Efficacy of Deferoxamine Mesylate in Serum and Serum-Free Media: Adult Ventral Root Schwann Cell Survival Following Hydrogen Peroxide-Induced Cell Death
Yee Hang Ethan Ma, Abhinay R. Putta, Cyrus H. H. Chan, Stephen R. Vidman, Paula Monje, Giles W. Plant

TL;DR
This study shows that Schwann cells survive well in serum-free conditions without DFO, but DFO helps in serum-containing conditions by reducing oxidative stress.
Contribution
The study reveals that DFO is unnecessary for Schwann cell survival in serum-free media and highlights the protective role of serum-free conditions.
Findings
Schwann cells survive without DFO pretreatment in serum-free conditions.
DFO increases survival in serum-containing conditions by reducing H2O2-induced cell death.
Serum-free conditions promote a pro-repair state with upregulated autophagy transcripts.
Abstract
Schwann cell (SC) transplantation shows promise in treating spinal cord injury as a pro-regenerative agent to allow host endogenous neurons to bridge over the lesion. However, SC transplants face significant oxidative stress facilitated by ROS in the lesion, leading to poor survival. deferoxamine mesylate (DFO) is a neuroprotective agent shown to reduce H2O2-induced cell death in serum-containing conditions. Here we show that DFO is not necessary to induce neuroprotection under serum-free conditions by cell survival quantification and phenotypic analysis via immunohistochemistry, Hif1α and collagen IV quantification via whole cell corrected total cell fluorescence, and cell death transcript changes via RT-qPCR. Our results indicate survival of SC regardless of DFO pretreatment in serum-free conditions and an increased survival facilitated by DFO in serum-containing conditions.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
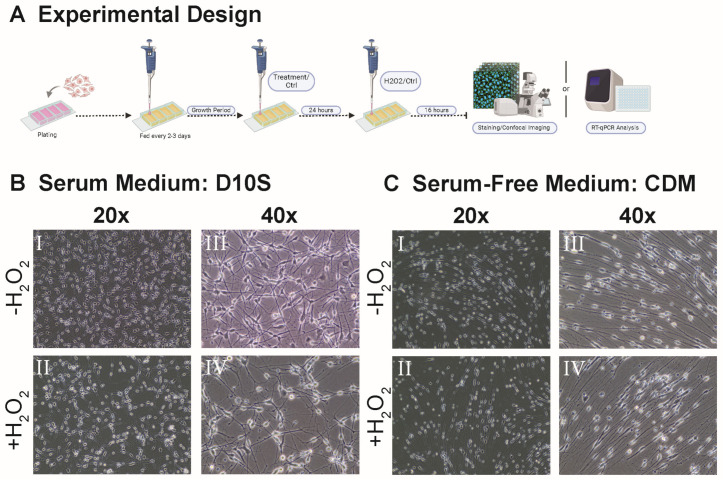 Figure 1
Figure 1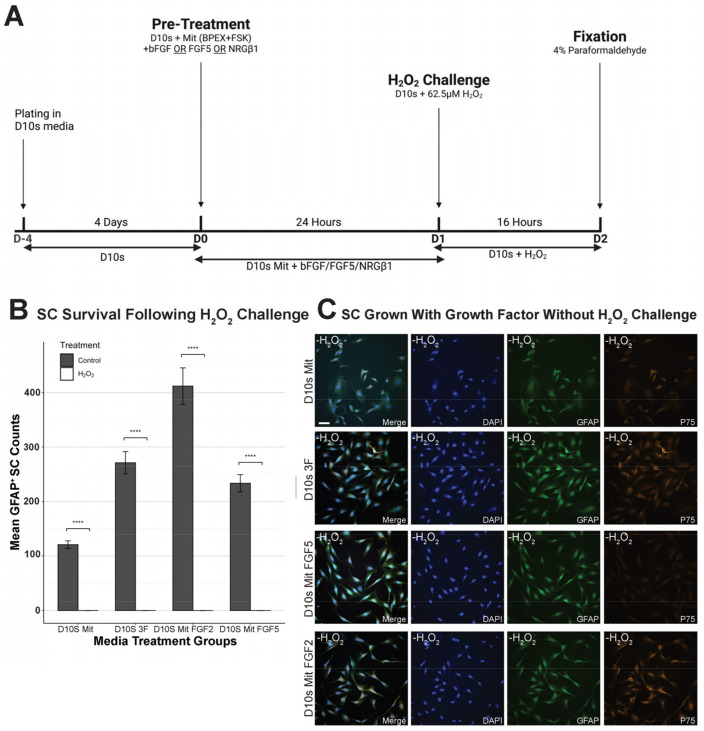 Figure 2
Figure 2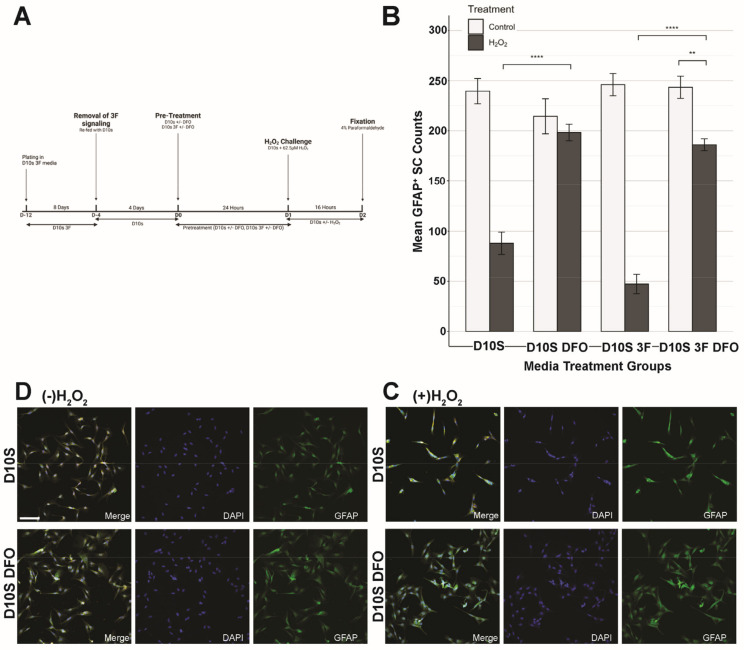 Figure 3
Figure 3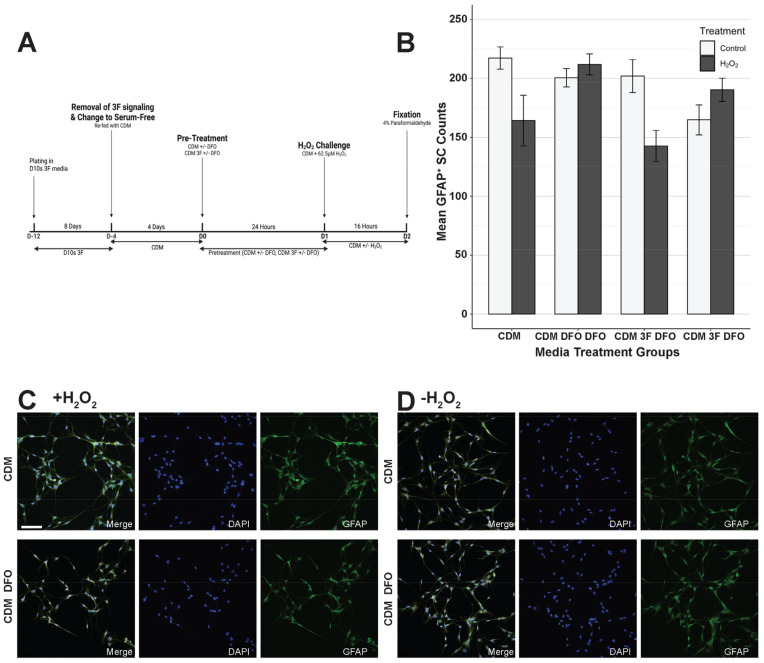 Figure 4
Figure 4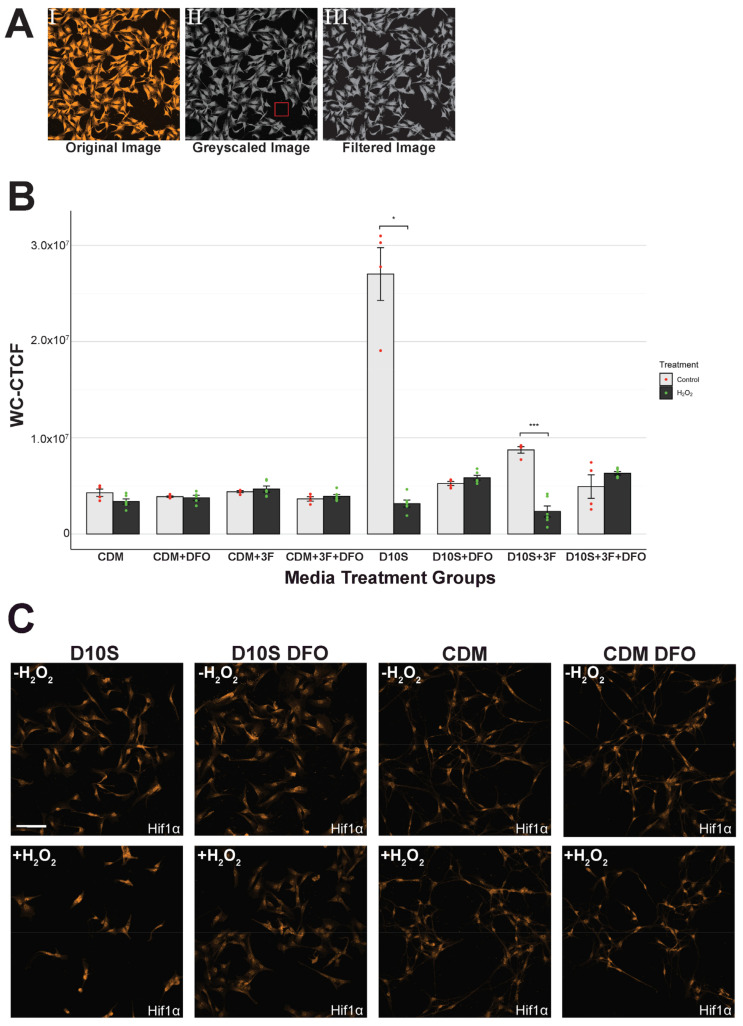 Figure 5
Figure 5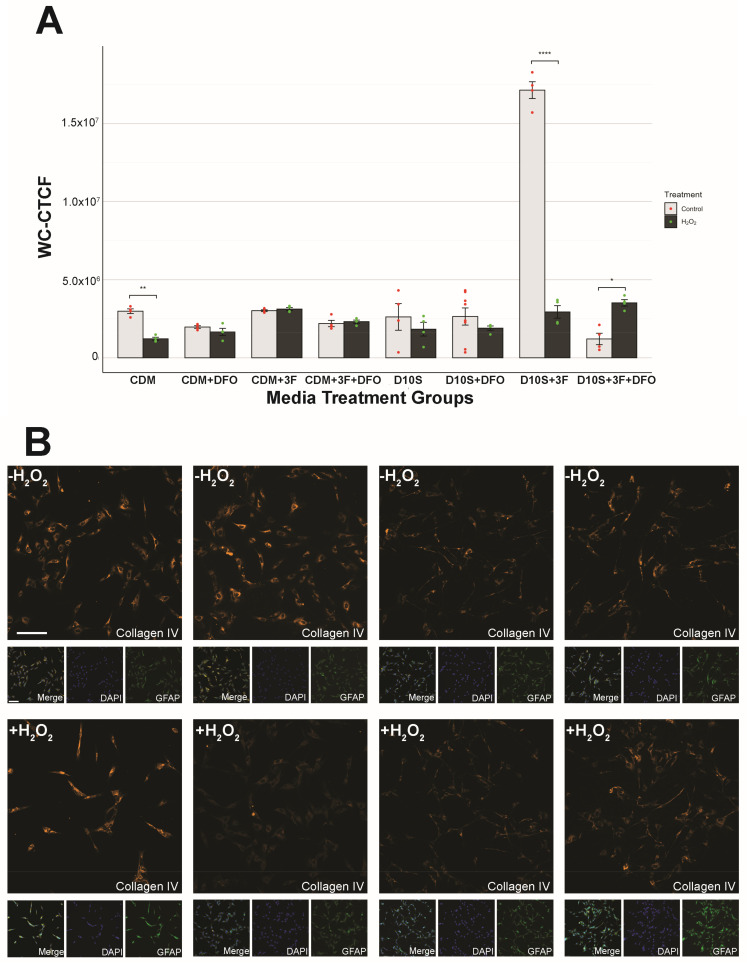 Figure 6
Figure 6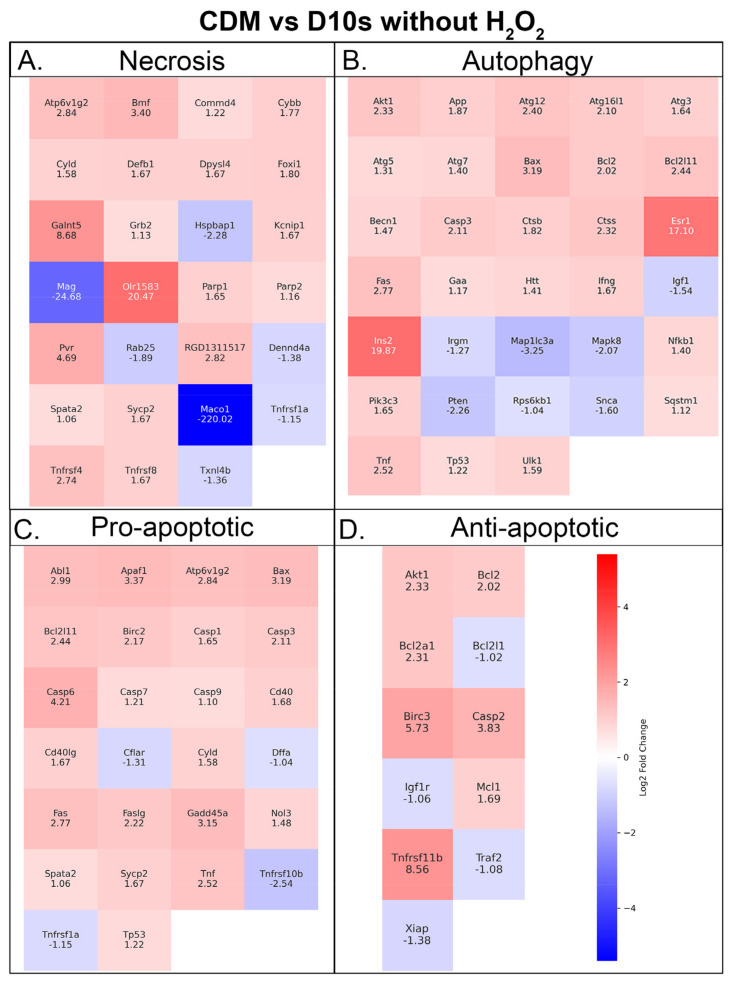 Figure 7
Figure 7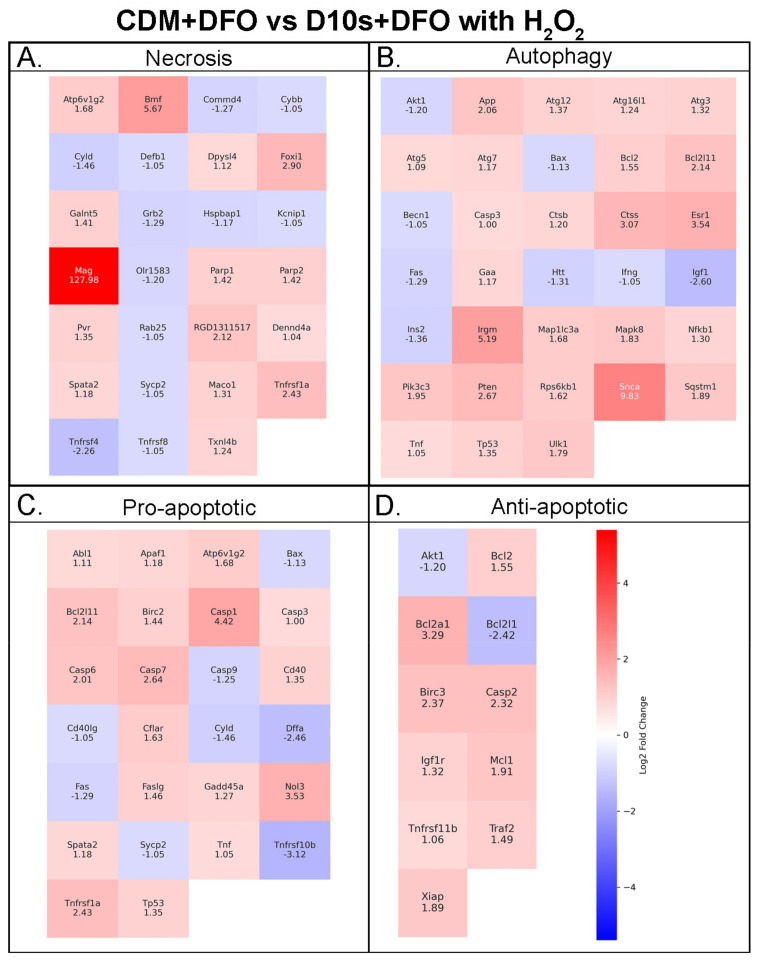 Figure 8
Figure 8- —Ohio State University
- —Ohio State University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNerve injury and regeneration · Spinal Cord Injury Research · Signaling Pathways in Disease
1. Introduction
Spinal cord injury (SCI) is a severe, life-altering condition affecting over 500,000 individuals annually, predominantly resulting from preventable causes such as motor vehicle accidents and violence [1]. A hallmark of SCI is axon degeneration, where the synaptic connections between neurons are disrupted, leading to loss of function and further neurological complications [2,3,4]. Cellular transplantation therapies have emerged as a promising approach for axon regeneration, with autologous Schwann cell (SC) transplants currently being evaluated in clinical trials for their efficacy in treating cervical and thoracic SCI [5,6,7,8,9,10,11,12,13,14]. Originally proposed by David and Aguayo in 1981, peripheral nerve transplants were shown to promote regeneration within the central nervous system by grafting peripheral nerve tissues, which induced axonal regrowth in focal injuries [14]. Since then, SCs have been characterized as peripheral glial cells and have shown considerable regenerative potential in SCI models, with numerous