MicroRNAs in the Mitochondria–Telomere Axis: Novel Insights into Cancer Development and Potential Therapeutic Targets
José Alfonso Cruz-Ramos, Emmanuel de la Mora-Jiménez, Beatriz Alejandra Llanes-Cervantes, Miguel Ángel Damián-Mejía

TL;DR
This paper reviews how microRNAs influence mitochondria and telomeres, impacting cancer development and possible treatments.
Contribution
It highlights novel roles of miRNAs in the mitochondria–telomere axis and their potential as cancer biomarkers or therapies.
Findings
miRNAs regulate telomeric stability and mitochondrial behavior, affecting cancer cell proliferation.
MitomiRs influence the cGAS/STING pathway, linking mitochondrial DNA to immune responses in cancer.
miRNAs are potential therapeutic targets due to their roles in metabolic changes and oncogenesis.
Abstract
The mitochondria–telomere axis is recognized as an important factor in the processes of metabolism, aging and oncogenesis. MicroRNAs (miRNAs) play an essential function in this complex interaction, having an impact on aspects such as cellular homeostasis, oxidative responses and apoptosis. In recent years, miRNAs have been found to be crucial for telomeric stability, as well as for mitochondrial behavior, factors that influence cell proliferation and viability. Furthermore, mitochondrial miRNAs (mitomiRs) are associated with gene expression and the activity of the cGAS/STING pathway activity, linking mitochondrial DNA recognition to immune system responses. Hence, miRNAs maintain a link to mitochondrial biogenesis, metabolic changes in cancer and cellular organelles. This review focuses on the roles of a variety of miRNAs in cancer progression and their potential application as…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
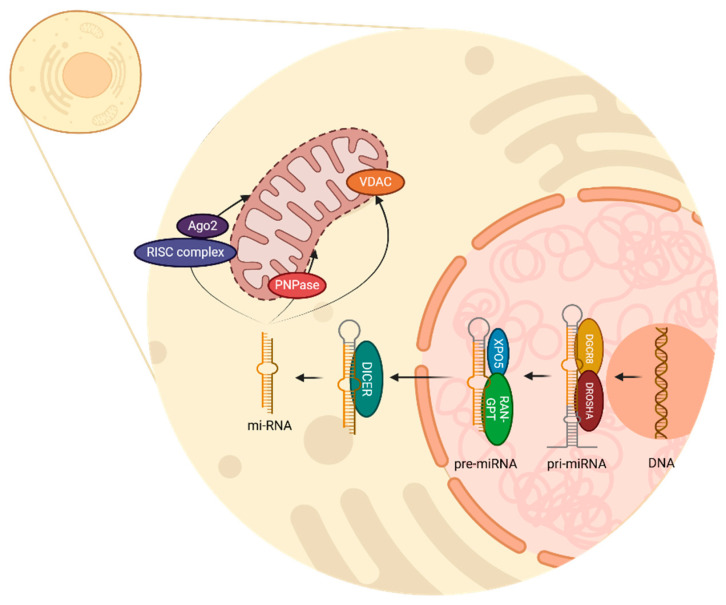 Figure 1
Figure 1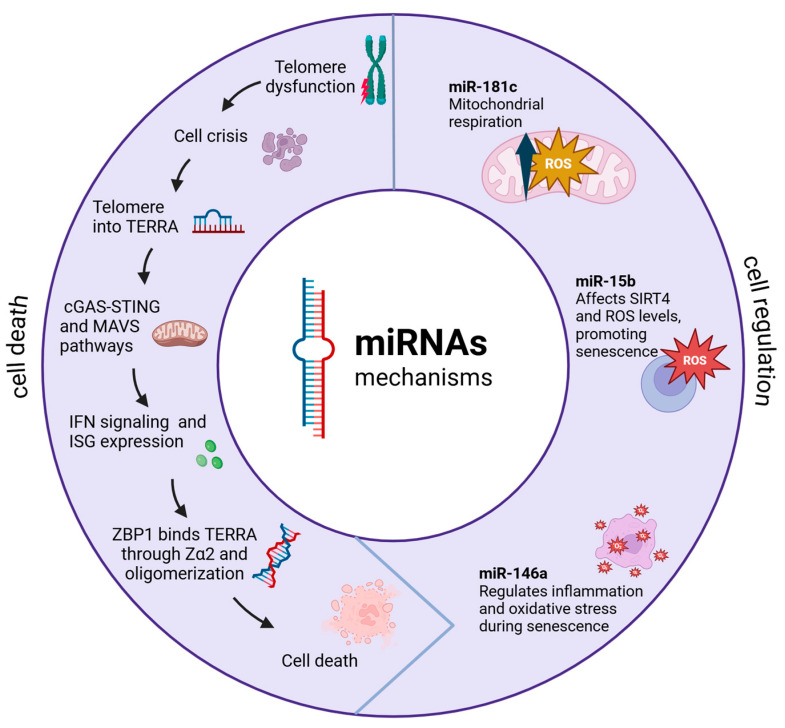 Figure 2
Figure 2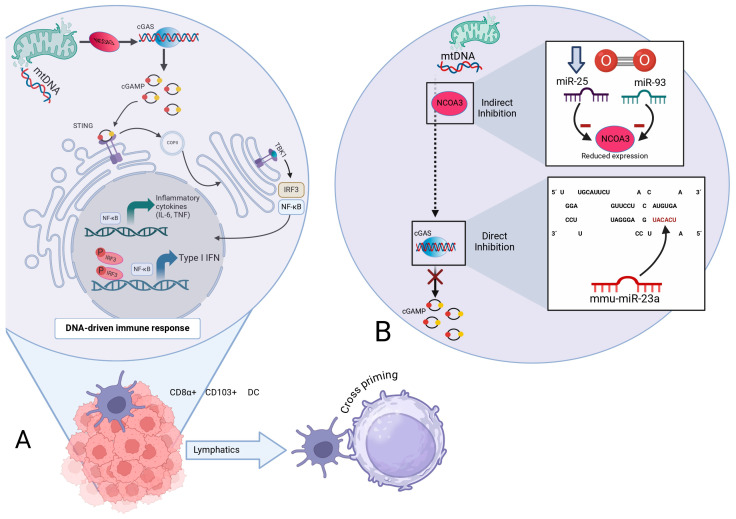 Figure 3
Figure 3- —APPAC-VII 2025
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
Topicsinterferon and immune responses · MicroRNA in disease regulation · RNA modifications and cancer
1. Introduction
Mitochondria regulate the metabolism and many cellular processes, while telomeres are responsible for controlling the lifespan of the cell; impaired function in either can trigger several pathways to failure, leading to a cascade of dysfunctional events within the cell [1]. Consequently, the interaction between telomeres and mitochondria has been implicated in maintaining cellular homeostasis, contributing to the development of neoplasms and oncogenic processes, emerging as an important field of research [2].
