Molecular Determination of Sex from Down and Feather in Wild and Reared Monomorphic and Dimorphic Birds at Juvenile Age
Antonio Ciro Guaricci, Mario Cinone, Salvatore Desantis, Giovanni Michele Lacalandra, Maria Albrizio

TL;DR
This paper shows how DNA from down and feathers can be used to determine the sex of young birds, helping in breeding and conservation efforts.
Contribution
The study demonstrates a non-invasive method for early sex determination in birds using DNA from feathers and down.
Findings
DNA from down and feathers is a suitable and non-invasive source for sex determination in young birds.
The method successfully identified the sex of 153 avian species, including 27 for the first time.
All tested pairs produced offspring, confirming the accuracy of the sexing method.
Abstract
Interest in the breeding of feathered species has grown increasingly in recent years to the extent that early sexing has become necessary to ensure the formation of the pairs that will guarantee the birth of new birds. When sex identification must be carried out in very young individuals, sex is hardly recognizable not only in monomorphic species but also in dimorphic ones. The employment of molecular techniques is particularly necessary when the assessment of biometric parameters does not allow a clear sex identification. Recovery of DNA is mandatory when molecular techniques are used. In this paper, we demonstrate that down and feathers are suitable sources for DNA recovery without causing severe stress to the bird. This is a crucial aspect when dealing with nestlings. Using this procedure, we identified the gender of 153 avian species, including 27 for the first time. The inability…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
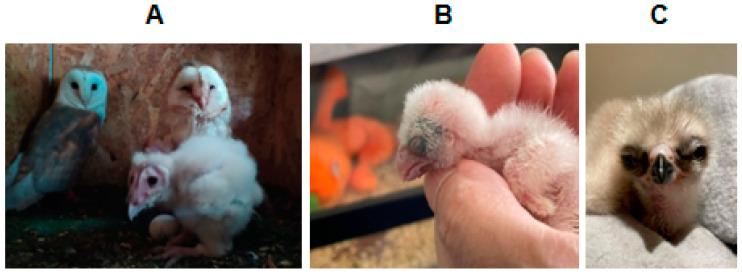 Figure 1
Figure 1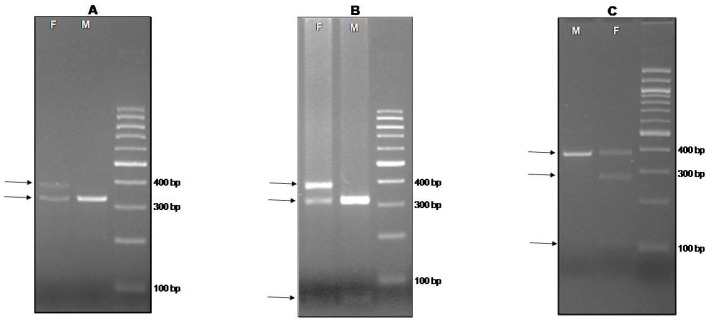 Figure 2
Figure 2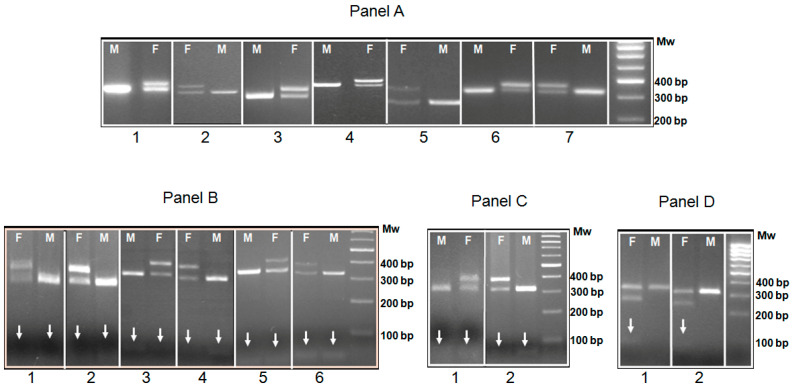 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetic and Clinical Aspects of Sex Determination and Chromosomal Abnormalities · Avian ecology and behavior · Animal Behavior and Reproduction
1. Introduction
In recent years, bird breeding has found renewed interest in several countries, both for basic scientific reasons and because of the gradual spread of ornamental–amateur value species breeding. In addition, the number of projects for the reintroduction of threatened species [1] and programs for integrated conservation through the release of wild or captive-bred individuals [2] has grown enormously. About 60% of bird species are monomorphic [3], making it difficult to identify the sex of juvenile and adult individuals.