preclinical studies highlighting their capability to facilitate axon regeneration [8,9,10,11,12,13,14]. Compared to other transplant paradigms such as neural progenitor cell (NPC) transplants [15,16,17], SCs offer several advantages. As SCs are mature cells, the risk of spontaneous differentiation into unintended cell types and the formation of ectopic colonies seen in neural stem cell transplantations are eliminated [18]. Furthermore, the autologous nature of the transplants limits the need for immunosuppression following transplantation [11,19]. There are also standardized and straightforward methods to isolate and characterize SCs from a patient’s peripheral nerves, making them an accessible and consistent candidate for cellular transplantation therapy [20,21].
Nonetheless, SC transplants face several challenges, particularly the acute death of transplanted cells caused by oxidative stress in the host spinal cord following injury. Reactive oxygen species (ROS) such as hydrogen peroxide, hydroxyl radicals, and superoxide anion radicals have been shown to induce oxidative stress and mediate cell death in spinal cord injury [22]. Thus, the efficacy of cellular transplant therapies has been limited due to the acute cell death that occurs post-transplantation. To address the issue, recent strategies have been focused on preconditioning SC with various pharmaceutical agents prior to transplantation [23,24,25,26].
One such agent of interest is deferoxamine mesylate (DFO) [22,26]. Mechanistically, DFO acts as an iron (Fe^2+^) chelator, thus inhibiting the function of HIF-PHDs, hypoxia-inducible factor-prolyl hydroxylase domain proteins, and other Fe^2+^, O_2_, and 2-oxogluterate-dependent enzymes that mark HIF, hypoxia-inducible factor, proteins for proteasomal degradation [23,27,28]. This inhibition results in the stabilization of HIFs, allowing them to accumulate and activate their downstream effects. HIF stabilization then increases the expression of hypoxia response element (HRE) genes, which creates a cytoprotective environment and decreases cell death in hypoxic conditions. However, it remains unclear how DFO affects SC protein expression, phenotype, morphology, survivability, and transcriptome in response to ROS-induced cell death. While it is ascertained that DFO provides protection against ferroptosis [29,30], questions remain on whether it is effective in preventing acute cell death mediated by ROS.
The efficacy of DFO as a protective agent against oxidative stress has previously been demonstrated in serum-containing conditions, where it has shown potential to stabilize the expression of hypoxia-inducible factor (HIF) family proteins and protect cells under hypoxic conditions, thereby reducing ROS-induced cell death [23,27,31,32]. However, the relevance of serum-based treatments to modern clinical practices is limited, due to the inherent variability in the composition of serum [33,34,35,36]. A serum-free approach with predefined and consistent composition offers a more reliable alternative, enabling reproducible and clinically relevant results. Hence, it is important to evaluate the effects of DFO on SCs in serum-free conditions to determine whether its protective properties remain effective and clinically relevant.
