Detection of Virulence Determinants of Uropathogenic Escherichia coli
Nivetha R M, Shanthi Mariappan, Uma Sekar, Aishwarya K.V.L

TL;DR
This study identifies common virulence genes in uropathogenic E. coli and examines their antibiotic resistance patterns.
Contribution
The study provides insights into the prevalence of specific virulence genes and their association with antibiotic resistance in clinical E. coli isolates.
Findings
The fim H gene was the most commonly detected virulence determinant in uropathogenic E. coli isolates.
High resistance rates were observed for ampicillin, ciprofloxacin, and cotrimoxazole among the isolates.
The sat gene was significantly more frequent in isolates resistant to gentamicin, ampicillin, cefotaxime, and cotrimoxazole.
Abstract
Background Escherichia coli is the major causative agent of urinary tract infections (UTIs). Emergence and spread of multidrug-resistant strains of E. coli have raised considerable interest in understanding their diversity and epidemiology of infections in humans. Knowledge of virulence factors of E. coli responsible for pathogenesis of UTIs and their antibiotic susceptibility pattern will help in better understanding of the treatment of UTI. This study was undertaken to characterize virulence determinants of uropathogenic E. coli (UPEC) by genotypic methods and to determine their susceptibility to various classes of antibiotics. Materials and methods A total of 150 clinically significant, non-repetitive, consecutive E. coli isolated from urine were included in this study. Polymerase chain reaction (PCR) was done to detect the presence of virulence genes namely fim H, iut A, hly A,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
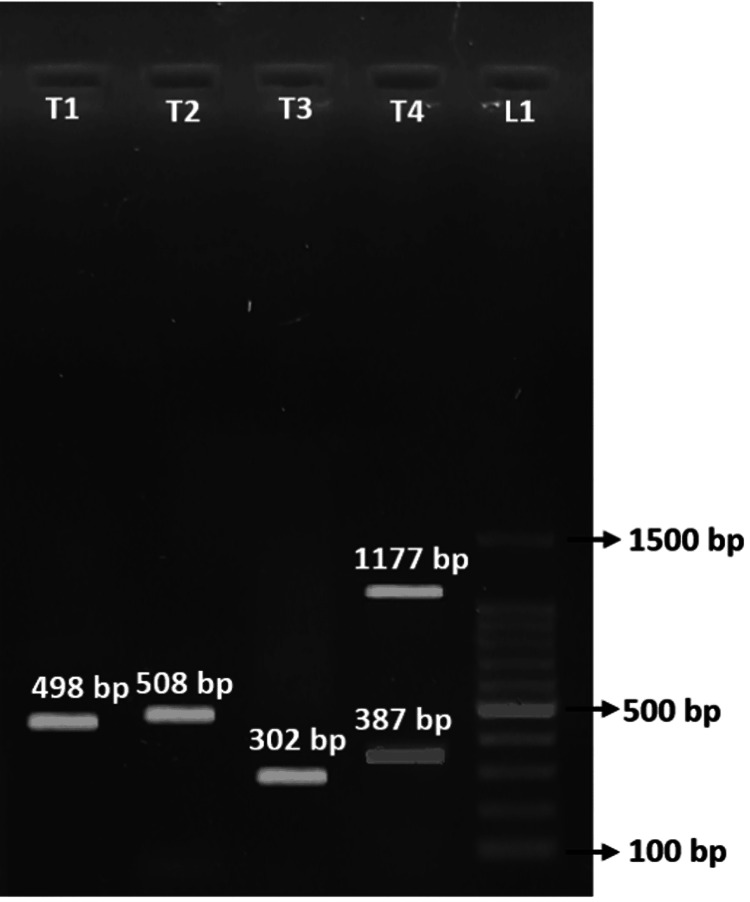 Figure 1
Figure 1| Virulence genes | Primers (5’- 3’) | Annealing temperature (degree celsius) | Amplicon size (base pair) |
| hly A | F: AACAAGGATAAGCACTGTTCTGGCT R: ACCATATAAGCGGTCATTCCCGTCA | 63 | 1177 |
| fim H | F: TGCAGAACGGATAAGCCGTGG R: GCAGTCACCTGCCCTCCGGTA | 63 | 508 |
| pap C | F: GTGGCAGTATGAGTAATGACCGTTA R: ATATCCTTTCTGCAGGGATGCAATA | 63 | 200 |
| iut A | F: GGCTGGACATCATGGGAACTGG R: CGTCGGGAACGGGTAGAATCG | 63 | 302 |
| cnf 1 | F: AAGATGGAGTTTCCTATGCAGGAG R: CATTCAGAGTCCTGCCCTCATTATT | 63 | 498 |
| sat | F: ACTGGCGGACTCATGCTGT R: AACCCTGTAAGAAGACTGAGC | 55 | 387 |
| Genes | Prevalence (Total = 150) (%) |
| fim H | 86 (76.78) |
| Iut A | 64 (57.14) |
| cnf 1 | 29 (25.89) |
| hly A | 2 (1.78) |
| sat | 50 (44.64) |
| pap C | 0 |
| Genes | Base pair |
| fim H | 508 bp |
| cnf 1 | 498 bp |
| hly A | 1177bp |
| Iut A | 302bp |
| sat | 387bp |
| Antibiotics | Virulence genes | ||||||
| fim H (86) | iut A (64) | hyl A (2) | pap C | cnf-1 (29) | sat (50) | ||
| Amikacin | Sensitive | 74 | 59 | 2 | - | 27 | 41 |
| Resistant | 12 | 5 | 0 | - | 2 | 9 | |
| P-value | 0.5836 | 0.1309 | - | - | 0.3091 | 0.1704 | |
| Gentamicin | Sensitive | 69 | 53 | 2 | - | 24 | 35 |
| Resistant | 17 | 11 | 0 | - | 5 | 15 | |
| P-value | 0.6886 | 0.6886 | - | - | 0.8265 | 0.0139 | |