Among the various problems to be addressed in the context of breeding plans for native and/or exotic birds, the knowledge of sex has primary importance for the correct pairing of the subjects that will constitute the pair. The minimal or no sexual dimorphism in many avian species makes sex determination based on morphological features unreliable and virtually impossible in juvenile subjects [4]. This difficulty is also evident in newly hatched birds [5].
Sex can also be determined through acoustic venting [6], laparoscopy [7], steroid levels [8], karyotyping [9], differences in morphometric traits [10], or by comparing blood plasma protein profiles between males and females [11]. However, all these approaches are not always suitable due to the long time needed to perform some of them or because of invasiveness and high cost or inapplicability to monomorphic species.
By contrast, molecular methods for sex identification are more feasible. Starting from 1996, the heterogameticity of the females (they have a Z and a W chromosome) and the homogameticity of the males (they have two equal Z chromosomes) were employed to highlight the differences in the nucleotide sequences of specific genes. Some genes were proposed as markers of sex identification based on variations in their intron lengths. One of the most used was the Chromodomain-Helicase DNA Binding Protein 1 (CHD1) because of its sex-linked intron length showing slight differences between the Z and W chromosomes. Using a specific primer pair and PCR, an amplification product is obtained that migrates as a single band in ZZ males and as two bands in ZW females when electrophoresed in agarose gel under the effect of an electric field [12,13].
In recent years, many molecular methods have been developed and employed for sexing avian species: single-strand conformation polymorphism (SSCP), restriction fragment length polymorphism (RFLP), random amplified polymorphic DNA (RAPD), amplified fragment length polymorphism (AFLP), microsatellites, allele-specific PCR (AS-PCR), capillary electrophoresis, real-time quantitative PCR (qPCR), real-time PCR combined with melting curve analysis, high-resolution melting (HRM) analysis ([14] for review), and single nucleotide polymorphism (SNP) [15]. These techniques overcome the main animal health risk deriving from surgery. Molecular sexing is virtually applicable to any bird, regardless of age, and becomes the best choice if it is aimed at sexing born offspring. The PCR technique can also be combined with the amplicon digestion by restriction endonucleases (PCR-RFLP) when the size of the amplified product shows the same size in the two sexes [16,17,18]. Different sources such as blood, feathers, feces, eggshell membrane [14], and cells from buccal swabs can be used to obtain DNA [19]. In this study, the RFLP-associated PCR method was used when the amplicons of the CHD1 gene showed non-appreciable differences between the two sexes. The aim of this work was to demonstrate that primers P2 and P8, first used by Griffiths et al. [20], are suitable for sexing more than 150 avian species belonging to different orders when combined with RFLP in wild and reared chicks. The merit of this work was to provide those interested in bird sexing with the correct relationship between species and testing methods. Although feather picking may induce temporary discomfort or stress, the birds are ultimately released unharmed, so we have shown that breast feathers are an adequate source for extracting enough DNA, especially for juvenile subjects.
2. Materials and Methods
This research was conducted over a seven-year period. Adult individuals were initially tested to see whether the couple of primers employed in the PCRs were effective in amplifying the CHD1 gene in all bird species analyzed and in distinguishing between males and females. When the PCR product was obtained without differences between the two sexes, the restriction enzyme HaeIII was tested, and when the restriction fragments produced were also not able to distinguish males from females, the restriction enzyme Asp 700I was used. Subsequently, once the correct PCR approach was established for each species, young birds were sexed, and approximately 70% of the pairs formed produced offspring during this study. In total, about 3500 samples from 153 different bird species were analyzed.
2.1. Sample Collection
Three to four down or feathers retrieved from the chest of adults, newly hatched chicks or juvenile birds (Figure 1) were provided by breeders, ornithologists, wild avifauna rehabilitation centers, and zoos. Samples retrieved from the Mediterranean area were brought directly to our laboratory or arrived by mail and courier. In some cases, samples arrived even 2–4 weeks after collection. Upon arrival, down/feathers were analyzed immediately. All the suppliers were recommended to collect feathers and down directly from the birds, avoiding the recovery of those fallen on the ground or in the cages.