The goal of the study is to characterize the effectiveness of DFO in increasing the survival of Schwann cells (SCs) upon hydrogen peroxide-induced cell death in an in vitro environment designed to simulate clinical transplant conditions. We hypothesized that SCs pre-treated with DFO would exhibit increased survival in ROS-induced cell death in serum-containing conditions. Additionally, we hypothesized that the neuroprotective effects of DFO would persist in serum-free conditions, which eliminates the need for patient serum extraction while decreasing serum-facilitated SC variation in vivo. We examined the efficacy of DFO in both serum-containing and serum-free conditions (abbreviations and components of medias shown in Supplementary Table S1) and assessed our results through phenotypic staining, cell count, whole cell corrected total cell fluorescence analysis, and RT-qPCR cell death transcriptomics. Our findings show a significant increase in cellular survival upon hydrogen peroxide challenge at 62.5 μM in populations pre-treated with DFO only in serum-containing conditions. Furthermore, we observed that serum-free media exhibited inherent neuroprotective properties, presenting it as a viable alternative to serum-containing media altogether.
2. Materials and Methods
2.1. Generation and Expansion of Schwann Cell Culture
Schwann cells (SCs) were isolated from the ventral roots of 6-month-old adult GFP transgenic Sprague Dawley rats according to our established protocols for SC culturing from immediately dissociated nerve tissues [37]. The initial cell harvest was plated in the form of droplets directly onto PLL-laminin-coated dishes and expanded in DMEM medium supplemented with 10% FBS, heregulin (10 nM), and forskolin (2 μM) up until confluency. Cells were subsequently lifted from their dishes by trypsinization and purified of contaminating fibroblasts by MACS sorting after incubation with Thy1.1 antibodies, as described in Ravelo et al. [37]. The purified SCs were expanded up to passage-1 (P1) in the abovementioned medium for the creation of cryogenic stocks [37]. These stocks were maintained in liquid nitrogen until use. Prior to the experiment, P1 SCs were expanded in D10S 3F (Supplementary Table S1). During expansion, SCs were cultured in poly-L-lysine (PLL, Millipore Sigma P6282, St. Louis, MO, USA) coated 100 mm Corning-treated tissue culture dishes and fed with 7 mL of CDM 3F (Supplementary Table S1) every 2–3 days. Once the desired confluency (approx. 5 million cells per plate) and maturation state were achieved by visual confirmation of cell swirling, SCs were either split into new 100 mm Corning-treated tissue culture dishes or split into experimental conditions. SCs were detached using 0.05% trypsin-EDTA (Gibco 25300054, Waltham, MA, USA) and seeded into PLL-coated four-well chamber slides (Nunc Lab-Tek II Chamber Slide System, ThermoFisher, Waltham, MA, USA), at 50,000 cells per well, for immunocytochemistry experiments and fed with 350 μL CDM 3F per well every 2–3 days until the start of the experiment (Section 2.3). For RT-qPCR experiments, SCs were plated into 60 mm PLL-coated TPP tissue culture dishes (TPP 93060, Trasadingen, Switzerland) at 450,000 cells/plate and fed with 5 mL of CDM 3F every 2–3 days until treatment (Section 2.4). All SCs were cultured in a 5% CO_2_, 37 °C incubator except for the cells at passage-zero (P0) and P1, which were cultured in a CO_2_ incubator set up at 9% CO_2_. P2-P5 SCs were used for all experiments. The purity of the SC populations was >98% at P1, as judged by co-immunostaining with S100B and Thy1.1 antibodies. SCs from ventral roots were highly proliferative and visually indistinguishable from the ones derived from the sciatic nerve [38].
