Understanding the genetic complexity of Leishmania infantum in the Americas: a focus on 3’NT/NU gene deletion
Monique Florêncio, Elisa Cupolillo, Mariana Côrtes Boité

TL;DR
This paper reviews the genetic diversity of Leishmania infantum in the Americas, focusing on a deletion in the 3’NT/NU gene and its potential role in adaptation.
Contribution
Highlights the underappreciated genetic complexity and adaptive strategies of L. infantum in Brazil due to 3’NT/NU gene deletion.
Findings
A subpopulation of L. infantum with a homozygous deletion in the 3’NT/NU gene is prevalent in Brazil.
The deletion may confer adaptive advantages under Brazilian ecological conditions.
The 3’NT/NU enzyme is a key virulence factor in L. infantum.
Abstract
Visceral Leishmaniasis in the Americas is primarily associated with Leishmania (Leishmania) infantum. This parasite is non-native and was imported during the colonisation era. The constitutive instability of the Leishmania genome allows this parasite to express flexibility in adapting to environmental fluctuations and different selective pressures, such as those the parasite faced when arrived in the New World. Therefore, genetic diversity is expected among the populations of L. infantum in the Americas, despite the bottle neck of importation route. Indeed, subpopulation of strains of L. infantum carrying a homozygous deletion in the genome was detected exclusively in the continent. These strains are more spread across Brazilian territory to the detriment of the non-deleted; the locus includes four genes, two of which encode the enzyme ecto-3’-nucleotidase/nuclease (3’NT/NU), a…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
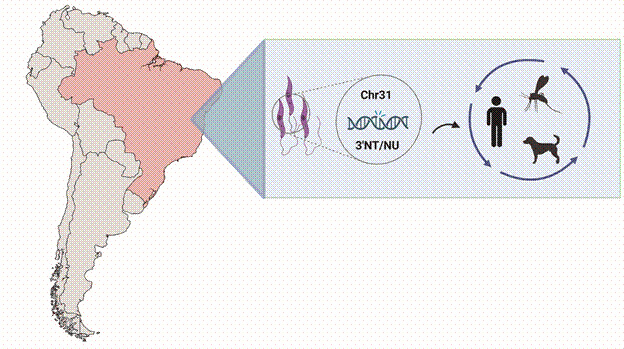 Figure 1
Figure 1- —MCB-PTR
- —FIOCRUZ-PAEF
- —CAPES
- —EC
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsResearch on Leishmaniasis Studies · Trypanosoma species research and implications
Leishmaniasis has three main clinical forms: visceral, cutaneous and mucocutaneous. According to the World Health Organization (WHO), constitutes a major public health problem around the globe, mainly affecting poverty-stricken regions. It is endemic in 92 countries, of which 72 are developing countries, and where one billion people are living at risk of contracting the disease.1 In the New World (NW), Brazil leads the number of cases.2 ^,^ 3 Leishmaniasis is an ancient disease, with several primitive descriptions found in archaeological findings and in molecular analyses of mummified human remains.4 ^,^ 5 ^,^ 6 There are three hypotheses to explain the origin of the genus Leishmania, all under ongoing debate.5 One of the most accepted is the supercontinent hypothesis, proposing that during the separation of Gondwana in the Mesozoic era, the subgenus Viannia evolved in South America, while the subgenera Sauroleishmania and Leishmania developed in Africa.7 Enzymatic and molecular analyses have corroborated that the origin of the agent of visceral leishmaniasis in the Americas - L. infantum, occurred during the period of European colonisation, approximately 500 years ago.7 ^,^ 8 ^,^ 9 It is a parasite imported by the colonisers. It is highly probably that there were multiple introductions of these trypanosomatids carried by asymptomatic dogs and other mammalian hosts. Once in the NW, this parasite found appropriate ecological conditions for transmission, such as the presence of its main urban reservoir, Canis familiaris, but with important differences from those found in the Old World (OW), such as the existence of permissive sandfly vectors. The scenario favoured parasite adaptation and the establishment of new transmission cycles.10 Therefore, these small founding populations of L. infantum have been facing selective pressures, distinct from those found in the OW, as well as unknown sampling effects that drive genetic drift.11 In addition to this historical epidemiological scenario and the evolutionary processes that permeate the parasite arrival in the NW, is the fact that Leishmania presents constitutive genomic adaptability. Among