Cortico-hippocampal networks underpin verbal memory encoding in temporal lobe epilepsy
Giorgio Fiore, Davide Giampiccolo, Fenglai Xiao, Matthias J Koepp, Juan E Iglesias, Sjoerd B Vos, Jane de Tisi, Andrew W McEvoy, Giulio A Bertani, Marco Locatelli, Roisin Finn, Lorenzo Caciagli, Meneka Sidhu, Marian Galovic, Sallie Baxendale, John S Duncan, Anna Miserocchi

TL;DR
This study identifies brain regions linked to verbal memory in epilepsy patients, showing how damage to specific areas affects memory performance.
Contribution
The study reveals a cortico-hippocampal network critical for verbal memory in temporal lobe epilepsy patients.
Findings
Smaller grey matter volumes in prefrontal, temporal, and cingulate cortices correlate with worse verbal learning.
Reduced volumes in left dentate gyrus, CA4, and CA3 hippocampal subfields are associated with impaired verbal memory.
Findings were consistent across the entire cohort and in a subgroup of hippocampal sclerosis patients.
Abstract
Knowledge of the structural underpinnings of human verbal memory is scarce. Understanding the human verbal memory network at a finer anatomical scale will have important clinical implications for the management of patients with verbal memory impairment. In this cross-sectional study, we aimed to assess the contributions of cerebral cortex and hippocampal subfields to verbal memory encoding in temporal lobe epilepsy. We included consecutive patients (n = 84) with radiologically and pathologically defined hippocampal sclerosis (HS) (44 left-sided) and unilateral temporal lobe epilepsy, and healthy volunteers (n = 43) who were comparable regarding age and sex. The morphometric and volumetric measures of cerebral cortex and hippocampal subfields were extracted from high-resolution MRI scans. People included in this study underwent standardized neuropsychological evaluation, including…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
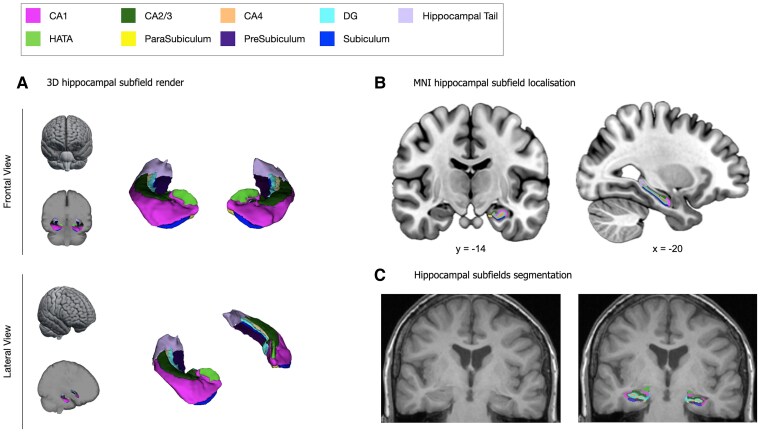 Figure 1
Figure 1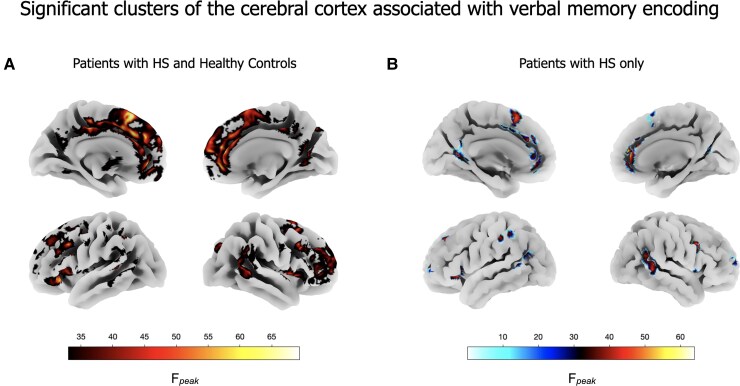 Figure 2