2.2. Experimental Treatments and Timeline
SCs were plated onto four-well chamber slides for phenotypic staining and subsequently onto 60 mm PLL-coated TPP tissue culture dishes for cell death transcriptomics analysis. All SCs are initially cultured in serum containing CDM 3F media until the desired confluency is reached by visual confirmation of swirling. Prior to the 62.5 μM H_2_O_2_ challenge or sham, SCs were treated with their respective pre-treatment conditions (Supplementary Table S2) for 24 h, followed by a 16-h exposure to 62.5 μM H_2_O_2_ in base media. 3F signaling was cleared for selected groups (Section 3.3 and Section 3.4 comparing SCs serum and serum-free. SCs were fed with their respective base medium for 4 days;. For sham, SCs were refed with base media only. After the treatment period, SCs were either fixed with 4% paraformaldehyde for subsequent phenotypic staining and density analysis or harvested as cell pellets for RT-qPCR cell death transcriptomics analysis (Section 2.4). Figure 1A illustrates the experimental timeline, and Figure 1B,C shows the effect of the 16-h exposure to 62.5 μM H_2_O_2_ on SCs. Figure 2A, Figure 3A, Figure 4A and Figure 7A show the timeline of the media condition changes during the experiments.
2.3. Immunofluorescence Staining
16 h after the H_2_O_2_ challenge, SCs were fixed via a 15 min incubation with approximately 300 μL of chilled 4% paraformaldehyde at room temperature. After PFA incubation, SCs were washed three times with 1X PB and stored at +4 °C with phosphate buffer (PB) until immunostaining. We stained SCs with GFAP (Chicken, 1:1000, Aves GFAP, Davis, CA, USA), P75 (Rabbit, 1:400, Bioss BS-0161R, Woburn, MA, USA), HiF1α (Rabbit, 1:500, Sigma Aldrich SAB5701087, St. Louis, MO, USA), and Collagen IV (Rabbit, 1:400, Rockland 600-401-106-0.1, Pottstown, PA, USA) primary antibodies. Secondary antibodies used were from Jackson Immuno (West Grove, PA, USA) and used at 1:800; we used donkey anti-chicken AlexaFluor 488 (703-546-155) and donkey anti-rabbit cy3 (711-166-152). SCs were incubated at room temperature with primary antibody in blocking solution: 90% 1X phosphate buffer (PB), 10% Donkey Serum (Lampire 7332100, Pipersville, PA, USA), and 0.2% Triton X-100 (Sigma Aldrich X100, St. Louis, MO, USA), for 2 h, then washed with 1X PB three times. SCs are then incubated with secondary antibody in blocking solution at room temperature for 45 min in the dark. Cells are subsequently washed with 1X PB three times and cover slipped with Prolong Gold or Diamond Antifade Mountant with DAPI (Invitrogen, Waltham, MA, USA).
2.4. Confocal Microscopy
All imaging was performed on a Nikon AXR confocal microscope using the Nikon NIS Elements software. The microscope is equipped with four lasers, ranging from 405 to 647. All cells were stained with DAPI (405) and their respective stains for qualitative and quantitative analysis (GFAP, P75, HIF1α, Collagen IV). Image acquisition parameters are described in the sections below in the context of their application. Representative images taken for Figure 2 were taken using a Nikon APO LWD 40Xx WI lS DIC N2 (NA = 1.15, WD = 610 μm) objective. Representative images for Figure 3, Figure 4, Figure 5 and Figure 6 were taken using a Nikon APO LWD 20x WI lS (NA = 0.95, WD = 950 μm).
2.5. Survival Quantification
Images taken from the wells were quantified using the NIH ImageJ software (ImageJ 2.14.0/1.54f; Java 1.8.0_322 [64 bit]). Quantification was semi-automated with human correction of software errors. Images were taken at 10× (NA = 0.45, WD = 4000 μm) magnification for each well. For each well, we randomly selected 3–4 fields. Using ImageJ, ND2 images were opened as grayscale images. The detection threshold was manually adjusted for all images to accurately highlight each individual DAPI stain. Images were then converted to binary images to remove background noise before being processed with the watershed algorithm to split any combined nucleus. DAPI nuclei were subsequently quantified using particle analysis with the size detection threshold set to 100 pixels to infinity. Quantification results were then compared with the original DAPI and GFAP stain manually to ensure accurate quantification and address any potential errors (e.g., splitting of a single nucleus, grouping of multiple nuclei, counting GFAP negative nuclei). Final quantifications were manually corrected for any software errors.
2.6. Hif1α and Collagen IV Quantification
Immunofluorescence images were obtained via confocal microscopy described in Section 2.5. Images were taken using a 20× objective (NA = 0.75, WD = 1000 μm), maintaining identical parameters for all Hif1α and Collagen IV scans as follows: laser power: 33, gain: 84.2, pinhole size: 1.2, zoom: 1×, z-stack: 50 μm, and step size: 2 μm. For Hif1α, we selected n = 6 per pre-treatment group (Supplementary Table S2). For collagen IV, we selected n = 4 per pre-treatment group. The images were then exported as ND2 files and further processed using a custom, semi-automated pipeline to quantify whole cell corrected total cell fluorescence (WC-CTCF) using FIJI/ImageJ software. In brief, ND2 files were split into single channels and compressed to maximum intensity projections (MIPs) for quantification. Then, the MIPs for Hif1α and Collagen IV were measured for total fluorescence in the image (integrated density), subtracted by the total background fluorescence. Utilizing this approach permits correction for size, density, or morphological differences between cells in their respective treatment groups. The formula used was CTCF = Integrated density − (total area × background intensity mean). We analyzed the whole cell fluorescence level to capture cytoplasmic and nuclear expression levels.
2.7. Statistical Analysis
All statistical analyses were conducted using the RStudio software 2024.04.2+764. Data were presented with error bars indicating standard error of the mean (SEM). Depending on data characteristics, either Welch’s one-way analysis of variance (Welch’s ANOVA) or standard ANOVA was used for F-tests. Post hoc comparisons were performed using Tukey’s HSD, Dunn’s Test, or Games–Howell test, as appropriate. p ≤ 0.05 denotes statistical significance. Significance was also presented with the following notations: * (p ≤ 0.05), ** (p ≤ 0.01), *** (p ≤ 0.001), **** (p ≤ 0.0001). For RT-qPCR analysis, fold change was reported on the log2 scale, and comparative analyses were performed on GeneGlobe (Qiagen, Hilden, Germany).
2.8. Cell Death Transcriptomics
2.8.1. RNA Isolation
Selected groups (Supplementary Table S3) were chosen for transcriptomic analysis. Samples were lysed using the Qiashredder (Qiagen 79656, Hilden, Germany). RNA was isolated using the RNeasy Micro Kit (Qiagen 74004, Hilden, Germany) following the manufacturer’s instructions. Isolated RNA was subsequently stored at –80 °C until use. The quantity of extracted RNA was evaluated using Nanodrop 2000 (ThermoFisher, Waltham, MA, USA) or BioTek Synergy HTX Multimode Reader (Agilent, Santa Clara, CA, USA), while the integrity was evaluated using Bioanalyzer 2100 (Agilent) for RNA Nano Chips (Agilent, Santa Clara, CA, USA) and TapeStation 4200 (Agilent, Santa Clara, CA, USA) for RNA ScreenTapes (Agilent, Santa Clara, CA, USA). RNA integrities were ensured to be of sufficient quality prior to PCR. 100 ng of total RNA was utilized for subsequent analysis within each group.