2.2. DNA Extraction
Pieces of calamus 1–2 mm long from three feathers or down were used as genomic DNA sources. Extraction was performed using a commercial kit (GenElute mammalian genomic DNA miniprep kit, Sigma-Aldrich, Milan, Italy) according to the manufacturer’s instructions with slight optimized modifications, such as the digestion of samples in lysis buffer T added with proteinase K 10 mg/mL at 56 °C overnight and the final elution of the DNA from the binding silica using only 100 μL of TE buffer. The obtained DNA was quantified by a UV spectrophotometer (Beckman Instruments Inc. Fullerton, CA, USA), its purity assessed (ratio A260/A280 = 1.8–2.0) and finally stored at −20 °C until PCR analysis.
2.3. PCR and RFLP Analysis
The polymerase chain reaction was performed using the P2-forward and P8-reverse primers (Sigma-Aldrich, Milan, Italy) proposed by Griffiths et al. [20] in 1998, in a volume of 50 μL with 100 ng of DNA, 1U Hot Master Taq polymerase (Eppendorf, Milan, Italy), 200 μM each dNTP and 1 X Taq-buffer. The amplification profile was the following: an initial denaturation step at 95 °C for 2 min, 35 cycles at 95 °C for 45 s, 48 °C for 45 s and 72 °C for 45 s, and a final elongation step at 72 °C for 5 min.
PCR products were run on 3% agarose gel in TAE buffer with 0.5 μg/mL ethidium bromide and a molecular weight marker (Eppendorf, Milan, Italy) to verify the dimensions of the resulting fragments. The gels were visualized under UV light with GelDoc 2000 apparatus (BioRad, Milan, Italy). The acquired images were analyzed to evaluate the sizes of the amplified products.
Restriction fragment length polymorphism analysis was conducted by digesting 15 μL of the PCR product with 5U of HaeIII enzyme (Roche Diagnostics, Milan, Italy) [16], 1 X corresponding digestion buffer in a total volume of 20 μL at 37 °C for 3 h; the samples were run on a 2% agarose gel and visualized under UV light with GelDoc 2000, as previously described. When the Hae III restriction enzyme was not useful for producing different-size fragments in the males and females, a new RFLP analysis was performed on another 15 μL aliquot of the same amplified product with 5 U of Asp 700I (Roche Diagnostics, Milan, Italy) [16] at the same conditions of incubation, electrophoresis, separation, and UV visualization.
3. Results
To test that the P2/P8 primer pair was capable of amplifying and discriminating juvenile males and females in all avian species belonging to the orders listed in Table 1, a preliminary analysis was conducted on adult individuals of known gender of the same species. For all samples received, even those collected up to one month before analysis, the extracted DNA was of good quality and in a concentration more than sufficient for subsequent analysis. In males, regardless of order and species, the amplicon consisted of a single band that was characterized by gel electrophoresis and whose approximate size is shown in Table 1 and Figure 2.
As for the females, the species belonging to the orders of Otidiformes, Podicipediformes, and Sphenisciformes showed a double amplicon of different sizes (shown in Table 1), so sex identification was easily performed by a simple PCR reaction followed by gel electrophoresis (Figure 2A).
All the other orders listed in Table 1 required the use of the PCR-RFLP approach for most species. All species were sexed by this technique, and the approximative size of the fragments resulting from the enzymatic digestion by HaeIII is listed in Table 1 and Figure 2, panel B.
In detail, in the order of Accipitriformes, the following species required PCR: Cathartes aura, Gypohierax angolensis, Neophron percnopterus, Pernis apivorus, and Sarcoramphus papa. All the others were sexed by HaeIII digestion. In the order of Anseriformes, HaeIII digestion was necessary for the species Anas platyrhynchos.