Figure 2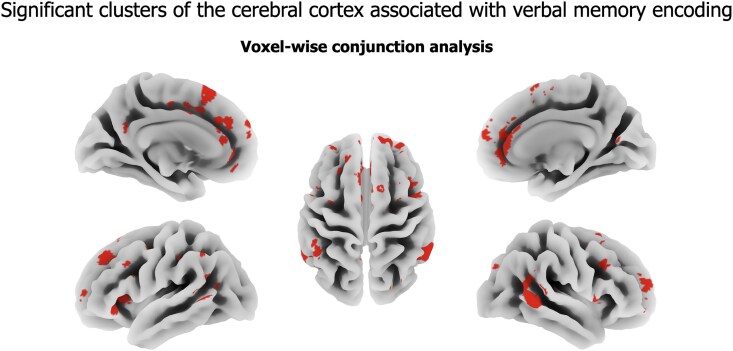 Figure 3
Figure 3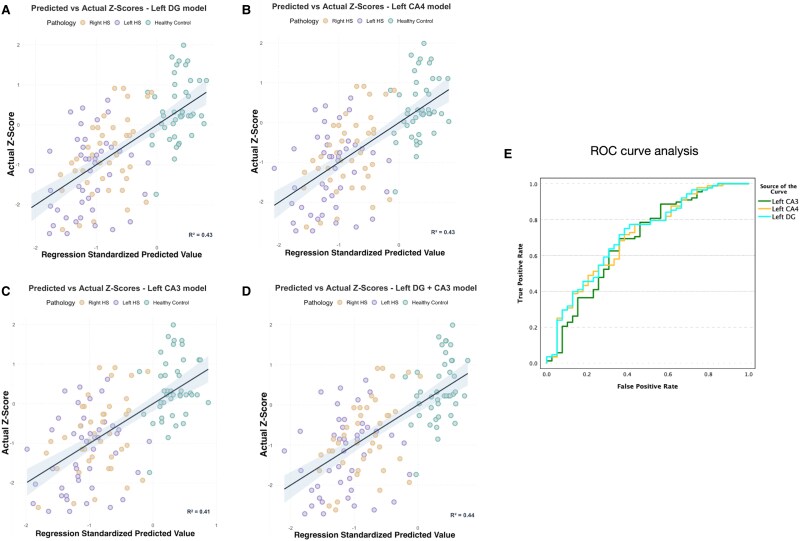 Figure 4
Figure 4| Variable | Left HS ( | Right HS ( | HC ( |
|
|---|---|---|---|---|
| Age (years) | 37.7 ± 11.9 | 38.2 ± 11 | 36.1 ± 11.8 | 0.67 |
| Duration (years) | 24.6 ± 14 | 27.2 ± 13.7 | 0.4 | |
| Sex (female) | 26 (59%) | 26 (65%) | 26 (60.5%) | 0.85 |
| Verbal learning | −1.22 ± 0.9 | −0.8 ± 0.9 | 0.4 ± 0.8 | <0.001 |
| Handedness (left/right) | 11/33 | 4/36 | 7/36 | 0.19 |
| Subfield | Left HS | Right HS | HC |
|
|---|---|---|---|---|
| Left Hippocampus | 1.93 ± 0.36 | 2.75 ± 0.32 | 2.77 ± 0.24 | <0.001 |
| Left DG | 0.21 ± 0.04 | 0.31 ± 0.04 | 0.3 ± 0.03 | <0.001 |
| Left CA4 | 0.18 ± 0.04 | 0.26 ± 0.04 | 0.26 ± 0.03 | <0.001 |
| Left CA2/3 | 0.16 ± 0.04 | 0.23 ± 0.04 | 0.23 ± 0.03 | <0.001 |
| Left CA1 | 0.46 ± 0.09 | 0.65 ± 0.09 | 0.65 ± 0.07 | <0.001 |
| Left subicular C | 0.56 ± 0.1 | 0.78 ± 0.1 | 0.8 ± 0.07 | <0.001 |
| Left tail | 0.32 ± 0.08 | 0.45 ± 0.08 | 0.47 ± 0.08 | <0.001 |
| Left HATA | 0.05 ± 0.01 | 0.07 ± 0.01 | 0.07 ± 0.01 | <0.001 |
| Right hippocampus | 2.92 ± 0.3 | 1.93 ± 0.4 | 2.76 ± 0.27 | <0.001 |
| Right DG | 0.33 ± 0.04 | 0.21 ± 0.05 | 0.31 ± 0.04 | <0.001 |
| Right CA4 | 0.28 ± 0.03 | 0.18 ± 0.04 | 0.26 ± 0.03 | <0.001 |
| Right CA2/3 | 0.25 ± 0.03 | 0.17 ± 0.03 | 0.24 ± 0.03 | <0.001 |
| Right CA1 | 0.69 ± 0.08 | 0.46 ± 0.1 | 0.64 ± 0.08 | <0.001 |
| Right subicular C | 0.81 ± 0.1 | 0.51 ± 0.1 | 0.77 ± 0.08 | <0.001 |
| Right tail | 0.49 ± 0.08 | 0.32 ± 0.08 | 0.49 ± 0.07 | <0.001 |
| Right HATA | 0.07 ± 0.01 | 0.05 ± 0.01 | 0.07 ± 0.01 | <0.001 |
- —Epilepsy Research UK10.13039/501100000295
- —NIHR University College London Hospitals Biomedical Research Centre10.13039/501100012317
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeuroscience and Neuropharmacology Research · Epilepsy research and treatment · Memory and Neural Mechanisms
Introduction
The structural basis of human verbal memory is poorly understood.