On the other hand, it has been described that the biological behavior of a cell is mediated by the interaction of ribonucleic acid (RNA) with telomeres, specifically through telomeric repeat-containing RNA (TERRA), a non-coding RNA (ncRNA) which acquires an important function in the regulation and maintenance of telomeres [3].
This has led to research into the importance of ncRNAs as regulators in cell processes, emerging as a strategy for the modulation of diverse cellular pathways, such as miRNAs, which have demonstrated active participation in messenger RNA (mRNA) regulation, by binding to the 3’untranslated region (3’ UTR) [4,5]. miRNAs consist of small sequences of about 21 to 23 nucleotides, and their coding genes are transcribed by RNA polymerase II [5].
During malignant cellular transformation, miRNAs play key roles in cell differentiation by regulating cancer stem cells and tumor development [6]. Additionally, the molecular heterogeneity of cancer and its variability in the response to medical treatment may contribute to the investigation of novel therapies and diagnostic models, since even single-nucleotide polymorphisms modulate miRNA expression in some cancers, for example breast cancer, demonstrating a wide field of research [7,8].
Understanding the mechanisms involved in the complex axis between miRNAs, telomeres and mitochondria is indispensable for the development of novel approaches to cancer treatment strategies.
2. Cellular Organization and Energy Metabolism
Cellular organelles in eukaryotes are highly complex and maintain complex bidirectional signaling relationships with each other and with other cellular structures both functionally and structurally. The signaling pathways result in cellular metabolism regulation and evolution towards a healthy state, cell death, or malignant transformation [9]. Smaller molecules have the advantage of being able to diffuse more easily across the cell membrane compared to larger molecules. However, within the cell, specifically in the mitochondria, membranes exhibit a highly complex structure, which is closely related to their unique energy production capacity. This structural complexity not only supports energy generation but is also fundamental for cellular metabolism and the determination of cell fate, whether tending towards apoptosis, carcinogenesis, or senescence, among other cellular processes [10]. Mitochondrial membranes interact in a distinct manner with molecules and mediators, compared to the cytoplasmic membrane. In mitochondria, the membrane folds or cristae increase the surface area for adenosine triphosphate (ATP) production during electron transport in the mitochondrial respiratory chain, as well as other forms of metabolic regulation and cell proliferation [11].