the molecular traits involved in such characteristics are: (i) the mosaic aneuploidy, a phenomenon where the number of copies of individual chromosomes varies within and between cells of a clonal population,12 ^,^ 13 configuring itself not only as a product of genomic plasticity, but as a reflection of this inherited trait, and (ii) a complex and predominantly clonal life cycle, presenting hybridisation events in an unknown frequency14 ^,^ 15 and a close ecological relationship with its hosts.16 ^,^ 17 ^,^ 18 Such characteristics are of great relevance for the biology and adaptability of the parasite and thus for the associated eco-epidemiological scenario. For instance, intra-specific genetic variation can be associated with the main differences in disease pathology19 and drug resistance.18 Importantly, losing genes is also a significant adaptive strategy for Leishmania.13 In this regard, Carnielli et al.,20 through a genome-wide association study (GWAS), identified populations of L. infantum in Minas Gerais, Maranhão and Piauí (Brazil) carrying a large deletion (> 12 kb) found in the often reported as a one of the most stable chromosome within Leishmania mosaic aneuploidy, the polysomic chromosome 31 (chr31)18 ^,^ 20 ^,^ 21 (as depicted in Figure). The genomic trait was associated with the phenotype of resistance to an important leishmanicidal drug, miltefosine, and thus named miltefosine sensitivity locus (MSL).20 The homozygous deletion covers four open reading frames: LinJ.31.2370 (ecto-3′-nucleotidase/nuclease), LinJ.31.2380 (ecto-3′-nucleotidase/nuclease precursor), LinJ.31.2390 (helicase-like protein), and LinJ.31.2400 (3,2-trans-enoyl-CoA isomerase). Afterwards, Schwabl et al.11 performed a comparative analysis of a wide panel of L. infantum genomes [107 from the NW and 19 from the OW, plus 75 additional samples from the NW by quantitative polymerase chain reaction (qPCR)], confirming the occurrence of the deletion. The deletion-carrying strains (Del) were found exclusively among American samples (mainly from Brazil - 126 of 177), while all available OW genomes examined were non-deleted (NonDel).11 Remarkably, Del parasites were more frequent and geographically widespread then NonDel strains. To this peculiar finding adds the functional description of two deleted copies within the site, encoding the enzyme ecto-3’-nucleotidase/nuclease (3’NT/NU). The enzyme plays an important role in parasite nutrition, establishment of infection in vertebrate host,22 uptake of purines,23 escape of parasites from neutrophil networks (NETs)24 and the ability to infect macrophages.25 ^,^ 26 Therefore, this enzyme is expected to be quite relevant for the parasite in the initial stages of infection and constitutes a virulence factor for L. infantum.22 ^,^ 23 Despite its projected importance, it has been confirmed that Del strains indeed lack 3’Nu activity in vitro.11 An open question, therefore, is how these deficient parasites are dispersing more successfully across Brazilian territory to the detriment of the NonDel strains, and what are, if any, the associated epidemiological consequences. One hypothesis is that the deletion unexpectedly increases parasite fitness in Brazilian ecological landscape. In this review, we discuss the aspects that may relate to the spread of deletion-carrying strains, possible implications of the deleted locus on the virulence, infectivity, and transmissibility of L. infantum in Brazil, and the epidemiological consequences of this scenario.
A subpopulation of Leishmania infantum strains with a homozygous deletion in the genome was detected exclusively in the Americas, predominantly in Brazil, at the expense of non-deleted strains. Among the deleted genes, two encode the enzyme ecto-3’-nucleotidase/nuclease (3’NT/NU), a known virulence factor in L. infantum. The epidemiological consequences and implications for the transmission cycle remain an open question, and represent a unique opportunity to advance our understanding of adaptive and evolutionary molecular strategies in Leishmania.