| Fosfomycin | Sensitive | 85 | 63 | 2 | - | 29 | 48 |
| Resistant | 1 | 1 | 0 | - | 0 | 2 | |
| P-value | 0.4145 | 0.7429 | - | - | 0.7159 | 0.2521 | |
| Ampicillin | Sensitive | 22 | 6 | 0 | - | 4 | 4 |
| Resistant | 74 | 58 | 2 | - | 25 | 46 | |
| P-value | 0.0809 | 0.0153 | - | - | 0.4560 | 0.0240 | |
| Cefotaxime | Sensitive | 23 | 18 | 0 | - | 8 | 9 |
| Resistant | 63 | 46 | 2 | - | 21 | 41 | |
| P-value | 0.1617 | 0.4654 | - | - | 0.6286 | 0.0173 | |
| Ciprofloxacin | Sensitive | 12 | 5 | 0 | - | 6 | 4 |
| Resistant | 74 | 59 | 2 | - | 23 | 46 | |
| P-value | 0.5872 | 0.0336 | - | - | 0.3758 | 0.0872 | |
| Cotrimoxazole | Sensitive | 30 | 15 | 2 | - | 10 | 13 |
| Resistant | 56 | 49 | 0 | - | 19 | 37 | |
| P-value | 0.3626 | 0.0019 | - | - | 0.6642 | 0.0343 | |
| Nitrofurantoin | Sensitive | 81 | 61 | 2 | - | 28 | 42 |
| Resistant | 5 | 3 | 0 | - | 1 | 8 | |
| P-value | 0.4121 | 0.2925 | - | - | 0.3872 | 0.3987 | |
| Piperacillin/Tazobactam | Sensitive | 67 | 56 | 2 | - | 24 | 38 |
| Resistant | 19 | 8 | 0 | - | 5 | 12 | |
| P-value | 0.7924 | 0.0261 | - | - | 0.5505 | 0.5734 | |
| Imipenem | Sensitive | 75 | 58 | 2 | - | 26 | 42 |
| Resistant | 11 | 6 | 0 | - | 3 | 8 | |
| P-value | 0.7300 | 0.3962 | - | - | 0.7604 | 0.2904 | |
| Virulence genes (+) | Virulence genes (-) | P-value | |
| ESBL producer | 79 | 24 | 0.5190 |
| Non-ESBL producer | 33 | 14 |
| Virulence genes (+) | Virulence genes (-) | P-value | |
| CRE | 14 | 4 | 0.97234 |
| Non-CRE | 98 | 34 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEscherichia coli research studies · Vibrio bacteria research studies · Urinary Tract Infections Management
Introduction
Urinary tract infections (UTIs) are one of the common bacterial illnesses that affect humans. According to the location of the infection, UTIs are categorized generally as pyelonephritis (the kidney) and cystitis (the bladder). The adhesion to host cells, colonization of tissues, and, in some cases, cellular invasion are necessary for the successful establishment of infection by bacterial pathogens. These steps are then followed by intracellular multiplication, dissemination to other tissues, or persistence. Uropathogenic Escherichia coli (UPEC) is the most common cause of UTI. A wide range of virulence factors are linked to UPEC for causing symptomatic UTIs. The bacterial cell surface and secreted virulence factor are the two broad categories into which the virulence factors of* E. coli* can be classified. These virulence factors include siderophores (aerobactin system), fimbrial adhesins (P, type 1, S, and F1C fimbriae), toxins (hemolysin and cytotoxic necrotizing factor), and capsular polysaccharide (group II capsules) [1]. Fimbriae, particularly type 1 fimbriae and P fimbriae are among the most frequent virulence factors found on bacterial cell surfaces. These fimbriae support adhesion to the host cell surface, cytokine induction, biofilm formation, and tissue invasion [2]. Adhesins aid in the adhesion of the organism to the surface of the epithelial cell, which allows it to avoid the