Within the mitochondrial matrix resides the mitochondrial genome, which is structurally distinct from nuclear DNA. The mitochondrial DNA (mtDNA) in mammals is a small, circular genome of about 16.5 kb, encoding 13 subunits of the oxidative phosphorylation (OXPHOS) system, as well as the transfer RNAs (tRNAs) and ribosomal (rRNAs) crucial for their synthesis. The mtDNA system, which resembles bacterial DNA in its organization, is capable of synthesizing proteins that operate in close coordination with nuclear-encoded proteins imported into the mitochondria [12].
The electron transport complexes (I through V) in the inner mitochondrial membrane generate ATP mostly through OXPHOS. This process results in the formation of reactive oxygen species (ROS) as a byproduct of biochemical reactions in ATP production [13]. Additionally, mitochondria are associated with several pathways outside the tricarboxylic acid (TCA) cycle, including fatty acid oxidation and amino acid metabolism. The varying energy requirements should in principle be met by the cell through plasticity in metabolic pathways to maintain homeostasis and survive in changing local microenvironments. In this sense, the basic canonical mechanisms observed under normal conditions undergo modifications that lead to metabolic shifts in energy systems. These alterations, which are characteristic of cancer cells, include increased aerobic glycolysis and lactic acid production. Such metabolic reprogramming supports tumor progression and enhances the survival of neoplastic cells [14].
3. The miRNA: Biogenesis in Animal Cells and Its Relationship with Mitochondria
The miRNAs have been described in different species such as viruses, animals, and plants [15]. Nomenclature to classify miRNAs remains undefined; however, it has been described that miRNAs with identical sequences at nucleotides 2–8 of the mature miRNA will appertain to the same family [16].
Furthermore, to maintain a consensus for miRNA characterization, specific criteria are considered in addition to length; there must be a relationship with the Argonaute proteins, the synthesis must be mediated by a ribonuclease (RNase) type III, and there must be phylogenetic conservation, an abundance of sequence reads and the homogeneous presence of the 5’ end [15,16].
Several studies have elucidated these processes by studying various animal species, mainly humans, Caenorhabditis elegans (C. elegans), Drosophila melanogaster, and Giardia intestinalis [15,16]. Moreover, phylogenetic conservation among mammals has been demonstrated [16,17].
However, it is important to mention the differences between miRNA biogenesis in animals and plants. In plants, it has been observed that the processing takes place entirely in the nucleus and unlike in animals, and there is no homolog of Drosha and DiGeorge syndrome critical region gene 8 (DGCR8); therefore, most of the processes are mediated by DICER-LIKE 1, another type III RNase [16]. In this review, we describe miRNA biogenesis in animal cells, focusing on human cells.
miRNA biogenesis begins in the cell nucleus with the transcription of a DNA strand to produce primary miRNA (pri-miRNA) by RNA polymerase II [16]. This primary transcription is mediated by the complex of Drosha and DGCR8, which acts as an enzyme and removes the non-structured endings of the pri-miRNA, therefore creating pre-miRNA as a precursor [16,18]. Drosha is an RNase type III which cuts pri-miRNA; meanwhile, DGCR8 acts as a cofactor through which to recognize and stabilize pri-miRNA structure [19].
Once pre-miRNA is recognized by exportin 5 (XPO5), and Ras-related nuclear protein bound with Guanosine Triphosphate (RAN-GTP) provides the necessary energy, pre-miRNA is transported from the nucleus to the cytoplasm. Then, the RNase type III DICER, recognizes and processes the chain by cutting the hairpin; thus, a double-stranded miRNA of approximately 22 nucleotides is generated [16,18]. This miRNA duplex has two strands, the mature miRNA, which will be the functional guide strand, and the passenger strand, which will be degraded.
In the cytoplasm, miRNA can conjugate with various complexes and regulate important functions. When incorporated into the RNA-induced silencing complex (RISC) and Argonaute 2 (Ago2) protein, miRNA can identify complementary sequences in mRNA and regulate two mechanisms: mRNA degradation and translation inhibition [20]. Additionally, it is known that several miRNAs can be found inside the mitochondria, accomplishing regulatory processes in mitochondrial genes. Polynucleotide phosphorylase (PNPase) facilitates the importation of miRNA into mitochondria, and the voltage-dependent anion channel (VDAC) participates in RNA and protein transportation through the external mitochondrion membrane [21,22]. All these processes are illustrated in Figure 1.