Aspects on the ecological scenario of L. infantum population in Brazil
Upon arriving in the Americas, L. infantum had to adapt to different, but suitable ecological conditions to establish new transmission cycles. This includes NW vertebrate hosts such as dogs in the domestic and peri-urban environment and sylvatic animals: endodentates, carnivores, marsupials, rodents, and primates,27 mostly diverse from the wildlife species that have been parasitised in the OW.28 The main vector species in the Americas is Lutzomyia longipalpis, a sandfly belonging to a different genus from that of the OW, Phlebotomus.29 The climatic conditions in NW are also distinct, favouring sandflies development trough out the year,28 unlike the scenario observed in the Mediterranean region, where the vector population presents a more defined seasonal pattern, typically from spring to autumn.30 Moreover, Lu. longipalpis exhibits a complex population structure, evidenced by chemical, behavioural and molecular traits.31 More recently, in Brazil, other sandfly species have been pointed as vectors, such as Lu. cruzi 32 or as putative vector, such as Lu. migonei. 33 This fact, combined to the American biomes, distinct from those in Europe, have likely contributed to the establishment of locally specific transmission cycles of the American visceral leishmaniasis (AVL).34 Several studies have shown that it is probable that the transmission, virulence, and clinical outcome are strongly influenced by coevolutionary interactions between the parasite species and specific sandfly genotypes.35 ^,^ 36 Thus, constitutive genomic characteristics of L. infantum (syn. Leishmania chagasi) might emerge as a reflex of the environmental selective pressures to which they are subjected to. In Brazil, Ferreira et al.37 examined by multilocus microsatellite typing (MLMT) 162 samples of L. infantum from dogs and humans, and a sample of fox (Cerdocyon sp.) and opossum (Didelphis marsupialis), from most Brazilian states and Paraguay. Despite the homogeneity expressed by multilocus enzyme electrophoresis (MLEE), MLMT exposed the circulation of three distinct parasite populations: POP1 composed of 111 strains and 36 genotypes, POP2 with 31 strains and 19 genotypes and POP3 with 31 strains and 12 genotypes. Curiously, the geographical distribution of these strains was not homogeneous. POP1 was found in 16 Brazilian states and in Paraguay, being predominant in some geographic areas, such as in Espírito Santo, where 95% of the strains were assigned POP1. In contrast, POP2 was found in 10 Brazilian states, but predominant only in Mato Grosso. POP3 was predominant in Mato Grosso do Sul (75%). When overlapped with reports of sandflies, one finds that Lu. cruzi, a closely related species to Lu. longipalpis,38 ^,^ 39 participates in transmission cycle of L. infantum in Mato Grosso and Mato Grosso do Sul.39 ^,^ 40 It is plausible, thus, to assume there is a parasite-sandfly association in that area. Schwabl et al.11 found data suggesting a similar pattern after screening the occurrence of Del L. infantum strains. Authors found the non-deleted strains (NonDel) were highly concentrated and frequent in the state of Mato Grosso do Sul, which, again, may be related to the alternative vector in this region, Lu. cruzi,11 or to the alternative introduction of parasites to that region by the bordering countries (e.g., Bolivia). Genetic exchange was already demonstrated for many Leishmania parasites, a process occurring manly, if not exclusively, in the sandflies.14 ^,^ 15 ^,^ 41 Contradicting the idea that the L. infantum population in Brazil is homogeneous and increasing the complexity of the population structure of this parasite, six heterozygous samples (HTZ) were described and confirmed as hybrids originated from Del and NonDel in contact zone regions.11 Therefore, and considering that parasite itself play a key role in clinical outcome,10 ^,^ 11 it is plausible to assume the epidemiological variations current described in AVL are not restrictedly linked to host and environment traits. The parasite, thus, is a major player to be considered, and the relatively short time L. infantum inhabits the continent should not be underestimated during epidemiological surveillance approaches.
