S. bryophila is known to inhabit diverse ecosystems across eastern Asia, ranging from lowlands to alpine regions, and it shows a clear habitat preference for moss [4]. To explore the evolution of the genus within Isotomidae and to validate the subfamilies, the mitochondrial genome of S. bryophila Potapov & Sun, 2020 was sequenced, assembled, and annotated, representing the first mitogenome of the genus.
2. Materials and Methods
Soil samples containing specimens of S. bryophila were collected in May 2015 using a soil corer from the Changbai Mountains in Jilin province, northeast China. Individuals were extracted from soil using Berlese funnels (diameter 20 cm, mesh size 0.84 mm) over ten days without heating, and preserved in 95% ethanol for further analysis. In total approximately 300 specimens were collected from multiple elevations within the Changbai Mountains: 950 m (41.858° N, 127.748° E), 1100 m (41.847° N, 127.798° E), and 1700 m (41.758° N, 127.939° E). At the 1100 m, this elevation appears to be the most favorable for the S. bryophila, as it had the highest abundance of specimens collected (approximately 200 individuals, leg. Donghui Wu). This suggests that environmental conditions at this elevation, such as temperature, humidity, vegetation, or other ecological factors, are optimal for their survival and reproduction. Consequently, specimens from this elevation were selected for morphological identification and molecular analyses. The vegetation at 1100 m is characterized by mixed coniferous forests, with dominant tree species including Pinus koraiensis, Picea jezoensis var. microsperma, Abies nephrolepis, and Larix olgensis. The preserved specimens were imaged using a Zeiss STEMI 508 stereo microscope (Carl Zeiss AG, Oberkochen, Germany) equipped with a DS-Fi1 camera (Figure 1). One specimen was deposited at the Nanjing Agricultural University, Nanjing, China (https://faculty.njau.edu.cn/zhangfeng/zh_CN/ (accessed on 2 February 2025), Prof. Dr. Feng Zhang, [email protected]) under the voucher number ‘B4’.
The voucher specimen was cleared in lactic acid and then mounted in cavity and flat slides with Gisin’s liquid and Marc André II solution, respectively. Morphology was studied under a Nikon Eclipse 80i microscope (Nikon Corporation, Tokyo, Japan). Species identification was based on the contrasting coloration (black trunk versus yellow-white legs), the number of spines on the dens, and sparsely ciliated strong macrochaetae [7]. It was identified by comparing it with the holotype of S. bishopi Maynard, 1951, kept in the Smithsonian National Museum of Natural History. This comparison focused on key diagnostic characters, including chaetotaxy (e.g., the number and arrangement of chaetae on the body and appendages), spine patterns on the dens (the presence of 2 + 2 short spine-like chaetae in S. bryophila versus 1 + 1 in S. bishopi), labial palp chaetae (5 basomedian chaetae in S. bryophila versus 4 in S. bishopi), and other morphological features such as the number of acuminate apical chaetae on Legs I–III (8, 9, 9 in S. bryophila versus 8, 8, 8 in S. bishopi). To further strengthen the results of identification process, we incorporated molecular tools for verification.
Specifically, we downloaded the cox1 sequences of S. bishopi from NCBI, amplified and sequenced the corresponding region for our S. bryophila specimens, and compared the resulting sequences. Sequences were aligned and manually checked and corrected, resulting in a final 658 bp segment of the Cytochrome c oxidase subunit I (COI) gene, with Orchesella villosa and Tomocerus qinae used as outgroups. Maximum likelihood tree was conducted using MEGA version X [8] with the Kimura-2 parameter model [9] and bootstrap method with 1000 replicates. The phylogenetic tree illustrates the genetic relationships among the studied specimens (Figure 2), including S. bryophila and S. bishopi. The tree highlights S. bryophila as a distinct lineage, further confirming the genetic distinction between S. bryophila and S. bishopi, and supporting the morphological findings. This combined approach—morphological and molecular—provides a robust and comprehensive method for species identification, particularly in cases where morphological differences are subtle or where geographical variation may obscure diagnostic traits.