Notably, most of the miRNAs known are synthesized outside the mitochondrion, in other words, they are nuclear miRNAs with targets in the mitochondrion. Nevertheless, the difference in the process of miRNAs synthesized within the mitochondria compared to the process of those encoded in the nucleus remains unclear [23]. Once localized inside the mitochondrion, the presence of miRNAs can modulate its dynamics, which could lead to structural changes, resulting in alterations in fission or fusion, causing damage to the organism [24].
4. Fusion and Fission Mitochondrial Processes
Mitochondria are complex organelles that can fuse and divide to some degree, similarly to bacterial cell division. Under certain conditions, mitochondria fuse specifically depending on the context, microenvironment, and cellular stress [10,25,26]. Golgi apparatus-dependent mitochondrial fusion is achieved through GTPases such as Mitofusin 1 (MFN1) and Mitofusin 2 (MFN2), which coordinate outer mitochondrial membrane fusion. On the other hand, Optic atrophy 1 (OPA1) protein is responsible for mediating inner mitochondrial membrane fusion [9,10,25].
Dysregulation of fusion and fission promotes the survival of malignant cells; dysfunctional mitochondria lead to abnormalities in both mitochondrial morphology and function [25,26]. Fusion processes are particularly important when metabolic demand is high, and therefore mitochondria can fuse and share components to maintain their functionality and energy efficiency [10,26].
The mitochondrial fission machinery relies on dynamin-related protein 1 (DRP1) and its receptors FIS1, MFF (mitochondrial gene-specific protein), and MiD49/51 to preserve mitochondrial morphology and their distribution in the cell matrix [9,10,25]. In cancer cells, aberrant fission is observed, which further promotes the proliferation and survival of malignant neoplastic cells, making it a stress response process [9,25,26].
5. Mitochondrial–Telomere Communications in Cancer
5.1. Molecular Basis of Crosstalk Communication Between the Mitochondria and Telomeres
The bidirectional interactions between mitochondria and telomeres at the molecular level contribute in a fundamental manner to cellular homeostasis and adaptation. This interaction is more complex than previously thought, particularly regarding oxidative stress and cellular signaling pathways [27,28].
A significant breakthrough in understanding this relationship comes from studies showing that mitochondrial H_2_O_2_ release and ROS production affect nuclear DNA and telomeres through sophisticated signaling mechanisms rather than direct oxidative damage, challenging previous assumptions about these interactions [27,29].
The telomere–mitochondria axis is particularly relevant in cellular senescence and aging. It has been shown that telomere damage influences mitochondrial function through specific signaling pathways, notably the p53-peroxisome proliferator-activated receptor gamma 1α coactivator (PGC-1α) pathway [1]. Furthermore, Sung et al. have shown that interventions targeting this axis, such as metformin treatment, can influence both telomere stability and mitochondrial function, potentially mitigating cellular senescence [30].
This complex interplay between telomeres and mitochondria represents a critical regulatory mechanism in cellular aging and disease progression, where dysfunction in either component can trigger a cascade of cellular responses affecting both structures [1,27]. Understanding these interactions has important implications for developing targeted therapeutic strategies for age-related diseases and cancer [29,30].
5.2. Mitochondrial–Telomere Communication via Non-Coding RNAs
ncRNAs are key in mediating communication among mitochondria and telomeres. Nassour et al. research has revealed sophisticated mechanisms through which these RNA molecules coordinate various cellular processes, particularly in the context of senescence and cancer [31].
A key discovery involves TERRA, which are long non-coding RNAs (lncRNAs) transcribed from dysfunctional telomeres [32]. These TERRA transcripts have been shown to interact specifically with ZBP1 (Z-DNA binding protein 1) on the outer mitochondria membrane, where they form distinct oligomeric structures [33,34].
Dysfunctional telomeres trigger cellular senescence through the activation of DNA damage response pathways. While senescence is mediated by p53 and retinoblastoma tumor suppressor (RB) pathways, cells with disrupted checkpoints bypass this protective mechanism [35]. Therefore, these cells enter a replicative crisis characterized by transcriptional changes due to an overlap of upregulated genes [31,32]. During crisis, telomeres undergo active transcription, producing TERRA: lncRNAs sequences containing UUAGGG repeats and subtelomeric-derived RNA [32]. TERRA has been involved in the sensing innate immune system pathways through the induction of interferon-stimulated genes (ISGs) [31]. Among ISG products, ZBP1 protein has arisen as a main mediator in cell death regulation and innate immunity through the induction of type I interferons (IFNs).
The synthesis of ZBP1 seems to be linked to the cyclic GMP-AMP synthase/stimulator of interferon genes (cGAS/STING) pathway, which becomes activated upon the cytosolic accumulation of nucleic acids. Furthermore, cGAS/STING-mediated DNA sensing amplifies ZBP1 expression [31,36].