flushing action that occurs during micturition. In addition to directly activating host and bacterial cell signaling pathways, the adhesins can also facilitate the delivery of other bacterial products to host tissues and encourage bacterial invasion. Type 1 pili or the f*im H *adhesin mediates bacterial attachment to several glycoproteins and non-glycosylated peptide epitopes in the bladder epithelium, which results in bacterial internalization and the formation of intracellular bacterial communities. The pap operon (pilus associated with pyelonephritis) codes for the various structural subunits of the P fimbriae [3,4].
Hemolysin, cytotoxic necrotizing factor type 1 (cnf1), and the secreted autotransporter toxin (sat) are the three types of toxins produced by UPEC. Hemolysin A (hly A), also referred to as the "pore-forming toxin," enters the host cell membrane and causes cell lysis, which makes it easier for iron and other nutrients that are crucial for bacterial growth to be released. Cytotoxicity and invasiveness are the mechanisms by which hemolysin and cytotoxic necrotizing factor act. The erythrocytes are lysed, releasing nutrients and other vitamins that are then accessible to the bacteria. In addition, it releases inflammatory mediators and enzymes that damage the renal proximal tubular epithelium by being cytotoxic to erythrocytes, leukocytes, and renal proximal tubular epithelial cells [4]. E. coli produce siderophores like iut A, which absorb iron from the host and aid in colonization and survival [3]. The sat toxin is a serine protease that belongs to the serine protease autotransporters of Enterobacteriaceae (SPATE), which is primarily found in strains of UPEC and is known for its cytopathic effects on the kidney and bladder. This toxin causes vacuolization in the cytoplasm of the uroepithelial cells [4]. Global data indicate that the pathogens that cause UTIs are becoming more resistant to traditional antimicrobial agents [1]. Even to the more recent and powerful antimicrobial agents, resistance has developed. To assess the severity of the issue and direct the empirical selection of antimicrobial agents to treat infected patients, antimicrobial resistance surveillance is required [1]. This study was undertaken to detect the genes encoding for virulence factors in UPEC and to determine their antimicrobial susceptibility profile.
Materials and methods
Ethical approval
The present study was carried out at a 1600-bedded tertiary care, teaching hospital (Sri Ramachandra Institute of Higher Education and Research, Chennai, India). The methodology was approved by the Institutional Ethics Committee (for Medical PG Students) SRIHER (DU) (CSP-MED/20/OCT/62/112).
Bacterial isolates
The study included 150 clinically significant, non-consecutive, non-duplicate urinary isolates of* E. coli*, collected over a period of one year from November 2020 to October 2021. The isolates were identified by standard biochemical tests or by the automated VITEK-2 system (Vitek-2 GN-card; BioMerieux, Marcy-l'Étoile, France).