2.8.2. cDNA Synthesis
cDNA was synthesized using the RT2 First Strand Kit (Qiagen, 330404, Hilden, Germany) following the manufacturer’s instructions. Samples were processed using the T100 Thermo-Cycler (BioRad, Hercules, CA, USA), starting with a 15-min incubation at 42 °C followed by a 5 min denaturation at 95 °C. Synthesized cDNA was stored at –20 °C until use. The quality and quantity of synthesized strands were evaluated using a Nanodrop 2000 (ThermoFisher, Waltham, MA, USA).
2.8.3. qPCR
The 96-well RT2 Profiler PCR Array Rat Cell Death PathwayFinder (Qiagen 330231 PARN-212ZA, Hilden, Germany) was used to detect expression levels of 84 targeting genes and 5 control genes (Supplementary Table S4). Selected genes were essential for the central mechanisms of cellular death, spanning apoptosis, autophagy, and necrosis. Following the manufacturer’s instructions, cDNA was mixed with RT2 SYBR Green qPCR Mastermix (Qiagen 330502, Hilden, Germany) before loading onto the sample plate. The sample plate was processed using the QuantStudio3 System (Applied Biosystems, Inc., Waltham, MA, USA) starting with a 10-min incubation at 95 °C, followed by 40 cycles of denaturation at 95 °C for 15 s and annealing/extension at 60 °C for 30 s. Geneglobe (Qiagen, Hilden, Germany) was used for comparison studies, fold change regulation, and heatmap generation. The detection threshold was standardized to 20,000. The CT threshold is set to 35, and control genes were normalized using geometric means.
2.8.4. Gene Ontology Analysis
A gene ontology (GO) analysis was performed based on the heatmaps generated from RT-qPCR data. Differentially expressed genes were quantified using log2-fold change and reported as up- or downregulated, relative to a CDM control. Groups compared CDM vs. CDM, both without H_2_O_2_, and CDM vs. D10S, both with H_2_O_2_ and DFO treatment. GO enrichment analysis was performed using the PANTHER Classification System (v19.0, https://pantherdb.org/ (accessed on 17 December 2024)). The individual genes were entered into the database, specifying Rattus norvegicus as the reference species. Functional annotation categories included biological process, molecular function, cellular component, and protein classification. Due to biological replicates of n = 1 per condition, statistical testing for the RT-qPCR results was not conducted.
3. Results
3.1. Variation in SC Morphology Between Serum-Containing and Serum-Free Conditions
Schwann cell morphology varied between serum-containing and serum-free conditions, with distinct differences observed in soma and cellular processes. In serum-containing conditions, SCs exhibited a more expanded soma, which appeared flatter and covered a larger surface area (Figure 1B). In contrast, SCs cultured in serum-free conditions displayed a more compact, needle-like bipolar soma characterized by a narrower and more defined structure (Figure 1C). Similarly, differences were observed in the morphology of SC processes. In serum-containing conditions, SC processes were comparatively shorter and remained proximal to the soma with limited extension (Figure 1B). In contrast, SCs in serum-free conditions displayed more extended, fiber-like processes that stretched distally and formed bipolar processes from the soma, indicative of a more differentiated state (Figure 1C). SCs in serum-containing and serum-free conditions also responded differently in the event of H_2_O_2_-induced oxidative stress. In serum-containing conditions, SC morphology and density were greatly altered in the presence of H_2_O_2_-challenge (Figure 1B), whereas a more consistent SC phenotype was observed in serum-free conditions (Figure 1C).
3.2. Growth Factors Failed to Promote SC Survival Following H2O2 Exposure
We investigated the potential of growth factors to enhance cellular survival following H_2_O_2_-induced oxidative stress. By following the experimental paradigm (Figure 2A), SCs were pre-treated with either fibroblast growth factor 2 (FGF2), fibroblast growth factor 5 (FGF5), or Neuregulin Beta1 (NRG)—in D10S Mit media (Supplementary Table S2) for 24 h prior to H_2_O_2_ exposure. Following their pretreatments, SCs were analyzed by immunofluorescence staining to evaluate DAPI, p75, and GFAP expression (Figure 2C). SC survival was evaluated by quantifying GFAP-positive SC nuclei, followed by statistical analysis. Comparing growth factors pretreated SCs without H_2_O_2_ control, D10S Mit, shows significantly increased proliferation (Figure 2B; D10S + Mit vs. D10S + Mit + FGF2, p = 7.26 × 10^−22^, ****; D10S + Mit vs. D10S + Mit + FGF5, p = 3.19 × 10^−4^, ***; D10S + Mit vs. D10S + 3F, p = 4.76 × 10^−7^, ****), validating their role in promoting proliferation of SCs. However, SCs were not protected against H_2_O_2_-induced cell death when pre-treated with growth factors, as differences in GFAP-positive SC nuclei across treatments were statistically insignificant (Figure 2B), indicating that pretreatment with growth factors is not sufficient to protect SCs against reactive oxygen species-induced cell death (D10S + Mit control vs. D10S + Mit + H_2_O_2_, p = 1.51 × 10^−2^, *; D10S + Mit + FGF2 control vs. D10S + Mit + FGF2 + H_2_O_2_, p = 7.26 × 10^−22^, ****; D10S + Mit + FGF5 control vs. D10S + Mit + FGF5 + H_2_O_2_, p = 1.63 × 10^−9^, ****; D10S + 3F control vs. D10S + 3F + H_2_O_2_, p = 1.09 × 10^−10^, ****). Morphological observations from confocal images of SCs grown in selected growth factors (Figure 2C) confirmed that growth factors promoted the typical elongated morphology of in vitro cultured SCs.
3.3. Deferoxamine Mesylate (DFO) Increased SC Survival Following H2O2 Challenge in Serum-Containing Conditions
Deferoxamine mesylate (DFO) is an FDA-approved iron chelator previously suggested to enhance Schwann cell survival in hypoxic conditions [22]. We tested if DFO could effectively prevent H_2_O_2_-induced SC death in serum-containing conditions (D10S). We also tested whether the addition of 3F during DFO treatment would influence DFO’s protective effect against H_2_O_2_-induced oxidative stress. We hypothesized that 3F would prime SCs for growth and proliferation, which could, in turn, impair their ability to respond to H_2_O_2_ induced cell death. Following their treatment protocols (Figure 3A and Figure 4A), SC survival was evaluated by quantifying GFAP-positive SC nuclei, followed by statistical analysis via Tukey’s HSD test. Results showed that, under H_2_O_2_-induced cell death, SC did not survive in D10S (Figure 3B; D10S + H_2_O_2_ vs. D10S -H_2_O_2_, p = 1.15 × 10^−13^, ****). Additionally, inclusion of 3F did not lead to any significant improvement in SC survival (Figure 3B; D10S + 3F + H_2_O_2_ vs. D10S + 3F control, p = 3.73 × 10^−14^, ****), consistent with our previous findings that 3F does not provide any protective effect against oxidative stress (Figure 2B; D10S 3F +H_2_O_2_ vs. D10S 3F control, p = 1.09 × 10^−10^, ****). We added DFO to evaluate its effects on SC survival and phenotype in the presence of H_2_O_2_-induced cell death. The inclusion of DFO in both D10S and D10S 3F medium protected SCs and increased survival following exposure to H_2_O_2_ (Figure 3B; D10S + H_2_O_2_ vs. D10S + DFO + H_2_O_2_, p = 2.39 × 10^−10^, ****; D10S 3F + H_2_O_2_ vs. D10S 3F + DFO +H_2_O_2_, p = 1.18 × 10^−13^, ****). Furthermore, comparisons between D10S DFO + H_2_O_2_ and D10S -H_2_O_2_ showed no significant difference in SC survival, indicating that DFO reduced H_2_O_2_-induced cell death and restored SC survival to baseline levels. However, in D10S 3F medium, DFO pre-treatment (D10S 3F + DFO +H_2_O_2_) did not protect SC survival to baseline levels (D10S -H_2_O_2_). This suggests that while 3F is essential in promoting SC proliferation, it can interfere with the effects of DFO and reduce its protectiveness. Nonetheless, these results showed that DFO conferred significant protection against H_2_O_2_-induced cell death in serum-containing conditions. Immunofluorescent staining indicated that H_2_O_2_ challenge resulted in altered SC morphology and non-uniform SC distribution in serum-containing conditions (Figure 3C).