Adaptation strategies and fitness gain in Leishmania vs the dispersion scenario of Del parasites
Schwabl et al.11 pointed out that the sub-chromosomal deletion found among American strains was possibly an inherited trait, i.e., was already present within the parasite population from Europe, then imported to Brazil. From the molecular point of view, Carnielli et al.20 suggested the deletion resulted from homologous recombination, in a genomic region flanked by smaller regions of DNA called repeated sequences (RSs). The circular extrachromosomal element produced was subsequently eliminated by the cell. Once the locus is lost, it remains absent from the population.20 Interestingly, despite potentially emerging also among OW strains, the deleted genotype was detected only in American samples, in a higher frequency and widely dispersed compared to the NonDel strains. The higher frequency, wide distribution, combined to restrict circulation to the Americas suggests greater fitness of Del samples to NW conditions. Yet, how a deficient parasite has managed to adapt so successfully in such distinct environment is an open question. The answer might be obtained from the genomic plasticity and peculiar gene expression regulation of Leishmania. It is reported that Leishmania has a unique, atypical, and unstable genome: it is organised in a peculiar way, i.e., functionally unrelated protein-coding genes are arranged in large polycistronic clusters.42 This parasite does have RNA polymerase II promoters, but they are not canonical promoters typically associated with protein-coding genes. The transcription occurs bidirectionally in regions between the polycistronic units, called strand switch regions, and then, the pre-mRNAs formed are processed into mature mRNAs.42 ^,^ 43 ^,^ 44 ^,^ 45 This results in weak transcriptional regulation of protein-coding genes.46 Moreover, the mosaic aneuploidy,12 and the predominantly clonal life cycle with frequent recombination events47 increases plasticity. Mosaic aneuploidy is expressed as the variation in the chromosome dosage between cells of the same clonal population, composed of tetrasomic, trisomic, disomic or monosomic cells in variable proportions.13 ^,^ 47 All Leishmania genomes current analysed expressed aneuploidy, but a stable polysomy uniquely in chromosome 31.48 ^,^ 49 The mosaic aneuploidy leads to changes in gene dosage16 and constitutes a strategy for the parasite to adapt to its complex life cycle in the vector and host, as well as to increase drug resistance.50 ^,^ 51 ^,^ 52 ^,^ 53 This intrinsic instability in the genome generates phenotypic and genetic variability intra and inter-strains,12 ^,^ 48 ^,^ 49 playing an important role in the biology and evolution of the parasite.54
Two distinct levels of biological organisation communicate in evolution: the molecular networks and the genes that encode their organisation and structure.55 ^,^ 56 Regarding molecular networks, two factors alter protein evolution: (i) translation selection, which refers to the evolutionary selection of characteristics that can increase translation efficiency; and (ii) functional constraint, which refers to the level at which random mutations are removed from the population by natural selection to avoid their deleterious effect on protein function.57 The enzyme in a metabolic pathway is also under the protection of functional constraint.56 This leads us to speculate that the deletion of 3’NT/NU may alter the purine salvage pathway in L. infantum, but does not necessarily represent a risk to the parasite. Despite the loss of 3’-ectonucleotidase activity and the possible reduction of uptake of purines, Leishmania can use compensatory means for this function, and/or adapt metabolically to this new nutritional condition. Thus, although the absence of the deleted genes may directly alter phenotype, the associated compensatory mechanisms for gene loss could additionally lead to relevant biological variation. Searching for signals of compensation, Schwabl et al.11 found significant copy number variation (CNV) between the NW Del and NonDel strains, in 38 coding regions. Although differences were mostly driven by population structure, after correcting for origin, five coding regions presented significant haplotype somy variance between the Del and NonDel. The annotated loci included nucleoside transporter 1, amastin-like protein and paraflagellar rod protein paralogs.11 These findings require further investigation to understand the linkage between the lack of the deleted genes and the CNV report. Amastin-like protein constitute a virulence factor in Leishmania and is expressed only in the form of amastigotes;58 ^,^ 59 nucleoside transporters, in turn, are responsible for mediating transport of nucleosides, nucleobases and therapeutic analogues into the parasite,60 ^,^ 61 and finally, paraflagellar rod protein are constituents of the paraflagellar rod, a unique network of cytoskeletal filaments that is located next to the axoneme in most trypanosomatids, essential for cell motility.62 ^,^ 63 Since correlation between CNV and abundance of transcripts has been described, it is expected to observe associated variation in phenotypes.64 ^,^ 65 To search for CNV and transcripts variation between Del and NonDel may reveal the molecular grounds for increased parasite fitness.
The sub-chromosomal 12Kb deletion among American L. infantum strains and associated features: a genetic marker?