It is accepted that the medial temporal lobe (MTL) is critically involved in human memory, relying on its ability to store and retrieve information.^1^ Impairment of episodic memory encoding is common in individuals with drug-resistant temporal lobe epilepsy (TLE),^2^ and anterior temporal lobe resection (ATLR) is associated with postoperative verbal memory decline in up to 60% of patients.^3,4^ However, the considerable individual variation in the extent and direction of verbal memory changes after ATLR suggests that there may be variation in the structural and functional networks supporting verbal memory.^5^
Processing verbal material engages multiple areas of the cerebral cortex to perceive, maintain and store information.^6^ The temporal cortex is hypothesized to serve a crucial role as an initial processing unit.^7^ Given the association between verbal learning and the acoustic properties of words,^8-10^ the temporal cortex may be essential for integrating acoustic and phonemic features during the initial stages of verbal memory encoding.^10^ Functional investigations indicate that the posterior temporal cortex, parietal–temporal–occipital junction (PTOj) and ventrolateral prefrontal cortex (vlPFC) are integral to the processing of verbal material during learning tasks,^10-12^ with frontal lobe contributions to memory encoding having recently become better understood.^13^ Neurophysiological and lesion studies have shown that increased activity of the prefrontal cortices is associated with better verbal memory encoding,^6,14,15^ and patients with lesions in these regions have verbal learning impairment.^16^ The cortical regions studied, however, were limited.
Also, the role of hippocampal subfields in verbal memory encoding needs clarification. Functional MRI (fMRI) studies showed that increased functional activation in the left hippocampus relates to better verbal memory preoperative scores,^17^ with lateralization of verbal memory activation being found to be the best predictor of verbal learning change after ATLR.^18^ The low spatial resolution of fMRI, however, does not allow the determination of the contribution of the various hippocampal subfields to verbal memory. Moreover, fMRI analyses can be affected by the coactivation of multiple brain regions, especially in patients with lesions.^17,19^ Pathological studies offer the potential to investigate the impact of the damage of hippocampal subfields on patients’ verbal memory performances.^20-23^ However, a quantitative relationship between the extent of hippocampal subfield degeneration and verbal memory performances is hard to establish with histology. The most common pattern of hippocampal cell loss, ILAE HS Type 1, manifests as predominant loss of neurons and gliosis in CA1 and CA4 subfields.^24,25^ Patterns of hippocampal sclerosis (HS) appear to be related to various clinical aspects of TLE and may have significance for postoperative prognosis of seizure outcome, but not cognitive outcomes.^26,27^
As above, both the cerebral cortex and hippocampus are established contributors to verbal learning. Animal studies suggest the cingulate cortex may serve as a direct pathway linking the hippocampus and these cortical regions.^28,29^ However, the extent to which the cingulate regions contribute to verbal learning remains to be fully elucidated.
Using voxel-based morphometry (VBM)^30^ and volumetric analyses of the hippocampus and its subfields,^31-33^ we aimed to investigate the anatomical basis of verbal memory encoding function and dysfunction in the cerebral cortex and the hippocampus of patients with TLE, in an objective and quantitative way. We hypothesized that grey matter atrophy of cortical regions involved in the perception of verbal material (e.g. temporal cortices) and in memory encoding (e.g. prefrontal cortices), as well as in regions that connect the neocortex and allocortex (e.g. the cingulum), would be associated with poorer verbal learning performance. Since there is evidence of predominantly left hippocampal lateralization for verbal memory,^17,18,34^ and based on human episodic memory models postulating the involvement of dentate gyrus (DG),^7,35,36^ CA4^37,38^ and CA3,^35,36,39^ for learning and memory encoding, we also hypothesized that a reduction of the volumes of the subfields in the left hippocampus will correlate with impairment in verbal learning.