3.4. Serum-Free Media Protected SCs Against H2O2-Induced Cell Death Without DFO
We also investigated if DFO could effectively prevent H_2_O_2_-induced cell death in serum-free medium: CDM, which is a DMEM/F12 medium supplemented with bovine insulin, human transferrin, sodium selenite, and putrescine (Supplementary Table S1). The experimental protocol and quantification methods outlined in Section 3.3 were repeated with the only modification being the substitution of serum-containing D10S with serum-free CDM as the base medium (Figure 4A). Interestingly, CDM alone was sufficient to promote SC survival under H_2_O_2_-induced cell death (Figure 4B, CDM -H_2_O_2_ vs. CDM +H_2_O_2_, p = 1.157 × 10^−13^, ns). Even without the addition of DFO, CDM alone restored SC survival to baseline levels, suggesting that factors in CDM may contribute to preventing H_2_O_2_-induced cell death. Here, the inclusion of 3F did not decrease the protection provided by CDM against H_2_O_2_ induced cell death (Figure 4B, CDM 3F -H_2_O_2_ vs. CDM 3F +H_2_O_2_, p = 5.037 × 10^−2^, ns), indicating that 3F is not sufficient to interfere with the effects of CDM and reduce its protectiveness, unlike their serum counterpart. The inclusion of DFO in serum-free CDM did not affect SC survival in H_2_O_2_-induced cell death, indicating that DFO is unnecessary in serum-free CDM to promote SC survival (Figure 4B, CDM DFO -H_2_O_2_ vs. CDM DFO +H_2_O_2_, p = 9.989 × 10^−1^, ns). Immunofluorescent staining showed SC distribution and morphology was mostly unaffected after H_2_O_2_-challenge in serum-free conditions (Figure 4C).
3.5. Hif1α Expression Consistent in Serum-Free Conditions but Inconsistent in Serum Conditions
It is previously reported that DFO acts as a neuroprotective agent through the upregulation of the hypoxia-inducible factor (HIF) protein family to promote cellular survival in hypoxic environments such as the ROS-rich environment in our study. We tested if Hif1α will be upregulated here with our SCs. To this end, we have stained for hypoxia-inducible factor 1 alpha (Hif1α) and subsequently quantified the protein expression using whole cell corrected total cell fluorescence (WC-CTCF). Statistical analysis of the Hif1α WC-CTCF reveals a consistent Hif1α expression across serum-free groups regardless of DFO pretreatment or H_2_O_2_ challenge (Figure 5B). Hif1α expression in serum shows variable expression with a significantly high expression of Hif1α in D10S (Figure 5B, D10S -H_2_O_2_ vs. D10S +H_2_O_2_, p = 1.800 × 10^−2^, *) and D10S 3F (Figure 5B, p = 2.300 × 10^−4^, ***) without H_2_O_2_ challenge (Figure 5B). Qualitative analysis on immunocytochemistry images reveals localized nuclear expression of Hif1α in serum-free conditions and higher levels of cytoplasmic Hif1α expression in non-DFO pretreated and non-H_2_O_2_ challenged SCs in serum conditions (Figure 5C).
3.6. Serum Drives High Variability in Collagen IV Expression
The secretion profile of the extracellular matrix (ECM) by SCs plays a critical role in SC adhesion, maturation, and subsequent nerve regeneration following transplantation. Collagen IV is one of the key ECM elements expressed by SCs involved in the regeneration of myelin sheath, an indicator of SC maturation [35]. Wei investigated whether the addition of DFO would influence the collagen IV expression profile of SCs under both serum and serum-free conditions, with and without H_2_O_2_-induced cell death. Collagen IV expression was analyzed through immunofluorescence staining, followed by WC-CTCF analysis to quantify collagen IV expression (Section 2.7). WC-CTCF analyses revealed varying patterns of collagen IV expression between SCs in serum-containing and serum-free medium. Collagen IV expression was generally consistent across SCs cultured in CDM. The addition of DFO did not result in any measurable changes in collagen IV expression in the majority of CDM groups when compared to their non-DFO treated counterparts (statistics shown in Supplementary Table S6), suggesting that DFO did not alter the ECM expression of collagen IV in serum-free conditions. Surprisingly, when SCs are challenged with H_2_O_2_ in CDM medium (CDM +H_2_O_2_), collagen IV expression shows a significant drop compared to control (CDM -H_2_O_2), indicating that DFO or 3F may be needed in serum-free conditions to promote the production of extracellular collagen IV in SCs in the presence of ROS. In serum conditions, collagen IV expression is highly variable. When pretreating SCs with 3F, collagen IV expression is highly upregulated without the presence of ROS (Figure 6; D10S 3F -H_2_O_2 vs. D10S 3F +H_2_O_2_, p = 1.000 × 10^−5^, ****). However, when pretreatment includes DFO, the upregulation of collagen IV is diminished. Overall, SCs in serum conditions exhibit largely variable collagen IV expression compared to serum-free conditions.
3.7. Serum vs. Serum-Free Conditions Show Differential Transcriptional Responses in Cell Death Pathways
We asked if treatment with DFO leads to transcriptional changes in cell death pathways, specifically apoptosis, necrosis, and autophagy pathways. To this end, we have isolated RNA from SCs in varying conditions (Supplementary Table S8) for use in RT-qPCR to investigate transcriptional changes in cell death pathways. The fold changes of genes were measured as upregulation or downregulation in serum-free (CDM) groups relative to serum-containing (D10S) groups. The largest fold changes when comparing CDM vs. D10S without H_2_O_2_ were observed in genes associated with necrosis, specifically downregulation of Maco1 (−220.02) and Mag (−24.68). The largest fold change observed in this group was for Olr1583 (20.47), which plays a key role in necrosis. In comparison, DFO and H_2_O_2_-treated groups (CDM + DFO +H_2_O_2_ vs. D10S + DFO + H_2_O_2_) showed the greatest fold changes in both anti-apoptotic genes and necrosis genes. The largest upregulation was seen in genes associated with necrosis, including Defb1 (21.09) and Hspbap1 (11.68). In contrast, the top downregulated genes were Tnfrsf11b (−17.64) and Bcl2L11 (−8.08), both of which are associated with anti-apoptotic pathways. Considering the substantial reduction of Maco1 transcription when comparing CDM vs. D10S, genes associated with autophagy, Ins2 (19.87) and Esr1 (17.1), showed the next largest increase in transcription. Overall, the data suggest CDM primarily reduces genes involved in necrosis while increasing genes associated with autophagy in the absence of H_2_O_2_. When both groups are treated with DFO and H_2_O_2_, genes involved in necrosis are upregulated, while anti-apoptotic transcripts are suppressed. Interestingly, fold changes for autophagy genes appear largely unchanged. Heatmaps of RT-qPCR are shown in Figure 7 and Figure 8.
4. Discussion
We examined the effect of deferoxamine mesylate (DFO) on the survival of ventral root Schwann cells isolated from adult Sprague Dawley rats in H_2_O_2_-induced cell death. It was previously reported that DFO increases SC survival in H_2_O_2_-induced cell death and hypoxic conditions in vitro in serum-containing conditions [23,39]. Here, we expanded the horizon to study DFO’s effectiveness in protecting SCs from H_2_O_2_-induced cell death in more clinically relevant, serum-free conditions and presented our results through phenotypic staining, SC survival quantification, whole cell corrected total cell fluorescence (WC-CTCF) analysis, and RT-qPCR cell death transcriptomics analysis.