2.1. DNA Extraction
DNA was extracted from an individual collected in Changbai Mountains, northeast China (41.847° N, 127.798° E; altitude ca 1100 m; May 2015, leg. Donghui Wu). The sample was deposited at Nanjing Agricultural University (BioSample accession SAMN41817341). DNA extraction was performed using the Ezup Column Animal Genomic DNA Purification Kit (Sangon Biotech, Shanghai, China) following the manufacturer’s protocol. The genomic DNA was stored at −80 °C prior to analysis.
2.2. Mitogenome Sequencing, Assembly, and Annotation
The cytochrome c oxidase subunit I (cox1) gene was amplified using primers LCO1490 and HCO2198 [10], following the protocol of Zhang et al. (2014) [11]. PCR products were visualized using 1% agarose gel electrophoresis, then purified and sequenced by Majorbio (Shanghai, China) using the ABI 3730XL DNA Analyzer (Applied Biosystems, Waltham, MA, USA). Sequences were assembled using Sequencher 4.5 (Gene Codes Corporation, Ann Arbor, MI, USA), and aligned using MEGA 7.0 [12]. The resulting cox1 alignment, spanning 658 bp, was used as the seed sequence for the assembly (cox1 accession number: PP913773).
The DNA concentration was quantified using a Qubit 3.0 with the Q33230 Qubit™ 1X dsDNA HS Assay Kit (Thermo Fisher Scientific, Waltham, MA, USA). A DNA pool was created by combining equal concentrations of DNA from the target species with seven other species. Libraries were sequenced with 350 bp insert sizes on the HiSeq X Ten platform (Tianjin Novogene Bioinformatics Technology Co., Ltd., Tianjin, China), producing 150 bp paired-end reads. The average read depth for the assembled mitochondrial genome was 279.02 X. Non-mitochondrial reads were filtered out using NextGenMap 0.5.5 [13] and SAMtools 0.1.18 [14]. Mitogenome assembly was conducted with NOVOPlasty v2.7.0 [15] using the cox1 sequence as a seed. Chimeric sequences were identified using VSEARCH 2.2.0 [16]. Annotations were performed with the MitoZ v2.4 [17]. The annotated genome sequence was deposited in GenBank under accession numbers: PP915875.
2.3. Phylogenetic Analysis
All published mitochondrial genes of Isotomidae were included in our analysis to provide novel insights into the family’s classification (Figure 3). Mitochondrial protein-coding gene (PCG) amino acid sequences were aligned with MAFFT v7.394 [18] and trimmed using trimAL v1.4 [19]. Supermatrices of 13 PCGs were constructed using FASconCAT-G v1.04 [20]. Maximum likelihood (ML) phylogenetic analysis on sequences from ten Isotomidae species: five species from the subfamily Isotominae (Folsomotoma octooculata, Isotomurus maculatus, Kaylathalia klovstadi, Metisotoma macnamarai, and S. bryophila), four from Anurophorinae (Cryptopygus antarcticus, C. terranovus, Folsomia candida, and Proisotoma minuta), and one from Pachyotominae (Paranurophorus simplex). Two Entomobryoidea species (Orchesella cincta, O. villosa) and one Tomoceroidea species (T. qinae) were used as outgroups. ModelFinder determined models for the supermatrices [21], analyzed in IQ-TREE version 1.6.12 [22] with 1000 replicates for both UFBoot (ultrafast bootstrap approximation) and SH-aLRT (SH-like approximate likelihood ratio) test to assess branch support values [23,24]. The phylogenetic tree was edited using FigTree version 1.4.4 (available at http://tree.bio.ed.ac.uk/software/figtree/, accessed on 2 February 2025).