The conformation of the ZBP1–TERRA complex through the Zα2 domain leads to the expression of mitochondrial antiviral signaling protein (MAVS), which leads to the activation of nuclear factor kappa–B (NF-kB) and interferon regulatory factor (IRF) 3/7 signaling 170 pathways, resulting in ISG expression [31,37]. Additionally, the complete activation of IFN-dependent ZBP1 requires the previous upregulation of ZBP1 by cGAS/STING and a signal from dysfunctional telomeres. When these conditions are fulfilled, an inflammatory loop is established, leading to enhanced ISG expression and to cell death [31]. This interaction triggers a cascade of events that can lead to programmed cell death in cancer cells, representing a novel tumor-suppressive mechanism; this mechanism is represented in Figure 2.
In this sense, miRNAs, particularly those targeting mitochondrial functions (mitomiRs), have emerged as important regulators of cellular metabolism and cancer progression. These mitochondria-localized miRNAs can originate from both nuclear and mitochondrial genomes, impacting various aspects of energy metabolism and cellular defense mechanisms [38]. Feng et al. have shown that specific mitomiRs are differentially expressed in various cancer types, affecting both mitochondrial function and the metabolic reprogramming of cancer cells [39].
5.3. Mitochondrial Nuclear-Encoded MitomiRs
mitomiRs act as essential post-transcriptional regulators. Kuo et al. have shown that mitomiRs require specific targeting mechanisms and specialized transport systems for proper mitochondrial localization [40]. Maurya et al. have demonstrated that these regulatory molecules play crucial roles in maintaining mitochondrial function and cellular homeostasis [41].
The regulation of mitochondrial function by nuclear-encoded mitomiRs occurs through multiple pathways. A well-documented example is miR-181c, which has been shown to significantly affect mitochondrial function by modulating ROS synthesis and glucose oxidation patterns in cancer cells. Luo et al. have identified its function as a tumor suppressor and its influence on drug responsiveness, highlighting that processes such as telomere attrition, oxidative stress, activation of tumor-suppressing pathways and mitochondrial impairment serve as primary triggers of cellular senescence [20,42].
Lang et al., in recent research, reported a link between miR-15b and ROS, where the inhibition of miR-15b induces mitochondrial ROS generation and reduces the mitochondrial membrane potential-dependent upregulation of SIRT4 [43]. On the other hand, miR-146a has been shown to have an important role in regulation processes regarding ROS production and inflammation, by interacting with elements of the NF-kB pathway and NADPH oxidase 4 (NOX4), where it functions as an anti-inflammatory and antioxidant, resulting in a decrease in oxidative stress [44].
Disruptions in the transport, assembly, or localization of mitochondrial elements may severely affect cellular homeostasis. For example, in tongue squamous cell carcinoma, an upregulation of miR-2392 has been demonstrated to specifically hinder mtDNA transcription, markedly decreasing the expression of COX1, ND4, and CYTB, ultimately inhibiting OXPHOS [20,45].
Additionally, a decline in miR-107 levels can result in mitochondrial dysfunction, characterized by diminished mitochondrial membrane potential and impaired electron transport chain (ETC) activity. Such alterations may profoundly influence key cellular processes, including resistance to drugs, inflammatory responses, and aging [20].
Alterations in mitomiRs expression are closely linked to cancer metabolism and mitochondrial stress signaling. The latest evidence, according to Maurya et al., demonstrates that tumor cells process mitochondrial ROS regulation to interact with various components in the tumor microenvironment, thereby affecting cancer progression [41]. Furthermore, Dasgupta et al. investigated a novel regulatory mechanism in mtDNA-less cells. Their analysis revealed a reduced expression of several mitochondrial RNAs, including COX1-3, 12S rRNA, and ND4-5. In contrast, Cyto b, ND1 and ND3 were entirely absent in 206 ρ° cells. This imbalance in mitochondrial RNA transcription indicates that post-transcriptional cleavage and processing mechanisms may be involved in controlling mitochondrial gene expression within these cells [46].
By partially base pairing with complementary sites in the cytosol, miRNAs are enabled to regulate mitochondrial RNA expression by aiming for specific mRNAs. This regulation of mitochondrial gene expression has significant implications for cellular energy metabolism [46,47].
5.4. Mitochondria-Encoded miRNAs
Mitochondrial-encoded miRNAs have recently begun to be studied, and while not much is known, it is understood that they are encoded in mtDNA and are processed by fine and specific machinery within the mitochondria [9]. These unique miRNAs have been shown to have distinct processing mechanisms that differ from their nuclear counterparts, suggesting specialized regulatory functions [20].