The same procedure was followed for the orders of Apodiformes (species Apus apus), Bucerotiformes (species Aceros nipalensis, and Bucorvus leadbeateri), Charadriiformes (species Ichthyaetus melanocephalus, Himantopus Himantopus, and Vanellus vanellus), Ciconiiformes (species Ciconia ciconia), Columbiformes (species Streptopelia decaocto), Coraciiformes (species Merops apiaster), Cuculiformes (species Cuculus canorus), for the order of Falconiformes (species Falco vespertinus, Falco ardosiaceus, Falco biarmicus, Falco jugger, Falco naumanni, and Falco subbuteo), Galliformes (species Coturnix coturnix and Polypectron napoleonis), Gruiformes (species Balearica regulorum, Fulica atra, and Grus grus), Passeriformes (species Monticula solitarius, Oriolus oriolus, Parus major, and Serinus serinus), Pelecaniformes (all species required PCR-RFLP except for Egretta garzetta), and Suliformes (the species Phalacrocorax carbo).
In the order of Strigiformes, the species Strix aluco, Strix rufipes, and Strix uralensis were sexed by simple PCR, while all the others were sexed by HaeIII digestion.
When even PCR-RFLP by HaeIII did not allow for the differentiation between females and males, the restriction enzyme Asp 700I was employed in the PCR-RFLP procedure (Figure 2, panel C). This approach was used to sex Chauna chavaria of the order Anseriformes and Phoenicopterus chilensis and Phoenicopterus roseus of the Phenicopteriformes order. The approximative sizes of the fragments produced after the enzymatic digestion of the amplicon are listed in Table 1.
After defining the size of the amplicons and/or restriction fragments in each species, we subjected the DNA obtained from the down of young individuals to amplification. Every young subject was easily sexed, obtaining the expected amplicon or restriction fragment. By this approach, to the best of our knowledge, 27 new species (shaded row in Table 1 and Figure 3) were sexed for the first time.
4. Discussion
Phenotypic traits cannot help in the sex determination of monomorphic species. To overcome this problem, a molecular PCR-based test to define the gender of 153 different avian species was employed.
Sex identification in birds is of considerable importance for many reasons, ranging from understanding changes in sex ratio between generations or in specific geographic regions to assessing the health, behavior, and ecological dynamics of populations and facilitating reproduction in captivity. In birds, predicting sex is a difficult task because temperature or other environmental factors have no effect on avian sex determination or gonadal development as they influence these processes in non-avian reptiles, amphibians, and some fish [21].
Furthermore, early sex determination is mandatory for managing captive birds and improving breeding programs because it allows for precocious pair formation.
Considering this primary objective, this study evaluated the efficacy of DNA extraction from a small number of down collected easily and with slight pain from the chest of nestling or juvenile birds. Previous studies have shown that up to 10 ng/mL of DNA is obtained from adult small feathers. This DNA is also of good quality and virtually contamination-free because, during growth, feathers supplied with blood keep the DNA inside the stem, protecting it from potential degradation and microbial contamination [22]. In our case, using growing feathers from young individuals, the DNA concentration obtained was around 15 ng/mL. This result was also achieved by making some minor adjustments to the extraction protocol, such as overnight digestion of the tissue and genomic DNA elution with only 100 mL buffer.
The recovery of DNA from a biological source that minimizes stress and eliminates surgical risk for the bird supports the idea of Peniche et al. [23] that molecular sexing is the only efficacious technique for sexing certain avian species, such as golden eagle chicks, especially when overlapping biometric measures cannot be used to differentiate between sexes at an early age.
The source of DNA extraction is a key point in determining the eligibility of the molecular approach for sex determination. Additionally, the employment of avian sex-specific primers eliminates the possibility of contaminant DNA amplification deriving from naturally present microorganisms. Peniche et al. [23] also proposed such an approach, but the extraction of DNA from blood or oral sampling, compared to that from down or feather, is more invasive and stressful if applied to very young individuals [24].
Therefore, the results of this study could provide a guideline for species that have never been sexed before (18% of all species that we sexed), which also include species that, although dimorphic as adults (such as bird of prey), cannot be differentiated by sex in their early stage. This is also the case for Accipiter nisus, which shows clear dimorphism in adulthood, when males show a specific orange plumage on the chest and flanks [25], but during the first six to eight months of age, even the laparoscopic observation of the gonads cannot help because the ovaries may resemble testicles [26].
Until now, several molecular approaches have been proposed to sex the majority of avian species, but their costly and time-consuming procedure have resulted in low implementation.