Antibiotic susceptibility testing
Susceptibility testing for various antibiotics was done by the Kirby-Bauer disc diffusion method and interpreted in accordance with the Clinical and Laboratory Standards Institute guidelines (CLSI 2021). The antibiotics tested were amikacin (30mcg), gentamicin (10mcg), fosfomycin (200mcg), ampicillin (10 mcg), cefotaxime (30mcg), ciprofloxacin (5 mcg), cotrimoxazole (1.25/23.75), nitrofurantoin (300 mcg), piperacillin-tazobactam (100/10), and imipenem (10mcg). The antibiotic discs were procured from Himedia Laboratories (Mumbai, Maharashtra, India).
Template DNA preparation
A single bacterial colony was inoculated into Luria-Bertani broth (Himedia Laboratories, Mumbai, Maharashtra, India) and incubated at 37°C overnight, and centrifuged at 10,000 rpm for 10 minutes. The supernatant was discarded, and the pellet was resuspended in 250 μL of Millipore water, boiled at 100°C for 10 minutes in a water bath, and centrifuged at 10,000 rpm for 10 minutes. The supernatant is used as the template DNA.
Detection of virulence genes
A total of six virulence genes of E. coli were detected by polymerase chain reaction (PCR). The primers used for different virulence genes, their annealing temperature, and the amplicon size are listed in Table 1.
The master mix for each PCR reaction volume contained 10 pmol of forward and reverse primers (Sigma-Aldrich), 10 mm deoxyribonucleotide triphosphate (Takara), 5 U Taq polymerase (Takara), and 10 buffer with MgCl2 (Takara). Each PCR reaction volume also contained 2 L of the deoxyribonucleic acid (DNA) template added to the master mix.
Amplification of the virulence genes was performed using a thermal cycler (Veriti 96 well; Applied Biosystems) under the following conditions: initial denaturation at 95°C for four minutes, followed by 32 cycles of denaturation at 94°C for 30 seconds, annealing based on the primer employed (Table 1) for 30 seconds with an extension at 72°C for 50 seconds, and a final extension for one cycle at 72°C for five minutes. The PCR product was then run on a 1.5% agarose gel stained with ethidium bromide for detection of the amplified virulence gene fragment.
DNA sequencing
PCR-positive amplicons for each virulence gene detected were purified and sequenced. The sequenced strains for each gene served as positive controls. Sequencing was done by using the Sanger AB13730 XL DNA analyzing instrument (AgriGenome; Kerala). Using the Bioedit sequence programme (product version 7.0.5.3), the nucleotide sequences were aligned, and they were then compared with the basic alignment search tool offered on the National Centre for Biotechnology Information website (www.ncbi.nIm.nih.gov). The nucleotide sequences that were analyzed were submitted to the GenBank and the accession numbers were obtained.
The DNA sequences of these virulence genes were submitted to GenBank, and the following accession numbers were obtained: MZ420493-iut A, MZ501821-sat, MZ198898-fim Hiut MZ460390-cnf 1 and MZ465579- hly A.
The statistical analysis was performed using IBM SPSS Statistics for Windows, Version 16 (Released 2007; IBM Corp., Armonk, New York, United States). To determine the significance of the link between virulence genes and antibiotic resistance, a univariate analysis was conducted. Proportions were compared using the chi-square test and the differences were considered significant if P-value ≤0.05 was considered statistically significant.
Results
A total of 150 isolates were included in this study; of them, 80 (53%) were from female patients and 70 (46%) were from male patients.
Molecular methods
Gene Profile
PCR analysis revealed that of the 150 E. coli included in the study, virulence genes were detected in 112 (74%). Overall, fim H gene was detected in 86 (76.78%) followed by iut A gene in 64 (57.14%), sat gene in 50 (44.64%), cnf gene in 29(25.89%) and hly A gene 2(1.78%). The pap C gene was not detected in any of the isolates depicted in Table 2.
Only a single virulence gene was detected in 44 (29.33%) isolates. Out of 44 the virulence genes that occurred singly were fim H (34), iut A (5), SAT (4) and *cnf-1 *(1). The hyl A gene did not occur solely but it coexisted with other virulence genes like fimH, iutA, sat, and cnf genes.