Materials and methods
Study design
This is a cross-sectional study involving morphometric and volumetric measures of cerebral cortex and hippocampal grey matter structures, in relation to human verbal memory encoding. We studied 84 HS patients (44 left-sided) included in our surgical epilepsy programme between January 2007 and November 2014 and 43 healthy control (HC) subjects. The clinical data, fully anonymized, were extracted from prospectively maintained databases at the National Hospital for Neurology and Neurosurgery (NHNN), London. The data included patients’ clinical history, examination, MRI findings, side of epileptogenic zone, number of antiseizure medications (ASMs), pathology^2^ and presurgical neuropsychological scores.^40^ Inclusion criteria for the current investigation were radiologically and pathologically defined evidence of HS, available preoperative scores on verbal memory tests and preoperative high-resolution structural MRI scans. HCs were volunteers with no history of neurological or psychiatric disease. This study was approved by the Health Research Authority Ethics Committee (22/SC/0016) and followed the STROBE guidelines for cross-sectional studies.^41^
Neuropsychological measures
Patients and controls received extensive standardized neuropsychological evaluation, including measures of verbal memory. Since verbal learning has clinical validity in patients with HS,^5^ the list learning task scores from the Adult Memory and Information Processing Battery (AMIPB) were used as measures of verbal memory in the study population. In the verbal learning task, subjects are read a list of 15 words and asked to recall as many as possible. After five trials, the total number of recalled words is recorded, and the percentage represents the score. The raw scores from the verbal learning test were converted to Z-scores using the means and standard deviations published for sample standardization.
MRI acquisition
The participants included in this study underwent structural high-resolution preoperative MRI on the same scanner (3T General Electric Excite HDx) at the Epilepsy Society, Chalfont, UK. The 3D T1 fast spoiled gradient echo scan was acquired with the following parameters: repetition time = 6.6 ms, echo time = 2.8 ms, inversion time = 450 ms, matrix = 256 × 256 × 192, for a voxel size of 0.9375 × 0.9375 × 1.1 mm.
Morphometric analysis
Voxel-based morphometry analyses were performed using the Computational Anatomy Toolbox (CAT) 12 (https://neuro-jena.github.io/cat/) run within Statistical Parametric Mapping (SPM) 12 (Wellcome Centre for Human Neuroimaging, London, UK; https://www.fil.ion.ucl.ac.uk/spm/software/spm12/). Briefly, CAT12 preprocessed and tissue-segmented individuals’ structural images. Then, spatial registration to the MNI 152 template was obtained through Geodesic Shooting. Grey matter differences were assessed through a multiple linear regression model. Verbal memory Z-scores were included as the main explanatory variable. Total intracranial volume (TIV), age, sex, frequency of focal unaware seizures, history of epilepsy, duration of epilepsy and the number of ASMs were used as covariates. A Bonferroni corrected statistical threshold of P < 0.05 was employed. To control for false positive findings and ensure the identification of meaningful cortical regions,^42^ a minimum cluster size of 100 contiguous voxels was applied. All data were quality controlled according to procedures implemented in CAT12 to address scans’ misalignment, misregistration or inaccurate thickness estimation. No anatomical masks were employed.
After analysing the entire cohort of patients with HS and HCs, we conducted a subgroup analysis focusing exclusively on HS patients. To assess the spatial overlap between the regions identified in the full cohort (HS patients and HCs) and the HS-only subgroup, we conducted a voxel-wise conjunction analysis in SPM. First, we obtained thresholded statistical parametric maps (P < 0.05, corrected) from each model. We then produced a binary mask representing significant clusters for each analysis and multiplied these masks to generate a conjunction map. This procedure isolates voxels that survive statistical thresholding in both models simultaneously, ensuring that any reported overlap reflects regions consistently associated with verbal memory, regardless of the inclusion of HCs.
Volumetric analysis
The segmentation of the whole hippocampus was obtained through Hipposeg software (http://niftyweb.cs.ucl.ac.uk/program.php?p=HIPPOSEG) which exploits a multiatlas-based segmentation propagation method using STAPLE (simultaneous truth and performance level estimation). Hipposeg is established for the segmentation variable morphology, including sclerotic hippocampi.^33^ As previously shown, Hipposeg delineates the hippocampus with no more variability than expert human raters. The segmentation masks were reviewed by two independently trained raters (G.F. and M.G.) and misclassifications were corrected as previously described.^43,44^ The estimated hippocampal volumes were corrected for TIV according to a well-established formula.^33^ TIV was extracted using a parcellation algorithm based on Geodesic Information Flows.^45^ Then, the segmentations of the hippocampal subfields were obtained using FreeSurfer by J.E.I. and S.B.V.^32^ The volumes of the following hippocampal grey matter structures were extracted, TIV-corrected, double-checked and included in the volumetric analysis: CA1, CA3, CA4, DG, subicular complex (para-, pre- and subiculum), hippocampus–amygdala-transition-area (HATA) and hippocampal tail (Fig. 1).
Hippocampal subfields. Localization of hippocampal subfields shown in a 3D brain representation (A) and the MNI 152 template (B). The automated segmentation of the hippocampal subfields displayed on a structural T1 MRI coronal slice (C). Both the 3D reconstruction and T1 MRI segmentation of hippocampal subfields are from a patient with HS and verbal memory deficit (AMIPB Z-score: −2 SD). CA, cornu ammonis; DG, dentate gyrus; HATA, hippocampus–amygdala-transition-area; MNI, Montreal Neurological Institute.