Some species included in this study had already been sexed by other authors. Canon et al. [27] employed cytofluorimetry in Amazona amazonica, Melopsittacus undulatus, and Nymphicus hollandicus with limitations due to overlapping DNA weights between the two genders and the failure of intercalating dye as a result of degradation. Elnomrosy et al. [28] employed loop-mediated isothermal amplification to sex some species of the Psittacidae, Cacatuidae, and Psittaculidae families. The method appears time-consuming and is not widely applicable because it needs specific settings for almost every species. Horng et al. [29] used random amplified polymorphic DNA (RAPD-PCR) in Columba livia, and although it does not require a known prior sequence of template, it needs careful planning for each species. Hence, RAPD using short primers to produce DNA fragments by PCR is affected by low reproducibility and sensitivity to reaction conditions and can be a species-specific method, as evidenced by Griffiths and Tiwari [30]. Also, the nuclear microsatellite amplification using DNA from feathers can produce errors if the DNA is highly fragmented or is in low concentrations, as demonstrated by Mills et al. [31]. The amplified fragment length polymorphism (AFLP) method requires acrylamide gels, radioactive markers, or primers purified by HPLC, so it becomes an expansive technique [32].
By contrast, the molecular approach utilized in this study has proved effective in sexing all tested species. A further point confirming this is the fact that all pairs formed following sexing, housed in the same cage, and reaching sexual maturity during this study, produced offspring. These represented about 70% of all sexed species. The method could be applied to other species, starting with amplification by P2/P8 and continuing with RFLP if the simple PCR does not discriminate against gender. Obviously, mutations at the level of the consensus region of the employed primers or at the restriction sites for HaeIII and Asp 700I are possible limitations of this approach, but this is the limit of all molecular techniques.
5. Conclusions
This is the first report showing that a simple molecular approach can be used to successfully sex more than 150 avian species. We demonstrated the usefulness of down retrieved from chicks as a source of sufficient DNA obtained by the optimization of its extraction procedure. The use of the well-established P2/P8 primers for PCR, eventually combined with the RFLP, allowed for the correct sexing of all juvenile birds. This means that researchers involved in reintroduction projects and the preservation of endangered birds, as well as professional or amateur breeders of pet birds, will find this publication a guide for determining the sex of newborns to facilitate early pair formation. With this fairly simple and inexpensive technique, zoos and wildlife rescue centers can easily identify bird pairs for captive breeding programs. The invasiveness of down and feather plucking, although moderate, remains a specific limitation of this protocol. Nonetheless, it is necessary if one wants to eliminate errors in sex assignment and, thus, in pair formation. This sexing protocol, if tested in bird species belonging to the other orders, could be a candidate to become the first applicable to sex almost all birds.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Seddon P.J. Armstrong D.P. Maloney R.F. Developing the science of reintroduction biology Conserv. Biol.20072130331210.1111/j.1523-1739.2006.00627.x 17391180 · doi ↗ · pubmed ↗
- 2IUCN (World Conservation Union) Guidelines for Re-introductions IUCN/SSC Re-introduction Specialist Group, IUCN Gland, Switzerland Cambridge, UK 1998
- 3Wu C.P. Horng Y.M. Wang R.T. Yang K.T. Huang M.C. A novel sex-specific DNA marker in Columbidae birds Theriogenology 20076732833310.1016/j.theriogenology.2006.08.00116965809 · doi ↗ · pubmed ↗
- 4Lee M.Y. Hong Y.J. Park S.K. Kim Y.J. Choi T.Y. Lee H. Min M.S. Application of two complementary molecular sexing methods for East Asian bird species Genes. Genom.200830365372 Available online: https://hdl.handle.net/10371/8289(accessed on 6 February 2025)
- 5Forsman D. The Raptors of Europe and the Middle East A Handbook of Field Identification T. & A.D. Poyser London, UK 1999
- 6Volodin I. Kaiser M. Matrosova V. Volodina E. Klenova A. Filatova O. Kholodova M. The technique of noninvasive distant sexing for four monomorphic Dendrocygna whistling duck species by their loud whistles Bioacoustics 20091827729010.1080/09524622.2009.9753606 · doi ↗
- 7Richner H. Avian laparoscopy as a field technique for sexing birds and an assessment of its effects on wild birds J. Field. Ornithol.198960137142
- 8Bercovitz A.B. Czekala N.M. Lasley B.L. A new method of sex determination in monomorphic birds J. Zoo Anim. Med.1978911412410.2307/20094391 · doi ↗