Gel Electrophoresis of PCR for Detecting Virulence Gene in UPEC
The band at 498bp (T1) represents the presence of cnf 1 gene, the band at 508bp (T2) represents the presence of fim H gene, the band at 302bp (T3) represents the presence of iut A gene, the band at 387bp (T4) represents the presence of sat gene, and the band at 1177bp (T4) represents the presence of hly A gene. L1 is the 100bp ladder (Figure 1, Table 3).
Image of gel electrophoresis of virulence genes detected
Of the 112 isolates that carried virulence genes, 68 (60.71%) carried more than one gene in various combinations. Some combinations of virulence genes were found in single isolates only. Thirty-eight (25.33%) of the study isolates did not anchor any of the virulence genes looked for in this study.
Antimicrobial susceptibility pattern
The Kirby-Bauer disc diffusion method revealed the resistance pattern for the antibiotics as follows: amikacin 19 (12.6%), gentamicin 28 (18.6%), fosfomycin 3 (2%), ampicillin 122 (81.3%), cefotaxime 103 (68.6%), ciprofloxacin 127 (84.6%), cotrimoxazole 93 (62%), nitrofurantoin 11 (7.3%), piperacillin-tazobactam 32 (21.3%), and imipenem 18 (12%).
*iut A *gene occurred more frequently in isolates resistant to ampicillin, ciprofloxacin, cotrimoxazole, and piperacillin-tazobactam. Similarly, the *sat *gene was found in isolates resistant to gentamicin, ampicillin, cefotaxime, and cotrimoxazole. No statistical significance was found between other virulence genes and antimicrobial resistance. This is depicted in Table 4.
Of the 150 isolates, 103 were ESBL producers. Among 112 isolates that carried the virulence genes, 79 were ESBL producers. The occurrence of sat genes and ESBL production was statistically significant. There was no difference in the distribution of other genes fim H, cnf-1, and hly A among ESBL and non-ESBL producers (Table 5).
Of the 150 isolates, 18 were carbapenem-resistant. Among the 112 isolates that carried the virulence genes, 14 were carbapenem-resistant. There was no difference in the distribution of any genes among the CRE and non-CRE (Table 6).
Discussion
A comprehensive understanding of the virulence markers of UPEC strains, particularly in hospitalized patients, is essential to monitor the pathogenicity trends of strains responsible for UTIs. In this study, E. coli isolated from urine samples of 150 inpatients with pyelonephritis, cystitis, and urosepsis were analyzed for the presence of six virulence genes (fim H, iut A, hly A, pap C, cnf 1, and sat) and their associated antibiotic susceptibility profiles.
The present study found that 74.66% of the isolates harbored at least one virulence gene, with *fim H (77%) being the most frequently detected, followed by iut A (57%) and sat (45%). The high prevalence of fim H is significant, as it encodes for type 1 fimbriae, essential for E. coli adhesion to host cells in the urinary tract. fim H *is a critical factor in initiating and establishing UTIs. This aligns with other studies where *fimH *was consistently reported as the most prevalent, in UPEC isolates, regardless of the geographical location or study population. One study in Iran found fim H in 99.2% of UPEC isolates [5]. Another study in Nepal showed that 52.3% of UPEC isolates were positive for fim H, compared to 8.5% in the control group [2]. A study in Mexico found that fimH was present in 86 % of isolates [4]. A meta-analysis of 13 studies also identified fim H as one of the most prevalent virulence factors 75.3% [3]. The consistently high prevalence of fim H across studies further signifies its importance as a key target for developing strategies to combat UTI.
iut A (57%) and sat (45%) were the other prevalent genes in the present study. A Mexican study reported iut A in 54.2% and sat in 26.2% of UPEC isolates. The study examining UPEC and APEC isolates in China found iut A in 83% of UPEC and 90% of APEC isolates, while sat was less prevalent, detected in 46.6% of UPEC isolates [4,6]. The low prevalence of hly A 2% in the present study contrasts with the study by Shah et al., who found hemolysin production in 32% of UPEC isolates, reflecting variability in virulence expression [2]. The hly A gene, encoding the toxin α-hemolysin, also shows a wide range of prevalence across the studies, ranging from 7.4% to 31.25% [4,7].