Statistical analysis
Frequencies were reported as a percentage and compared by χ^2^ and Fisher exact tests according to sample size. Continuous normally distributed variables were reported as mean ± standard deviation (SD) and compared with Student’s t-test or analysis of variance. Continuous nonparametrically distributed variables were reported as the median and interquartile range (IQR) and compared via the Mann–Whitney U-test and Kruskal–Wallis test. Linear regression models were used to assess the predictive value of hippocampus and hippocampal subfields’ volumes on verbal learning Z-scores. Multiple linear regression models were employed to correct for covariates. The covariates TIV, age, sex, handedness, history, duration of epilepsy, frequency of seizures, presence and side of HS and number of ASM were chosen based on the literature. Multicollinearity was assessed at each step through tolerance and the variance inflation factor (VIF). Residuals were checked for normality. Statistical significance was regarded for P-values <0.05. Bootstrapping with 1000 samples was employed to control and check the stability of the multivariate models. In the case of more hippocampal subfields predicting verbal memory scores, we planned to use backward stepwise regression methods to identify the regression model and subfield(s) with the highest predictive value. Receiver operating characteristic (ROC) analyses were used to inspect the diagnostic value of the significant subfields in identifying patients with HS and verbal memory impairment. All statistical analyses were performed using IBM SPSS (version 28.0, International Business Machines Corp, New York, USA) and R software 4.2.2 (R Foundation for Statistical Computing, Vienna, Austria; http://www.r-project.org/index.html).
Results
The study included 127 subjects. The clinical and demographical features of the participants are summarized in Table 1. The control population was well matched for age (P = 0.67) and sex (P = 0.85) to the HS population.
Voxel-based morphometry
Grey matter volume reduction of the left and right medial prefrontal cortex (mPFC) and anterior cingulate cortex (ACC) (cluster size = 30 676, Pcorr < 0.0001, Fpeak = 71), right dorsolateral prefrontal cortex (dlPFC; cluster size = 1675, Pcorr < 0.0001, Fpeak = 56), superior and middle temporal gyri (STG and MTG; right: cluster size = 2040, Pcorr <0.0001, Fpeak = 49; left: cluster size = 845, Pcorr < 0.0001, Fpeak = 45), posterior cingulate cortices (PCC; cluster size = 1217, Pcorr < 0.0001, Fpeak = 48) and the left dlPFC and ventrolateral PFC (vlPFC; cluster size = 30 676, Pcorr < 0.0001, Fpeak = 67) and PTOj (cluster size = 606, Pcorr < 0.0001, Fpeak = 58) were associated with worse verbal learning performances in patients with HS and HCs (Fig. 2A).
Significant clusters of the cerebral cortex associated with verbal memory encoding. Cortical representation of brain cortex clusters where greater grey matter volume was associated with better verbal memory performance in the entire (N = 127) cohort (A), including patients with HS and HCs, as well as in the subgroup analysis of HS patients (N = 84) alone (B). Multiple linear regression models were used, and an F-score thresholded map of the significant clusters (P < 0.05, Bonferroni corrected) is shown. The regression model that included patients with HS and HCs was constructed as follows: grey matter volume = β0 + β1⋅(verbal memory Z-score) + β2⋅(age) + β3⋅(sex) + β4⋅(frequency of focal seizures by year) + β5⋅(duration of epilepsy in years)+ β6⋅(number of ASMs) + β7⋅(TIV) + β8⋅(patient or HC group) + ɛ, where β0 is the intercept, β1 is the variable of interest’s coefficient, βn represents the coefficients for the covariates and ɛ is the error. For the model including only HS patients, the same formula was used, but without the categorical variable for group (patient or HC), since all participants in that model were HS patients.
In the subgroup analysis focusing exclusively on HS patients (Fig. 2B), we found that the grey matter volume reduction of the following cortical regions was significantly associated with worse verbal learning performance: grey matter volume of the left and right mPFC and ACC (cluster size = 1681, Pcorr < 0.0001, Fpeak = 65), dlPFC (left: cluster size = 273, Pcorr < 0.0001, Fpeak = 45; right: cluster size = 388, Pcorr < 0.0001, Fpeak = 53), STG and MTG (right: cluster size = 900, Pcorr < 0.0001, Fpeak = 59; left: cluster size = 118, Pcorr < 0.0001, Fpeak = 41), PCC (cluster size = 503, Pcorr < 0.0001, Fpeak = 46) and the left vlPFC (cluster size = 1223, Pcorr < 0.0001, Fpeak = 53) and PTOj (cluster size = 209, Pcorr < 0.0001, Fpeak = 45).