During a phase 2 clinical trial in Brazil to test efficacy of miltefosine to treat human AVL, the disappointing rate of 60% of clinical cure was reported against the successful 94% rate in India. The reasons behind it remained blurred, until Carnielli et al. revealed the phenotype of natural drug resistance among parasites carrying a 12 Kb deletion in chromosome 31.20 The deleted genomic site, MSL was reported as marker for susceptibility to the drug. Patients infected with MSL^-^ parasites presented higher chance to treatment failure. Further on, in vitro assays using the same strains confirmed the association between the MSL and the susceptibility to miltefosine. An additional study, however, did not detect any correlation.66 The lack of convergence between these studies is a reflex of how controversial genetic markers in Leishmania might be. One of the reasons may be, among other molecular traits, the genomic plasticity of these parasites. So, phenotypes such as drug resistance are subjected to the various adaptive strategies the parasite develops to adapt.48 ^,^ 67 The search for a convergent, unique molecular path is, thus, frequently unproductive. Nevertheless, the deletion is located within the one of more stable chromosome among the mosaic aneuploidies.21 The loss of all homologous copies in the locus is, thus, remarkable and might indeed result in stable phenotypic variance. An additional feature to consider regarding the relevance of this natural deletion is the possible compensatory mechanisms developed by the parasite (addressed in next topics), and the annotated function of the four open reading frames within. We believe the main loci at the deleted site that could correlate or lead to vital phenotypic differences between Del an NonDel are the two copies of ecto-3’-nucleotidase/nuclease. The reasons for it are depicted bellow.
The ecto-3’-nucleotidase: why it calls up attention in detriment to the other deleted genes
The ecto-3’-nucleotidase/nuclease (3’NT/NU) enzyme was first described by Gottlieb and Dwyer68 through the isolation and purification of surface mem-branes from L. donovani promastigotes. Later was found in other trypanosomatids including L. infantum.26 It is a 43 kDa transmembrane enzyme from the family of class l nucleases, bi-functional, capable of hydrolysing 3’-monophosphorylated nucleotides and nucleic acids, configuring itself as a phosphodiesterase or phosphomonoesterase.68 ^,^ 69 This enzyme plays an important role in the nutrition of the parasite and in the establishment of infection in the vertebrate host.22 Leishmania are auxotrophs for purines, unable to synthesise them via de novo. For this reason, they make use of the purine salvage pathway to capture preformed nucleotides from the host cell and synthesise their purine nucleotides, necessary for nucleic acid synthesis and other relevant biomolecules.70 ^,^ 71 3’NT/NU participates in this process, i.e., the uptake of purines by hydrolysing extracellular nucleotides, conversion into nucleosides such as adenosine,23 and transport into the parasite by nucleoside/nucleobase transporters (NT).61 In addition to nutritional factors, 3’NT/NU is also associated with the escape of parasites from neutrophil networks (NETs), due to its ability to hydrolyse nucleic acids,24 and the ability to infect macrophages.25 ^,^ 26 Besides this, the fact of 3’NT/NU does not occur in mammals, makes it a potential therapeutic target.25 The literature reports that 3’NT/NU has greater activity in infective metacyclic forms and is absent in amastigotes. Due to these characteristics, this enzyme is likely to be more relevant for the parasite in the initial stages of infection and constitutes a virulence factor for L. infantum.22 ^,^ 23 Vieira et al.26 have demonstrated that the activity of this enzyme is superior in viscerotropic species of Leishmania. L. infantum was the species that demonstrated the greatest 3’-nucleotidase activity, followed by L. donovani. In the same work, it was also demonstrated that the activity of 3’NT/NU is 10-fold higher than the activity of the enzyme ecto-5’-nucleotidase/nuclease, reinforcing that the deletion of 3’NT/NU cannot be compensated by such path. Lastly, a query in TriTrypDB (Kinetoplastid Informatics Resources, https://tritrypdb.org/tritrypdb/app) reveals that only an extra copy of 3’NT/NU is annotated on chromosome 12, supporting the hypothesis of biological consequence associated to the full deletion of the two copies in chromosome 31. Indeed, the Del strains exhibited reduced/absent 3’NT/NU activity in relation to the NonDel and HTZ strains11 indicating that the expression of this enzyme is dependent on the genes deleted. Combined, these facts suggest the existence of relevant differences among Del and NonDel parasites potentially linked to the 3’NT/NU. If such variability is true, distinct hypothesis can be elaborated and should be tested. These will be addressed in the next topics.