Initially, their existence was doubted; however, several studies have confirmed not only their existence but also their crucial participation in cellular metabolism and mitochondria–nucleus communication [46]. Yumeng et al. have emphasized the critical role of miRNAs in regulating both nuclear and mitochondrial proteins, thereby establishing a complex network of cellular regulations. Notably, abnormal methylation of mitochondrial RNA can impact mtDNA function by altering transcription stability and structure. Ribosomal RNA (rRNA) methylation is primarily regulated by mitochondrial rRNA methyltransferases, which include enzymes targeting both large (MRM1, MRM2, MRM3, TRMT61B) and small (TFB1M, TRMT2B, NSUN4, METTL15) ribosomal subunits [48,49].
Their processing is not traditional like nuclear miRNAs, suggesting a divergent evolution of these processing mechanisms. This evolutionary adaptation appears specifically aimed at regulating mitochondrial processes and facilitating communication with the nucleus [9,48]. Luo et al. have shown that these mitochondrial-specific miRNAs play crucial roles in maintaining cellular homeostasis and energy metabolism [20].
5.5. RNA-Dependent Modulation of the cGAS/STING Axis: Convergence of Mitochondrial Dynamics, Telomeric Integrity, and Programmed Cell Death Pathways
Pattern recognition receptors (PRRs) are of great importance in the innate immune system. PRRs recognize pathogen-associated molecular patterns (PAMPs) and host damage-associated molecular patterns (DAMPs) on the surface of innate immune cells [50].
The PRRs that have been mainly related to the cGAS/STING pathways are AIM2-like receptors (ALRs) and 2’-5’oligoadenylate synthetase-like receptors (OLRs) [51]. Once DNA is detected in the cell, ALR activates the STING-dependent ISG pathway. This mechanism was originally studied in antiviral responses [51,52]. OLRs, when activated in the cytoplasm by double-stranded nucleic acids, generate second messenger molecules like cGAMP. These molecules bind to and activate STING, triggering a downstream innate immune response [53].
cGAS activation enables the synthesis of cGAMP from GTP and ATP, which then binds to STING dimers localized in the endoplasmic reticulum (ER) membrane, resulting in their incorporation into coatomer protein complex II (COPII); then, it recruits and promotes the autophosphorylation of TANK-binding kinase 1 (TBK1), STING phosphorylation at Ser366, and the recruitment and further phosphorylation of IRF3, which is required for the induction of genes encoding IL-6 and IL-12. IRF3 dimerizes and translocates to the nucleus, ultimately leading to the expression of type I IFN and ISGs, boosting the cross-presentation of antigens to CD8+ T cells [54,55].
It was demonstrated that mitochondria experiencing minority mitochondrial outer-membrane permeability (miMOP) release mtDNA into the cytosol [56]. The release of mtDNA into the cell triggers the mtDNA-dependent cGAS/STING pathway, leading to the synthesis of type I IFN and inflammatory markers, which leads to the senescence-associated secretory phenotype (SASP) and inflammation [55,56].
It can be inferred that mtDNA-dependent cGAS/STING signaling is of great importance not only in senescence but also in certain illnesses such as cancer, where it has been shown to be key for antitumor immunity. STING activation is believed to promote tumor rejection by eliciting CD8+ T cell responses [57].
It promotes the enlistment of T cells and natural killer (NK) cells, into the malignancy microenvironment. Additionally, activation of the cGAS/STING signaling helps suppress the tumor metastasis capacity; as these cells attempt to spread, active STING signaling enhances immune recognition and destruction, thereby restricting the formation of new tumors [57,58,59].
miRNAs play an important role in regulating the cGAS/STING pathways. The 3’UTRs of cGAS/STING mRNA carry probable miRNA binding sites [51]. Yu et al. have stated that miRNAs can suppress the immune response through different mechanisms. For instance, certain miRNAs, such as miR-23a/b, directly bind to the 3’ UTR of cGAS, inhibiting its expression and thereby suppressing the cGAS-mediated innate immune response [60]. Indirectly, during hypoxia, miR-25 and miR-93 regulate cGAS expression by targeting NCOA3, a crucial epigenetic factor for maintaining cGAS expression levels. This regulation results in the downregulation of cGAS mRNA levels, facilitating hypoxic tumor cells to evade immune detection by modulating the cGAS/STING pathway [61], Figure 3.
6. Metabolic Reprogramming in Cancer
Malignant cells are profoundly reprogrammed in terms of energy metabolism; these changes are linked to and affect telomere and mitochondrial function. Aerobic glycolysis is prominently used by tumor cells; this has been called the Warburg effect [62]. Cancer cells maintain a balance between aerobic glycolysis, which involves glucose fermentation even in the presence of oxygen, and OXPHOS, which produces ATP through the oxidation of glucose carbon bridges [63].