The conjunction analysis revealed a substantial degree of spatial concordance in the significant clusters identified in the entire cohort and the HS-only subgroup, with reduced cluster sizes in the latter (Fig. 3).
Voxel-wise conjunction analysis. Conjunction map illustrating the overlap of cortical regions significantly associated with verbal memory performance in both VBM analyses of the full cohort (N = 127; HS patients and HCs) and the subgroup restricted to HS-only patients (N = 84). Multiple linear regression models were run as described in Fig. 2. The resulting statistical parametric maps were thresholded at P < 0.05 (Bonferroni-corrected), with a minimum cluster size of 100 contiguous voxels applied. Clusters surviving statistical thresholding in both analyses are displayed.
Volumetric study
The volumetric measures are available in Table 2. After correction for TIV, handedness, age, sex, history, duration of epilepsy, frequency of seizures, presence and side of HS and number of ASM, the right hippocampal volume was not associated with verbal learning Z-scores. In the whole cohort, the left hippocampal volume was correlated with verbal learning performance (multivariate model R^2^ = 0.42; b = 0.47; SE = 0.2; P = 0.03).
Concerning hippocampal subfields, unique predictors of verbal learning Z-scores were the volumes of the left DG (R^2^ = 0.43; b = 5.2; SE = 1.7; P = 0.003), CA4 (R^2^ = 0.43; b = 5.8; SE = 2; P = 0.005) and CA3 (R^2^ = 0.41; b = 4.5; SE = 2; P = 0.03) (Fig. 4). In contrast, a greater volume of the right hippocampal tail was associated with worse verbal learning (R^2^ = 0.42; b = −2.6; SE = 1; P = 0.01). Using a backward stepwise method, the left DG and CA3 were included in the multiple regression model with the highest predictive power [R^2^ = 0.44; significant predictors: left DG (b = 10; SE = 4; P = 0.017)]. The multivariate backward stepwise method identified the left DG as the most predictive hippocampal subfield on verbal learning *Z-*scores (multivariate model R^2^ = 0.42; b = 0.47; SE = 0.2; P = 0.03).
Verbal learning Z-scores and hippocampal subfield volumes. Prediction of verbal learning performances (Z-scores) using multiregression models including hippocampal subfield volumes in the entire cohort (N = 127). Each dot shows the relationship between the regression standardized predicted value (x-axis) and the actual values (y-axis) of the individual Z-score. The straight line represents the linear regression line, while the shadow on either side indicates the confidence intervals, illustrating the uncertainty around the predicted values. Each dot corresponds to an individual observation, reflecting the alignment between predicted and actual outcomes. The confidence intervals provide insight into the precision of the model’s predictions across the observed range. (A–D) The association between the reduced chance of verbal memory impairment (Z-scores < −1 SD) and volumes of the left dentate gyrus (DG), CA4 and CA3 is described by receiver operating characteristics curves (AUC = 0.712, 0.704 and 0.684, respectively); N = 84 (E).
To investigate the diagnostic value of hippocampal subfield volumes in patients with HS, a ROC curve analysis was conducted. The ROC curve showed that a greater volume of left DG, CA4 and CA3 was associated with a reduced chance of verbal memory impairment (verbal learning Z-score < −1 SD), with a marginally stronger association for the left DG than CA4, and CA3 (AUC = 0.712, 0.704 and 0.684, respectively; Fig. 4).
Discussion
Using a large sample of TLE patients with HS and healthy subjects, we determined that verbal learning relates to the volume of distinct cortical regions and hippocampal subfields. Our results align with the role that these cortical areas and hippocampal structures have in human computational theories for episodic memory, and animal behavioural models. Further, the results support the thesis of a corticohippocampal network underpinning verbal memory encoding in individuals with TLE.
Cortical regions involved in verbal learning
Grey matter volumes of the medial and dorsolateral PFC, STG and MTG, anterior and posterior cingulate of both hemispheres, and the left vlPFC and PTOj were associated with verbal learning performances (Figs 2 and 3); the greater the grey matter volumes, the better was verbal learning. Notably, the VBM analysis revealed consistent findings across both the entire cohort and the subgroup analysis focusing exclusively on patients with HS. As expected, the size of the significant clusters was reduced when the analysis was restricted to patients. This reduction may reflect decreased variability in grey matter volumes and verbal learning performances after excluding the HC group, underscoring the robustness of these findings within the patient population.