4.1. Distinctive SC Morphology in Serum and Serum-Free Conditions
Consistent with previous findings from Plant and colleagues [30,39], SCs exhibited drastically different morphology between serum-containing and serum-free conditions (Figure 1). The differential morphology of the two culture conditions affects SC maturation and regenerative capacity in vivo. For instance, the bipolar and spindle morphology of SCs in CDM shows a pro-regenerative phenotype in comparison to the small and flat phenotype of SCs in D10S. The spindle morphology allows for neurites to grow on top, acting as a bridging mechanism and allowing for nerve regeneration. However, SCs in serum conditions switch from a regenerative state into a proliferative state and abandon cell contact [40]. Thus, SCs grown in serum appear to be more prone to H_2_O_2_-induced cell death due to the energetic switch to a proliferative state. Furthermore, serum has been reported to lead to unstable and variable phenotypes in vitro in mesenchymal stromal cells and olfactory ensheathing glia [41,42,43]. Although the literature lacks work describing SCs in serum versus serum-free medium, the use of serum would still largely decrease the reproductive capacity of SC cultures in transplantation studies due to the lot-to-lot variability in serum production.
4.2. Growth Factors Provide No Protection Against H2O2-Induced Cell Death
We asked if growth factors would be able to attenuate H_2_O_2_-induced cell death in Schwann cells (SCs). We pre-treated SCs with fibroblast growth factor 2 (FGF2), or fibroblast growth factor 5 (FGF5), or neuregulin beta 1 (NRGb1), for 24 h prior to 16 h of 62.5 mM H_2_O_2_ challenge. Our results indicate no survival of SCs following H_2_O_2_ challenge regardless of pretreatment. Although it has been previously reported that these factors attenuate H_2_O_2_-induced cell death in alternative cell types such as embryonic cortical neurons [44] and bone marrow mesenchymal stem cells [45], we have shown that they do not attenuate H_2_O_2_ cell death in adult SCs. Here, we did not test these growth factors independently in serum-free conditions. In serum-free conditions, growth factors could produce a stronger protectiveness in addition to the serum-free protectiveness through the promotion of proliferation. However, as seen with SCs grown in 3F, it may also drive SCs into growth states and thus reduce their capacity to withstand H_2_O_2_-induced cell death.
4.3. DFO Efficacy in Serum and Serum-Free Conditions
Here, we have shown that serum-free CDM medium seems to protect SCs from H_2_O_2_-induced cell death regardless of DFO pre-treatment. In serum conditions, Hill and colleagues [22,26] have previously established DFO effectiveness in vitro, showing its ability to upregulate Hif1α and increase cellular survival by means of live–dead staining [22]. However, we have shown here that the use of CDM without DFO pretreatment was able to achieve similar levels of protection against H_2_O_2_ cell death to DFO in serum-containing D10S medium. The use of CDM is also shown to be a better alternative due to its consistency that is largely unachievable in serum-based treatments due to high variation in serum, which contains variable and trace amounts of growth factors such as BDNF, FGF5, FGF2, and NRG beta 1, which vary from batch to batch in serum production [46]. While DFO has been proven to increase SC survival in serum-containing conditions [22], most of its effects are masked by the inherent protectiveness provided in CDM, suggesting that the inclusion of DFO in CDM may not be necessary to achieve H_2_O_2_-induced cell death protection.
4.4. Transferrin in CDM Medium May Influence HiF-PhD to Override DFO Effects in SCs
The combined factors of transferrin, insulin, and sodium selenite, also known as ITS serum-free supplement, are a long-established serum-free alternative to the use of fetal bovine serum supplement in tissue culture medium. These factors are critical in supporting cellular proliferation, survival, and glucose uptake. Specifically, transferrin acts as an iron transporter and has been reported to decrease iron accumulation in Parkinson’s disease-affected neurons [47]. Mechanistically, transferrin receptor (TfR) acts downstream of Hif1α expression, allowing cells to uptake higher levels of iron during low oxygen environments for increased mitochondrial activity [48,49]. With the addition of transferrin in the serum-free medium, the process of Hif1α modulated upregulation of TfR may be overwritten by the presence of transferrin to sufficiently provide iron to SCs and may shut down DFO-facilitated HiF-PhD inhibition with increased iron levels, leading to our observed levels of SC Hif1α expression in serum-free conditions (Figure 5C). It may be of interest to pursue the individual effects of each supplement component in H_2_O_2_-induced SC death to further characterize the protection offered in CDM.
4.5. Nuclear Expression of Hif1α Facilitates Protection in Serum-Free Cultured or DFO Pre-Treated SCs
We have quantitated whole cell Hif1α expression via WC-CTCF. Here, we see a consistent nuclear expression of Hif1α in SCs grown in serum-free conditions (Figure 5B,C) but with an inconsistent expression of Hif1α in SCs grown in serum conditions (Figure 5B). Furthermore, ICC reveals a cytoplasmic expression of Hif1α in non-H_2_O_2_ challenged SCs in serum conditions (D10S -H_2_O_2_ and D10S 3F -H_2_O_2_, Figure 5C). Previous work has reported the expression of Hif1α enhances SC survival when transplanted into the spinal cord [47]. Stabilization and subsequent nuclear translocation of Hif1α allows for binding with hypoxia-response element (HRE) genes and induces protection against oxidative stress through the upregulation of critical angiogenic and protective genes such as vascular endothelial growth factor, or VEGF [50,51,52], which has previously been reported to be upregulated through western blot and qPCR analysis [22]. Here, our data add strong evidence for this protection such that the DFO pre-treated SCs survived compared to non-DFO pretreated SCs in serum conditions. However, in serum-free conditions, this protectiveness seems to be mediated by an alternative mechanism since, regardless of DFO pretreatment, SCs survived the H_2_O_2_ challenge. Further, the nuclear expression of Hif1α seems to be present in SCs in serum-free conditions but further enhanced by the presence of DFO. It may be crucial to determine the mechanism of this protection to understand the protection of SCs against H_2_O_2_-induced cell death in serum-free conditions. It would be of interest to explore the differences of Hif1α presence in the cytoplasm and nucleus in serum versus serum-free conditions as well.
4.6. Collagen IV Expression Reveals Proliferative SCs in Serum Media and Pro-Regenerative SCs in Serum-Free Media
We quantitated whole cell fluorescence of collagen IV in SCs to determine if there are any extracellular matrix changes. Collagen IV is an important factor in determining states of SCs; it serves as an indicator of the proliferation and remyelination potential of SCs. We have shown here that by inducing proliferative states in SCs via the addition of the 3F (forskolin, bovine pituitary extract, and neuregulin) in serum-containing medium, survival in H_2_O_2_-induced cell death is negatively affected such that survival is decreased compared to SCs without proliferative 3F signaling in serum conditions (Figure 4B, D10S 3F -H_2_O_2_ vs. D10S 3F +H_2_O_2_, p = 3.730 × 10^−14^). Subsequent collagen IV quantitation also reveals a higher proliferative state in the D10S + 3F medium condition through a high collagen IV expression when not challenged with H_2_O_2_, where the proliferative state is shut down following the H_2_O_2_ challenge (Figure 6, D10S 3F -H_2_O_2_ vs. D10S 3F +H_2_O_2_, ****, p = 1.000 × 10^−5^). Previous work has shown immature or proliferative Schwann cells do not facilitate regeneration as efficiently as matured Schwann cells [26]; our results thus indicate that the more mature states of Schwann cells in serum-free media will facilitate higher regeneration as a cellular transplant, owing to the more stabilized extracellular matrix (Figure 6) and their bipolar process phenotype, which allows for host neurons to bridge the lesion gap (Figure 2 and Figure 4).