The balance in energy acquisition is directed towards the prolonged cellular survival characteristics of certain malignant tumor clones. This type of mixed metabolism favors the maintenance of telomere integrity, characteristic of cancer cells [64]. Thus, this type of cancer cell metabolism promotes proliferation and telomere length maintenance through the modification of the TCA cycle, amino acid metabolism, and fatty acid oxidation [64].
The AMP-activated protein kinase (AMPK)–mammalian target of rapamycin (mTOR) axis regulates cellular energy metabolism, mitochondrial function, and telomere maintenance. This pathway is tightly dysregulated in cancer cells; therefore, these changes support both metabolism and cell survival [65,66]. Also, sirtuins (SIRT), particularly SIRT1 and SIRT3, act as metabolism-associated sensors that regulate mitochondria and telomeres in cellular stress responses and cancer cell adaptation to specific contexts [67].
Central to cellular metabolism regulation is the mTOR pathway, which acts aberrantly in cancer cells and has multiple effects on cellular metabolism. Indeed, mTOR and AMPK are regarded as metabolic sensors that relay cell survival signals under stress, and such crosstalk with mitochondrial function and telomere maintenance is essential for promoting cancer [68,69]. This means that cancer cell metabolic plasticity, mitochondrial and telomere structure and function changes play an important role in the survival of neoplastic cells and their proliferation [65].
The SIRT system modulates mitochondrial and telomere physiology through interaction with multiple molecules via histone protein deacetylation and the regulation of mitochondrial function and apoptosis. These proteins regulate cell proliferation and differentiation through various catalytic activities that depend on NAD+. Additionally, due to their deacetylase activity, they interact closely with telomeres, preventing their erosion and supporting their repair. Therefore, the processes of oncogenesis and tumor progression are closely linked to these axes and metabolic pathways related to AMPK, mTOR, and SIRTs, which in turn are precisely regulated by the large family of miRNAs. The complexity of these systems is very high, and their effects are dependent on the cellular microenvironment. Energy metabolism is strongly linked to the regulation of these highly specific molecular systems, with pleiotropic functions in most organs and systems, both under healthy and diseased conditions [70,71].
In hepatocellular carcinoma (HCC), SIRT1 is overexpressed and correlates with tumor grade, predicting adverse clinical outcomes and poor prognosis. miR-34a, decreased in severe HCC cases, has been shown, in mice, to induce tumor regression and prevent recurrence when expressed. Similarly, miRNA-29c acts as a tumor suppressor by inhibiting SIRT1 in HCC [72,73]. However, SIRTs also have antitumoral effects or dual effects (pro- and anti-tumoral) that can be context-dependent. For example, SIRT1 is associated with protective effects against tumor growth and promotes the cell death of tumor cells. It inhibits the proliferation of various types of cancer, such as ovarian, lung, and colorectal cancer [74].
Regarding the link between miRNA activity and metabolic reprogramming, it is closely associated with cancer cell transformation. The miR–200c–SIRT2 interaction axis plays a crucial role in establishing the metabolic shift known as the Warburg effect by regulating glycolytic enzymes [75].
In summary, metabolic reprogramming in cancer is regulated by miRNAs, which in turn regulate the canonical metabolic pathways of malignant transformation and progression such as mTOR, AMPK, and SIRT. These pathways reversely modulate cell cycle and fate, impacting mitochondrial structure and function and telomeric maintenance, confirming that the elucidation of these processes intrinsically involves investigating cancer reprogramming or the Warburg effect, demonstrating that the mitochondria-telomere axis is fundamental in this regulation, with miRNAs serving as the way that all these processes are fine-tuned.
7. Clinical Implications and Therapeutic Applications in Cancer
The clinical applications of miRNA appear to cover a vast range, and yet are largely unexplored. miRNAs play crucial roles in various diseases. Additionally, numerous miRNAs have been associated in degenerative diseases and cellular senescence, including increased expression of miR-34a, miR-21-22, miR-26b, miR-29 and miR-2010, as well as decreased expression of miR-106a, miR-19b, miR-20a and miR-320c [76].
Specifically, in the field of cancer research, miRNAs have garnered significant attention for their potential diagnostic, prognostic, and therapeutic applications.
The relationship between mitochondria and telomeres and their connection to senescence and malignant transformation processes represent opportunities for the diagnosis and monitoring of multiple diseases, especially oncological diseases. RNA species associated with oncological processes and cellular senescence can be useful as diagnostic markers and for monitoring disease progression, potentially resulting in increased specificity to traditional oncological markers [77,78].