** Leishmania’s 3’NT/NU and the modulation of extracellular adenosine (eADO)**
Leishmania’s 3’NT/NU contributes to generate extracellular adenosine (eADO).26 This trait ― together with the host’s ectonucleotidases ―, potentially impact purinergic signalling pathway in the vertebrate by the increase of extracellular level of purines.54 ^,^ 72 The host’s ectonucleotidases CD39 and CD73 are highly expressed in situations of injury, stress and infection.73 When the cell is under one of these conditions, ATP can be released into the extracellular environment as a signal of damage, alerting the host’s immune system.74 This extracellular ATP activates receptors of the P1-type purinergic signalling pathway,72 triggering an inflammatory response characterised by the activation of macrophages, dendritic cells, and the secretion of pro-inflammatory cytokines.75 In this way, CD39 and CD73 are expressed in cell membranes to regulate extracellular levels of ATP, hydrolysing it into adenosine (ADO).54 Certain Leishmania species, including L. infantum, can subvert the host’s inflammatory response by inducing the cell to produce and release more ATP into the extracellular environment, positively regulating the expression of CD39 and CD73 and generating sustained levels of ADO.76 ^,^ 77 ^,^ 78 Extracellular ADO will activate two G protein-coupled adenosine receptors, A2AR (high affinity) and A2BR (low affinity), culminating in the inhibition of the secretion of pro-inflammatory cytokines, inhibition of nitric oxide (NO) production and the microbicidal effects of macrophages, generating an anti-inflammatory milieu conducive to parasite survival.77 ^,^ 79 ^,^ 80 The activation of A2AR receptors also negatively regulates the migration and activation of neutrophils that are induced by Th1 cells, impairing their influx. The absence of A2AR receptor leads to a strong Th1 response and a decrease in spleen and liver tissue parasitism, suggesting that this receptor and its respective activation are important for the establishment of visceral infection by Leishmania.76 This gives us a glimpse of the importance of eADO generation and its role in the success of infection by this parasite. Thus, two main points must be highlighted in this regard: 1) ADO is a signalling molecule that directly interferes with the host’s immune response and influences the survival and visceralisation of Leishmania infantum; 2) Leishmania contributes to modulate the extracellular levels of ADO through the hydrolysis of 3’-AMP by its 3’-NT/NU enzymes;22 ^,^ 26 3) Ultimately, Del and NonDel L. infantum strains exhibiting distinct 3’-NT/NU activity11 and thus, variable abilities to generate eADO might consequently express distinct virulence.
The biological effect of eADO on Leishmania is not restricted to the vertebrate host. Serafim et al.81 demonstrate that ADO restriction triggers Leishmania differentiation into infective metacyclic forms within the sandfly and in vitro. Cultures of promastigotes treated with an antagonist of mammalian adenosine receptors, CGS 15943 (also capable of inhibiting adenosine uptake in Leishmania), presented higher metacyclogenesis. When CGS-treated promastigotes were cultured in medium poor in nutrients, the parasites were not able to proliferate, and metacyclogenesis was triggered; the events were reversed by the addition of adenosine. They also confirmed in experiments with Lu. longipalpis, that infected insects fed with ADO and sucrose presented reduced number of metacyclic. These findings confirmed that ADO is an essential nutrient for the multiplication of parasites, and its absence triggers Leishmania differentiation even in the in-vertebrate host. Moreover, it is reported that, Lu. longipalpis and Phlebotomus (vectors in the OW) differ, among other aspect, in the constituents of their saliva. ADO and AMP were present in the salivary extract of Phlebotomus papatasi e Phlebotomus duboscqi, but, curiously, not in Lu. longipalpis.82 Exporting the reports from literature to the current scenario of Del and NonDel circulation, we raise the question whether the fitness of these strains differ in intra vectorial stage and thus affect the frequency and distribution of these genotypes in Brazil.
3’NT/NU association to miltefosine susceptibility
The association described between infection by Del parasites and patients presenting miltefosine treatment failure20 was further explored by the same authors using CRISPR Cas9.83 The connection between phenotype / genotype was validated by the addback parasites and, importantly, authors identified the genes within the deleted site (MSL) responsible to the miltefosine susceptibility. Knockout cell lines were generated by CRISPR Cas9, for each of the genes deleted in MSL: ∆nuc1 (NUC1 - deletion of the 3’-nucleotidase/nuclease), ∆nuc2 (NUC2 - deletion of the 3’-nucleotidase/nuclease precursor), ∆hlp (HLP - deletion of the helicase-like protein) and ∆tei (TEI - deletion of 3,2-trans-enoyl-CoA isomerase), for both deleted 3’-nucleotidase/nuclease genes, ∆nuc1/nuc2, and for deletion of the entire MSL locus, ∆msl. Using these strains, the authors evaluated different parameters related to the resistance of Del strains (MSL^-^) to miltefosine previously described20 and concluded that isogenic cell lines with the complete deletion ∆msl or with the 3’NT/NU genes deleted ∆nuc1/ nuc2, showed a significant reduction in sensibility to miltefosine, in both amastigote and promastigote forms. It was also observed that these same mutants showed a better ability to control the disturbances caused by miltefosine in the lipidome, reinforcing that 3’NT/NU is a component crucial among the deleted genes for the manifestation of the miltefosine-susceptible phenotype.