Historically, memory formation has been thought of as the direct transfer to the cortex of elements initially encoded by the hippocampus.^46^ There is now evidence that new memories are not simply transferred to the cortex. They are assimilated into cortical networks, called ‘schemas’, with memory encoding relying on the development and updating of these schemas. When a new experience occurs, this usually conflicts with pre-existing associations. The inclusion of new information into pre-existing cortical schemas represents the key mechanism for memory formation and learning.^47^ Prefrontal control of hippocampal encoding was suggested to have a critical role in learning, resolving the conflict between new information and pre-existing schemas.^46-48^ It seems likely that this accounts for the prefrontal cortices’ grey matter volumes associated with verbal learning in our findings.
The prefrontal cortex underwent great development in primates and humans,^49^ meeting the need for new areas to subserve further processing of sensorial information.^7^ Several prefrontal mechanisms contribute to schema integration and formation during learning. The first requirement for new information to be included in an existing schema is to hold the object of attention active and online among thousands of neural attractors in the cortex.^7^ Through a top-down neural control mechanism, the PFC enhances the selection of populations of neurons useful for the various steps of memory encoding, favouring an efficient allocation of neural resources during learning.^50^ Further, the number of active memories that are integrated into a schema is affected by neural processes such as synaptic facilitation of the type found in the PFC.^51^ This is ascribed to the recurrent collateral connections to nearby neurons which is a typical feature of PFC architecture.^51^ Organization and strategies are other important mechanisms for dealing with large amounts of verbal information^52^; again, these processes were found to be related to PFC.^15^
The pivotal role of PFC during verbal learning is further supported by animal and human studies. During memory encoding, a striking learning-associated upregulation of genes associated with plasticity was demonstrated in the mPFC of rats,^48^ with pharmacological interventions targeting this area preventing learning.^48^ In a similar way, human investigations demonstrated a maximal increased metabolic activity in the mPFC and dlPFC during the execution of auditory-verbal memory tests similar to those used in this study,^15^ with lesions located in the dlPFC and mPFC being specifically related to verbal learning and encoding deficits in patients with strokes, tumours and haemorrhages.^16^ Further, our results align with findings derived from intracranial electrophysiological recording.^6^ During verbal memory encoding, increased low-band neuronal activity was found in cortical regions of the left vlPFC that was consistent with the hotspot of the left vlPFC grey matter volume related to verbal learning in our study.^6^
The posterior STG and MTG represented the main foci of grey matter volumes of the posterior cortex that were associated with verbal learning in our study. These areas, which overlap the Brodmann areas 41, 42, 21 and 22, represent primary auditory receptive and auditory association cortical areas.^15^ Their role in verbal encoding is attributed to the relationship between the acoustic and phonemic properties of words and verbal learning, with acoustic similarities being an acknowledged factor associated with the learning of words.^8-10^ Moreover, these regions may contribute to the learning of verbal material through their function as a storage component of verbal working memory and their active role in rehearsal, a recognized mechanism used during immediate verbal recall.^12,53^ In both functional and lesion studies, researchers demonstrated that these cortical regions may play a major role in verbal memory performances.^6,15,54^ Increased cerebral metabolism,^15^ and recorded neural activity,^6^ were previously described in these cortical regions during the execution of verbal learning tasks, and lesions involving these cortical hubs related to deficits in verbal learning and encoding.^54^ Interestingly, the left PTOj was also related to verbal learning in our study. This region of the posterior cortex has been suggested to be a short-term memory system for human auditory-verbal memory;^7^ that is, patients with damage to this system cannot repeat a heard string of words.^7^
The anterior and posterior cingulate (retrosplenial) cortex was associated with verbal learning in our study. A possible explanation may lie in the connections that ACC and PCC have with the parahippocampal gyrus (representing a direct route to the hippocampus via the entorhinal cortex), the auditory superior temporal gyrus, the dorsal bank of the superior temporal sulcus and with areas of the mPFC.^28,29^ Interestingly, these regions are also those identified as significant by our morphometric analysis. Therefore, we suggest that the cingulate cortices are of pivotal importance to connect the two systems—cortical and hippocampal—and to allow them to efficiently work together, as illustrated by evidence that compensatory network shifts involving the cingulate in response to hippocampal dysfunction.^55-57^ This hypothesis is also corroborated by previous studies showing that GABA cingulate interneuron activity was highly involved in synchronizing network activity and establishing functional oscillations that are associated with information processing and learning.^58^ Lesions in both the ACC and PCC were related to deficits of verbal learning,^16,59^ with functional studies further supporting the role of ACC and PCC in encoding and learning of verbal material.^15,60^
Hippocampal subfields involved in verbal learning
We determined that verbal learning is related to the volumes of the left DG, CA4 and CA3 hippocampal subfields. Patients with reduced volume of the left DG, CA4 and CA3 had worse verbal learning, with the left DG being the main driver.