In this sense, miRNAs are known to interact with several pathological conditions; high miR-181c expression is associated with mitochondrial dysfunction, and differences in miR-138 levels are associated with telomere abnormalities [76]. miRNAs, as an entire family of transcriptional regulators and molecular receptors, are likely to be useful in clinical assessments for the identification of improved subtypes and prognostication of oncological diseases [78].
In this regard, these molecules can also be used in RNA-based therapy, which can be directed at specific therapeutic targets and be used as regulators of mitochondria and telomeres. This represents a new frontier in personalized medicine, as miRNAs can be used to regulate metabolic pathways that can be utilized according to the context of the tumor type and the pathways that are altered [76,78].
The goal is to restore cellular functions, specifically mitochondria and telomeres, through synthetic RNA or regulatory molecules of these RNAs. In the case of miR-34a, mimicking molecules can be developed for the treatment of certain neoplasms to regulate mitochondrial functions and telomere maintenance, thereby restoring the homeostasis of both organelles [79,80].
8. Current Challenges and Future Directions
Developing miRNA-based therapeutics is challenging and needs to address the intrinsic stability of miRNA-based therapeutic agents and their delivery to the cellular compartments in which they are needed to modulate cellular functions. Nanoparticle delivery and other cell-specific transport methods have been applied for reduce side effects and enhance therapeutic efficacy [81,82].
Basic and clinical research is required to understand the complex network of molecular interactions of miRNAs with other RNA species. Bioinformatics and the development of highly complex and sophisticated computational models can shed light on the network of interactions between RNA species, telomeres, and mitochondria, thereby enabling the development of personalized therapies [83,84].
Moreover, integrating artificial intelligence methods and machine learning can lead to precise predictions and characterizations of different types of RNA and their relationship with mitochondrial and telomeric function and structure [85,86].
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gao X. Yu X. Zhang C. Wang Y. Sun Y. Sun H. Zhang H. Shi Y. He X. Telomeres and Mitochondrial Metabolism: Implications for Cellular Senescence and Age-Related Diseases Stem Cell Rev. Rep.2022182315232710.1007/s 12015-022-10370-835460064 PMC 9033418 · doi ↗ · pubmed ↗
- 2Assalve G. Lunetti P. Rocca M.S. Cosci I. Di Nisio A. Ferlin A. Zara V. Ferramosca A. Exploring the Link Between Telomeres and Mitochondria: Mechanisms and Implications in Different Cell Types Int. J. Mol. Sci.20252699310.3390/ijms 2603099339940762 PMC 11817679 · doi ↗ · pubmed ↗
- 3Cusanelli E. Chartrand P. Telomeric Repeat-Containing RNA TERRA: A Noncoding RNA Connecting Telomere Biology to Genome Integrity Front. Genet.2015614310.3389/fgene.2015.0014325926849 PMC 4396414 · doi ↗ · pubmed ↗
- 4Brosnan C.A. Voinnet O. The Long and the Short of Noncoding RN As Curr. Opin. Cell Biol.20092141642510.1016/j.ceb.2009.04.00119447594 · doi ↗ · pubmed ↗
- 5Slack F.J. Chinnaiyan A.M. The Role of Non-Coding RN As in Oncology Cell 20191791033105510.1016/j.cell.2019.10.01731730848 PMC 7347159 · doi ↗ · pubmed ↗
- 6Khan A.Q. Ahmed E.I. Elareer N.R. Junejo K. Steinhoff M. Uddin S. Role of mi RNA-Regulated Cancer Stem Cells in the Pathogenesis of Human Malignancies Cells 2019884010.3390/cells 808084031530793 PMC 6721829 · doi ↗ · pubmed ↗
- 7Lehmann B.D. JovanovićB. Chen X. Estrada M.V. Johnson K.N. Shyr Y. Moses H.L. Sanders M.E. Pietenpol J.A. Refinement of Triple-Negative Breast Cancer Molecular Subtypes: Implications for Neoadjuvant Chemotherapy Selection P Lo S ONE 201611 e 015736810.1371/journal.pone.015736827310713 PMC 4911051 · doi ↗ · pubmed ↗
- 8Lehmann B.D. Pietenpol J.A. Tan A.R. Triple-Negative Breast Cancer: Molecular Subtypes and New Targets for Therapy Am. Soc. Clin. Oncol. Educ. Book 201535 e 31e 3910.14694/Ed Book_AM.2015.35.e 3125993190 · doi ↗ · pubmed ↗