Remarks and open questions
The study of the deletion-carrying strains described in Carnielli et al.20 and Schwabl et al.,11 represent a unique opportunity to advance the understanding of adaptive and evolutionary molecular strategies in Leishmania. The biological effects and compensatory responses at genomic, post-transcriptional and translational levels likely contributed to fitness gain for the parasite, which in turn may lead to key variation of epidemiological outcomes. As an example, is the association between Del parasites, (more precisely, lacking 3’NU/NT) and the reduced susceptibility to miltefosine. The drug, although not used in Brazil to treat visceral leishmaniasis in human, was recently approved to treat infected dogs. As the main urban reservoir, a treated dog infected with the not-susceptible Del genotype would continue to harbour significant parasite load, and thus remain a source of infection for the vector. Another major point to address is why Del parasites are widely dispersed and more frequent. Both vertebrate and invertebrate hosts must be considered as major players in this scenario. For vertebrate host, it is plausible to expect reduced virulence during infection by Del strains due to the nature of the deleted genes, especially 3’NU/NT, associated with parasite viability and infectivity. The reduced CNV for Amastin in Del samples (genes also relevant for infection), additionally supports the idea of altered infectivity in these strains. A reasonable hypothesis, thus, is that dogs infected by Del L. infantum progress as asymptomatic, remaining undetected by surveillance services, ultimately contributing to keep the circulation, and spread of these parasites. For the invertebrate, it is important to cogitate coevolutionary interactions between parasite and phlebotomine genotypes that may have shaped variable transmission cycles. Differences in saliva content, for instance, which includes presence of ADO, AMP and ectonucleotidases among vector species, may act as important factors for the transmissibility of L. infantum Del and NonDel. By addressing the presented open questions associated to the cocirculation of the distinct genotypes of L. infantum in Brazil, researchers will generate valuable data on adaptive molecular mechanisms and drug susceptibility of Leishmania, the parasite-hosts interaction, and, ultimately, transmission control.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1WHO Leishmaniasis 2023 https://www.who.int/health-topics/leishmaniasis#tab=tab_1
- 2WHO Number of cases of cutaneous leishmaniasis reported Data by country 2023 https://apps.who.int/gho/data/node.main.NTDLEISHCNUM?lang=en
- 3WHO Number of cases of visceral leishmaniasis reported data by country 2023 https://apps.who.int/gho/data/node.main.NTDLEISHVNUM?lang=en
- 4Akhoundi M Kuhls K Cannet A Votýpka J.Marty P.Delaunay P A historical overview of the classification, evolution, and dispersion of Leishmania parasites and sandflies P Lo S Negl Trop Dis 2016103 e 00043492693764410.1371/journal.pntd.0004349 PMC 4777430 · doi ↗ · pubmed ↗
- 5Steverding D The history of leishmaniasis Parasit Vectors 201710182822820204410.1186/s 13071-017-2028-5PMC 5312593 · doi ↗ · pubmed ↗
- 6Altamirano-Enciso AJ Marquez MG Ferreira MC Rocha-Santos AA Borja-Flores MB Sobre a origem e dispersão das leishmanioses cutânea e mucosa com base em fontes históricas pré e pós-colombianas Hist Cienc Saude-Manguinhos 200310385388314994709 · pubmed ↗
- 7Momen H Cupolillo E Speculations on the origin and evolution of the genus Leishmania Mem Inst Oswaldo Cruz 20009545835881090441910.1590/s 0074-02762000000400023 · doi ↗ · pubmed ↗
- 8Momen H Cupolillo E Grimaldi JG Molecular evidence for the importation of Old World Leishmania into the Americas Biol Res 19932632492557545502 · pubmed ↗