3. Results
3.1. Genome Organization and Composition
The mitogenome of S. bryophila is 15,247 bp in length (Figure 3), which is comparable to species from the Isotomidae family (Table 1), which typically range from 15,147 bp (F. candida) to 15,930 bp (P. minuta). Similarly, in related families such as Tomoceridae and Orchesellidae, mitogenome lengths vary from 14,924 bp (O. villosa) to 15,728 bp (O. cincta), indicating a general consistency in mitogenome size across Collembola. The mitogenome of S. bryophila comprises 13 protein-coding genes (PCGs), 22 transfer RNA (tRNA) genes, two ribosomal RNA (rRNA) genes (Table 2). The base composition is 38.05% adenine, 33.64% thymine, 10.17% guanine, and 15.03% cytosine. Additionally, the sequenced portion lacks the A+T-rich region, although a 436 bp non-coding sequence is present at the junction between the 12S rRNA and trnI genes. Although the sequence is already available in GenBank (accession number PP915875), we were the first to determine the species name and reviewed the published taxonomic [7] and ecological [25] literature related to this species.
3.2. Phylogeny of Collembola
The phylogenetic analysis (Figure 4) confirmed the monophyly of Isotomidae, including the subfamilies Isotominae, Anurophorinae, and Pachyotominae, with Entomobryoidae and Tomoceroidea as outgroups. This finding aligns with the traditional classification of Isotomidae as a distinct family within Collembola.
The subfamily Isotominae appears to be paraphyletic, as Metisotoma, Isotomurus and Kaylathalia clustered together, while F. octoculata does not group with them. Instead, Folsomotoma shows a closer relationship to Anurophorinae (C. antarcticus, C. terranovus, F. candida, and P. minuta). This challenges the traditional classification of Isotominae based on morphological traits. S. bryophila clusters outside the core Isotominae group and is more closely related to Anurophorinae and Pachyotominae (P. simplex). This finding suggests that Semicerura may not belong to Isotominae, despite its morphological similarities, and highlights the need for a re-evaluation of its taxonomic placement.
The subfamily Anurophorinae is validated as a monophyletic group. However, Pachyotominae is nested within Anurophorinae, suggesting that it may not deserve separate recognition as a distinct subfamily. This finding is consistent with recent morphological studies that question the distinctiveness of Pachyotominae.
4. Discussion
The first mitogenome of the species S. bryophila comprises 13 PCGs genes, 22 tRNA genes, two rRNA genes, which is typical for metazoans [37,38]. The mitogenome length of S. bryophila (15,247 bp) is consistent with the typical size range observed in Isotomidae and related families of Collembola. This suggests that the mitogenome structure and organization in S. bryophila are similar to those of other Collembola, with minor variations likely due to, e.g., species-specific evolutionary adaptations. The comparative data highlight the stability of mitogenome size within Collembola, despite some exceptions such as P. simplex (9518 bp), which may reflect unique genomic features or assembly artifacts.
According to the modern and widely accepted classification of Isotomidae [6], the three subfamilies primarily differ in the anatomy of the furca: Isotominae have a long furca with slender, crenulated dens; while in Pachyotominae and Anurophorinae the furca may be middle-sized, short or absent. When present, Pachyotominae exhibit stout, non-crenulated dens. Additionally, the body cuticle is smooth in Isotominae and Anurophorinae but granulated in Pachyotominae.
In our phylogenetic reconstruction, the subfamily Isotominae appears to be paraphyletic, suggesting that its morphological diagnosis should be reconsidered in future studies. The genera Folsomotoma and Semicerura cluster outside the reference Isotominae (comprising Metisotoma, Isotomurus and Kaylathalia) and are closely related to Anurophorinae (C. antarcticus, C. terranovus, F. candida, and P. minuta) and Pachyotominae (P. simplex). Rather than forming a distinct clade, Folsomotoma and Semicerura exhibit a paraphyletic grouping within Anurophorinae, challenging their traditional placement within Isotominae. This moderate relationship is partially supported by morphological evidence, as both Folsomotoma and Semicerura tend to develop spines on the furca. However, despite Semicerura exhibiting several unique and rare morphological traits [7], none of these features are shared with Anurophorinae. In addition, genera within the subfamily Isotominae appear to be largely separated latitudinally, with Semicerura predominantly restricted to the Northern Hemisphere, while Folsomotoma is found exclusively in the Southern Hemisphere [39].