Notably, the pap C gene that encodes for pyelonephritis-associated pilus, commonly linked to pyelonephritis, was absent in all isolates, differing from findings reported in other geographical regions. Reported rates of occurrence of pap C ranged from 26 to 62%, indicating substantial variability [6,7].
Thus, variation in the virulence gene profile could be attributed to regional differences in UPEC strains, the types of UTIs included in the studies (cystitis, pyelonephritis, urosepsis), or the specific patient populations studied.
The study on antimicrobial resistance revealed varied proportions of resistance among various groups of drugs. Higher resistance rates were observed for ampicillin (81.3%), ciprofloxacin (84.6%), cotrimoxazole (62%), and extended-spectrum cephalosporins (68.6%). One study reported a resistance rate of 74.3% for aminopenicillins among UPEC isolates, highlighting the widespread resistance to this class of antibiotics [3,4].
High resistance to ciprofloxacin (84.6%) and cotrimoxazole (62%) parallels findings by Shah et al., who reported similar resistance levels in UPEC isolates [2]. Many reports have been published on the high rate of resistance to fluoroquinolones, particularly ciprofloxacin, which was an effective treatment option for UTI. One study noted that fluoroquinolone-resistant UPEC represented 31.3% of isolates among hospitalized patients in the US between 2007 and 2010 [8]. A meta-analysis of 13 studies found a pooled resistance rate of 49.4% for quinolones [3]. Resistance to cotrimoxazole, another commonly prescribed antibiotic for UTIs, is also a significant concern. A study in Iran found that 45% of UPEC isolates were resistant to cotrimoxazole [1].
Among the isolates, 68% were ESBL producers. The rise of ESBL-producing UPEC strains is a significant driver of resistance to extended-spectrum cephalosporins. This resistance presents a serious therapeutic challenge, as it limits treatment options and often requires the use of carbapenems, which are reserved for more severe infections. In many studies, the ESBL production among UPEC ranged from 26.9% to 53% [5,7].
The relationship between virulence and antibiotic resistance in UPEC is complex and not fully understood. Some studies suggest that there is a positive correlation between virulence and resistance, while others suggest that there is a negative correlation. A possible explanation for a positive correlation is that virulence factors may help UPEC to survive in the presence of antibiotics. Biofilm formation can protect UPEC from the effects of antibiotics. Additionally, some virulence factors, such as siderophores, can help UPEC acquire iron, which is essential for bacterial growth. This may give UPEC a competitive advantage in the presence of antibiotics, which can limit the availability of iron. In some strains, both the virulence and resistance genes may be located on the same plasmids, resulting in their simultaneous transfer [1,3].
However, some studies have also shown a negative correlation between virulence and antibiotic resistance. One study found that quinolone-resistant UPEC strains were less likely to encode for certain virulence genes, such as sfa, hly, and cnf 1. This suggests that the acquisition of quinolone resistance may come at the cost of reduced virulence [1-3]. It has been reported that the UPEC strains lose hemolytic capacity in subinhibitory concentrations of ciprofloxacin, showing a partial or total loss of the pathogenicity island (PAI) containing the hly and cnf-1 genes. It has been reported that quinolone-resistant UPEC isolates carried virulence factor genes related to their ability to invade the urinary tract. Virulence factors, like hemolysin, aerobactin,* cnf-1,* and sat, are chromosomally encoded in the PAIs, which can be deleted from the chromosome spontaneously. Quinolones can act by increasing the deletion and transposition of DNA regions during the development of quinolone resistance facilitated by exposure to quinolones [3].
In the present study, a significant association between the occurrence of sat genes and ESBL production was observed. A study in Iran found that the presence of papGII, iut A, and PAI markers was significantly associated with ESBL production. Conversely, a study in India found that the presence of multiple virulence genes was significantly higher in non-ESBL strains than in ESBL-producing strain which examined for the hly, pap C, and cnf-1 genes [5,9].