4.7. Serum-Free, Chemically Defined Media Shifts SCs Towards Heightened Metabolism and Cellular Recycling Phenotype Without the Presence of H2O2 and DFO
Here, we identified numerous differentially expressed genes in our in vitro model of hydrogen peroxide-induced cell death. Transcriptomic changes associated with necrosis, autophagy, and apoptosis were observed, and the GO enrichment analysis has provided key insights into how these genes affect cell death pathways. In the absence of DFO or H_2_O_2_, the serum-free group, CDM-H_2_O_2_, primarily showed changes in genes of the necrotic pathway, especially downregulation of Maco1 and Mag. In addition to the necrosis pathway, these genes play a role in cytoskeletal dynamics, cell adhesion, myelination, and metabolic shifts in response to environmental cues [47]. Given their combined and robust downregulation, these data suggest that the SCs may be undergoing a metabolic shift, allocating cellular resources for energy production and autophagy rather than myelin maintenance or proliferation [53,54]. Within the same treatment group, OLR1583, Ins2, and Esr1 were strongly upregulated, which are involved in the necrosis and autophagy pathways [54]. Although supplementary roles of these genes support the inference that the SCs are indeed shifting toward a pro-survival phenotype. Aside from smell, olfactory receptors can aid in modulating intracellular signaling in response to a dynamic extracellular environment [55,56]. In addition, they can influence myelin maintenance pathways, which rely on lipid synthesis and degradation. Acting as an extracellular sensor, Olr1583 may facilitate metabolic adaptations and reallocation required for myelin recycling and maintenance [57,58]. Furthermore, Ins2 and Esr1 have been found to have supporting roles in these processes. Upregulation of the Ins2 gene suggests an increased activation of insulin signaling pathways, such as the MAPK/ERK cascade. These pathways are known to promote autophagy and survival under cellular stress [59]. In an accompanying role, it is well established that increases in estrogen-related pathways play critical roles in cellular homeostasis, particularly supporting autophagy [60,61]. Taken together, the combined downregulation of Maco1 and Mag, along with the upregulation of Olr1583, Ins2, and Esr1, supports the consensus that serum-free media in the absence of H_2_O_2_ increases transcription of genes associated with a phenotypic and metabolic shift toward autophagy or cellular stress when compared to serum-containing media. Alternatively, the increased survival phenotype could be induced by serum starvation, which has historically shifted SCs into a survival state [62]. This pro-survival state has been shown to be reversible upon re-exposure to serum.
4.8. Chemically Defined Media Facilitates Pro-Regenerative States and Cell Metabolism in the Presence of Deferoxamine Mesylate and 62.5 mM H2O2
When both serum-free CDM and serum-containing media, D10S, were treated with DFO in the presence of 62.5 mM H_2_O_2_, the transcriptional landscape was largely changed (Figure 8). The most prominent fold changes were observed in Defb1 (21.09) and Tnfrsf11b (−17.64). Beta defensins, such as Defb1, are known to directly participate in redox-dependent mechanisms. Their reduced form improves anti-microbial function [63]. In SCs, increased ROS levels could induce Defb1 expression to mitigate oxidative stress. The observed upregulation suggests an active oxidative stress management response, which can result from increased ROS production during high metabolic activity or in the presence of H_2_O_2_ [64]. Conventionally, an increase in oxidative stress would concurrently show a rise in Tnfrsf11b expression. This upregulation can act as a “gauge” of cellular stress, as elevated Tnfrsf11b exacerbates ROS production by mitochondria [65]. If oxidative damage begins to overwhelm these compensatory mechanisms, Tnfrsf11b expression would further increase, ultimately shifting the SC from autophagy toward an apoptotic phenotype [66]. Combined, both the upregulation of Defb1 and downregulation of Tnfrsf11b suggest the SCs are actively managing oxidative stress, carefully balancing autophagy and apoptosis. Supporting evidence includes the suppression of Bcl2l11 and Bcl1l12, which is consistent with the assumption that the SCs are indeed successfully regulating oxidative stress [59,67,68]. Additionally, changes in Hspbap1 (11.68), Sycp2 (7.45), Ins2 (6.5), Birc3 (−5.42), and Htt (4.64) further indicate a pro-autophagy phenotype, favoring intracellular stability [69,70,71,72]. Notably, Ins2 upregulation suggests an adaptation needed to meet the increased energy demands of these metabolic changes [73], while upregulation of Htt indicates a persisting myelinating phenotype [74]. Overall, the observed transcriptional changes when comparing serum-free vs. serum-containing media indicate that the former promotes autophagy and cellular recycling at greater levels when in the presence of 62.5 mM H_2_O_2_ and DFO. This adaptation permits the continued repair and maintenance required for SCs to retain their pro-repair and regenerative phenotypes in the more hostile microenvironment of injury.
5. Conclusions
We have investigated the efficacy of deferoxamine mesylate (DFO) in protecting ventral root Schwann cells (SCs) isolated from adult Sprague Dawley rats from H_2_O_2_-induced cell death. Our results show that pre-treatment using DFO increases SCs survival in serum conditions but has no effect in serum-free conditions, which is not consistent with our hypothesis. Instead, in serum-free conditions, SCs survive H_2_O_2_-induced cell death regardless of being pre-treated with DFO. Our analysis of SC phenotype and transcriptomics reveals a pro-repair and pro-survival state of SCs in serum-free conditions. In serum, DFO maintained efficacy in preventing cell death. Per our hypothesis, SCs were expected to have an increased survival upon DFO pretreatment; our results have shown that it is only the case when SCs are grown in serum-containing conditions. Without serum, SCs innately turn into a pro-survival state.
Previously, studies on DFO on increasing SC survival have been performed in serum-containing conditions; our results show a more pro-survival state of SCs in serum-free media, making DFO seemingly unnecessary. Thus, our findings highlight the importance of media differences and the need to replicate clinically used, serum-free media in in vitro and in vivo studies to accurately reflect SC physiology when treated with factors.
Translationally, we have shown that DFO is not worthy of further investigation since it fails to increase cellular survival in serum-free mediums. However, it emphasized the importance of studying the effects of similar pharmaceutical agents in serum-free conditions to accurately reflect clinical practice. Future studies must consider the effects of serum in modifying Schwann cell phenotypes when conducting a similar study. Additionally, it may be of interest for future studies to ascertain the molecular mechanisms of how the use of serum-free medium or serum starvation shifts SCs into a pro-survival state and thus increases SC survival in vivo.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bennett J. Das J.M. Emmady P.D. Spinal Cord Injuries Stat Pearls [Internet]Stat Pearls Publishing Treasure Island, FL, USA 2024 Available online: https://www.ncbi.nlm.nih.gov/books/NBK 560721/(accessed on 23 August 2024)
- 2O’Shea T.M. Burda J.E. Sofroniew M.V. Cell biology of spinal cord injury and repair J. Clin. Investig.20171273259327010.1172/JCI 9060828737515 PMC 5669582 · doi ↗ · pubmed ↗
- 3Tran A.P. Warren P.M. Silver J. The Biology of Regeneration Failure and Success After Spinal Cord Injury Physiol. Rev.20189888191710.1152/physrev.00017.201729513146 PMC 5966716 · doi ↗ · pubmed ↗
- 4Hill C.E. A view from the ending: Axonal dieback and regeneration following SCI Neurosci. Lett.2017652112410.1016/j.neulet.2016.11.00227825985 · doi ↗ · pubmed ↗
- 5Bunge M.B. Pearse D.D. Transplantation strategies to promote repair of the injured spinal cord J. Rehabil. Res. Dev.200340(Suppl. 1)556210.1682/JRRD.2003.08.005515077649 · doi ↗ · pubmed ↗
- 6Assinck P. Duncan G.J. Hilton B.J. Plemel J.R. Tetzlaff W. Cell transplantation therapy for spinal cord injury Nat. Neurosci.20172063764710.1038/nn.454128440805 · doi ↗ · pubmed ↗
- 7Ribeiro B.F. da Cruz B.C. de Sousa B.M. Correia P.D. David N. Rocha C. Almeida R.D. Ribeiro da Cunha M. Marques Baptista A.A. Vieira S.I. Cell therapies for spinal cord injury: A review of the clinical trials and cell-type therapeutic potential Brain 202314626722693 Erratum in Brain 2023, 146, e 128. https://doi.org/10.1093/brain/awad 20410.1093/brain/awad 04736848323 · doi ↗ · pubmed ↗
- 8Deng L.X. Walker C. Xu X.M. Schwann cell transplantation and descending propriospinal regeneration after spinal cord injury Brain Res.2015161910411410.1016/j.brainres.2014.09.03825257034 PMC 4375094 · doi ↗ · pubmed ↗