The paraphyletic grouping of Semicerura and Folsomotoma aligns with findings from previous molecular studies, which also suggest that Isotominae may not be monophyletic [40,41,42]. Earlier phylogenetic analyses have shown that some genera traditionally placed in Isotominae (e.g., Folsomotoma) exhibit closer genetic relationships to Anurophorinae, further challenging the current classification [26]. This finding challenges the current classification of these genera and raises important questions about the morphological and genetic relationships within Isotomidae. Our results suggest that Isotominae may need to be redefined, excluding Semicerura and Folsomotoma, which appear more closely related to Anurophorinae. Additionally, the morphological traits historically used to define Isotominae (e.g., furca structure) may not be reliable diagnostic characters, as they seem to have evolved convergently in different lineages. Therefore, a revised classification system integrating both morphological and molecular data will be essential to accurately reflect evolutionary relationships within Isotomidae.
The independence of Pachyotominae is not supported by our results—Paranurophorus appears as a typical member of Anurophorinae in the phylogenetic tree. This finding is consistent with recent morphological studies showing that Pachyotominae does not consistently exhibit numerous s-setae on the body surface [43]. This character was one of the differentiated characters in primary diagnosis of the family [3,44]. Secondary granulation of body cuticle in Pachyotominae was also previously considered a reliable diagnostic trait until the study by Potapov et al. [45], which demonstrated that the genus Isotopenola also exhibits a granulated cuticle, despite belonging to Anurophorinae. However, it is important to note that only one species from Pachyotominae (P. simplex) has been sequenced to date in NCBI. This limited representation restricts our ability to draw definitive conclusions about the subfamily’s phylogenetic status. Future work should include additional taxa from Pachyotominae, as well as other underrepresented subfamilies, to better resolve their evolutionary relationships and clarify the taxonomic boundaries within Isotomidae.
5. Conclusions
This study represents the first mitogenome of S. bryophila. All 37 genes of the S. bryophila mitogenome were encoded on the heavy chain, and the gene order of 13 PCGs was completely consistent with that of all known Collembola sequences. To provide novel insights into the family’s classification, all published mitochondrial genes of Isotomidae were included in our analysis. S. bryophila is more closely related to the subfamily Anurophorinae than to Isotominae. The monophyly of subfamily Anurophorinae was validated, Pachyotominae was found to be a part of Anurophorinae, and Isotominae appeared to be paraphyletic. The first mitochondrial genome of the springtail S. bryophila (Collembola, Isotomidae) challenges the relevance of the current subfamilies within the family. Further analyses incorporating more mitochondrial DNA sequences from these three subfamilies are needed to better resolve the evolutionary relationships, as the current support values are insufficient to draw definitive conclusions. These findings highlight the need for a revised classification system that integrates both morphological and molecular data.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hopkin S.P. Biology of the Springtails: (Insecta: Collembola)OUP Oxford Oxford, UK 1997
- 2Rusek J. Biodiversity of Collembola and Their Functional Role in the Ecosystem Biodivers. Conserv.199871207121910.1023/A:1008887817883 · doi ↗
- 3Deharveng L. Recent Advances in Collembola Systematics Pedobiologia 20044841543310.1016/j.pedobi.2004.08.001 · doi ↗
- 4Bellinger P.F. Christiansen K. Janssens F. Checklist of the Collembola of the World Available online: https://www.collembola.org/(accessed on 2 February 2025)
- 5Carl B. Die Familien Der Collembolen Zool. Anz.191341315322
- 6Potapov M. Synopses on Palaearctic Collembola: Isotomidae Senckenberg Museum of Natural History Görlitz Görlitz, Germany 2001
- 7Potapov M. Xie Z. Kuprin A. Sun X. The Genus Semicerura (Collembola; Isotomidae) in Asia Zootaxa 2020475110511810.11646/zootaxa.4751.1.532230433 · doi ↗ · pubmed ↗
- 8Kumar S. Stecher G. Li M. Knyaz C. Tamura K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms Mol. Biol. Evol.2018351547154910.1093/molbev/msy 09629722887 PMC 5967553 · doi ↗ · pubmed ↗