In this study, 18 (12%) of the isolates were CRE. A recent study from Egypt reported a rate of 17 (13.07%), in Turkey and Iran it was 11 (22%) and 7%, respectively [10-12]. In the present study, there was no significant difference observed in the distribution of virulence genes between CRE and non-CRE isolates, indicating that resistance and virulence might have evolved independently. A study by Shah et al. observed no statistically significant association between major virulence genes (fim H, pap C, hly A) and carbapenem resistance in UPEC isolates. This aligns with the present findings, highlighting that while resistance mechanisms such as the production of carbapenemases confer survival advantages against antibiotics, they do not inherently enhance or diminish virulence factor expression.
The findings of this study suggest that virulence factors and resistance mechanisms may not always intersect but can independently influence clinical outcomes.
Limitations
Only few common virulence genes were studied. The limited sample size and the limited number of genes could have hindered the statics in their association with various classes of antibiotics. The antibiotic resistance coding genes could also be looked for which might have brought accurate associations and might have supported the study.
Conclusions
The study reveals that f*im H *is the most prevalent virulence factor among UPEC and occurrence of multiple virulence genes within individual isolates. The study reports a very high prevalence of ESBL producers as well as a concerning carbapenem resistance among isolates. Despite the high resistance rates, the study found no significant association between virulence gene presence and antimicrobial resistance profile. Further investigation is essential to explore the potential relationships between resistance and specific virulence mechanisms among UPEC. This knowledge will be vital for developing effective strategies to combat the growing threat of multidrug-resistant UPEC infections.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1The association of virulence determinants of uropathogenic Escherichia coli with antibiotic resistance Jundishapur J Microbiol Asadi S Kargar M Solhjoo K Najafi A Ghorbani-Dalini S 07201410.5812/jjm.9936 PMC 413864425147722 · doi ↗ · pubmed ↗
- 2Virulence factors of uropathogenic Escherichia coli (UPEC) and correlation with antimicrobial resistance BMC Microbiol Shah C Baral R Bartaula B Shrestha LB 2041920193147701810.1186/s 12866-019-1587-3PMC 6720075 · doi ↗ · pubmed ↗
- 3Virulence factors and antimicrobial resistance of uropathogenic Escherichia coli (UPEC) isolated from urinary tract infections: a systematic review and meta-analysis BMC Infect Dis Bunduki GK Heinz E Phiri VS Noah P Feasey N Musaya J 7532120213434864610.1186/s 12879-021-06435-7PMC 8336361 · doi ↗ · pubmed ↗
- 4Relationship between virulence factors, resistance to antibiotics and phylogenetic groups of uropathogenic Escherichia coli in two locations in Mexico Enferm Infecc Microbiol Clin Miranda-Estrada LI Ruíz-Rosas M Molina-López J Parra-Rojas I González-Villalobos E Castro-Alarcón N 4264333520172704896410.1016/j.eimc.2016.02.021 · doi ↗ · pubmed ↗
- 5Distribution of virulence genes and their association with antimicrobial resistance among uropathogenic Escherichia coli isolates from Iranian patients BMC Infect Dis Malekzadegan Y Khashei R Sedigh Ebrahim-Saraie H Jahanabadi Z 5721820183044210110.1186/s 12879-018-3467-0PMC 6238375 · doi ↗ · pubmed ↗
- 6Comparison of virulence factors and expression of specific genes between uropathogenic Escherichia coli and avian pathogenic E. coli in a murine urinary tract infection model and a chicken challenge model Microbiology (Reading) Zhao L Gao S Huan H 1634164415520091937215410.1099/mic.0.024869-0 · doi ↗ · pubmed ↗
- 7Virulence factors and antimicrobial resistance in uropathogenic Escherichiacoli strains isolated from cystitis and pyelonephritis Turk J Med Sci Ghazvini H Taheri K Edalati E Sedighi M Mirkalantari S 3613674920193076184710.3906/sag-1805-100PMC 7350878 · doi ↗ · pubmed ↗
- 8Uro Pathogenic Escherichia coli (UPEC) infections: virulence factors, bladder responses, antibiotic, and non-antibiotic antimicrobial strategies Front Microbiol Terlizzi ME Gribaudo G Maffei ME 1566820172886107210.3389/fmicb.2017.01566 PMC 5559502 · doi ↗ · pubmed ↗