The DG has a key role in pattern separation and, consequently, in learning and memory encoding. Briefly, signals from the entorhinal cortex are decorrelated and augmented in the DG, before the inputs are presented to the CA3 associative network.^7,35,39,61,62^ Larger volumes of DG were related to a better lure discrimination index in humans.^63^ Our results identified the DG volume as having the strongest association with verbal learning performance, suggesting its role as the main encoding hippocampal unit in verbal memory. Previous pathological investigations confirmed our theory. Degeneration of DG has been described as the strongest predictor of worse verbal learning performances.^21^ The DG’s encoding role relies on neurogenesis,^22,23,64^ with the suggestion that pattern separation can be facilitated by new granule cells’ formation.^22^ Since neurogenesis is highly influenced by granule-cell density in DG,^23^ further impairment of the learning processes in verbal memory can be explained by the reduced neurogenesis in patients with more atrophic, less granule-cell-populated, DGs.
The anatomical and functional connection between DG and CA3 is another fundamental aspect of learning.^7^ Particularly, the mossy fibre system is necessary for the optimal storage of new information in the CA3 associative network.^7,36^ Thus, a significant relationship between the extent of CA3 atrophy and verbal learning performances was anticipated. The importance of CA3 for verbal learning can reside in the associative role that this subfield exerts on inputs from the DG (via the mossy fibre system) and entorhinal cortex (via the perforant pathway), with CA3 demonstrating pattern separation activity under some circumstances and pattern completion activity under others.^39^ Consistent with this theory, pathological studies showed a significant association between severe verbal learning deficits and the lack of a preserved mossy fibre pathway.^21^ Although the CA3 associative role has been suggested to be more important for recall,^35^ pattern completion (and pattern separation under some circumstances) is still a fundamental aspect of learning. The results of this study are concordant with this hypothesis and the suggestion that CA3 is fundamental in some learning tests.^35^
Reduced volumes of the left CA4 predicted worse verbal learning scores. CA4 contributes to pattern separation and context recognition during learning.^37,38^ Its cells are thought to play a complementary role in pattern separation by preprocessing the inputs that will be transmitted to CA3.^37^ Thus, CA4 can contribute to verbal learning by preprocessing information from the DG and directed to CA3, with reduced CA4 volumes being related to worse verbal learning performances. Our results align with previous pathological data. Worse verbal learning performance was seen in patients with degeneration of CA4 than in patients who showed degeneration of CA1.^20^
Although there is evidence that the left and right hippocampi in humans might work together to process qualitatively different information relevant to visual memory tasks,^65^ it is still debated whether task lateralization exists for verbal memory. Our results support a left hippocampal lateralization for verbal memory with verbal learning impairment being related to reduced volumes of the left DG, CA4 and CA3 subfields.^17,66^ Further, we observed an inverse relationship between verbal learning and right hippocampal volumes, with the increased right hippocampal tail volume being related to worse verbal learning Z-scores. In a recent longitudinal study, preserved preoperative network and function before surgery facilitated memory adequacy postsurgery, with memory adequacy relying on the ipsilateral connectivity of structures situated close to the resected areas.^57^ While for other types of memory, such as associative memory,^67^ a compensatory role of the nonaffected contralateral hippocampus was hypothesized, our results concur with the previously proposed functional adequacy model for verbal memory,^34,57^ postulating that the contralateral nonaffected (nondominant) hippocampus is not able to compensate for the affected left-sided (dominant) HS-related verbal memory deficit. These observations are in keeping with the postoperative functional outcomes reported after left ATLR with verbal memory decline in up to 60% of patients,^3,4,34,68^ and a substantial overlap (20% of patients)^69^ between the pre- and postoperative verbal memory scores in patients receiving right ATLR.^70^
Cortical–hippocampal networks underpin human verbal encoding
Our results suggest a cortical–hippocampal network for verbal memory in individuals with TLE. The theory states that prefrontal cortices receive auditory-verbal information from primary and associative auditory cortices of STG and MTG. Via the cingulum, prefrontal cortices and left hippocampus cooperate to assimilate the new information into pre-existing schemas. Within the left hippocampus, the DG and CA4 are fundamental to decorrelate and augment the information through pattern separation, favouring their storage in the CA3 associative network. A schematic representation of our theory is available in the graphical abstract.
The proposed network aligns with the architecture of the involved cortical hubs and the connectivity of white matter tracts. Primary sensory cortical areas have a well-developed granule-cell layer which relies on their role of input first-processing units where representations of the inputs are sparser.^7^ The architecture of the PFC differs from that of sensory cortices in the complexity of the dendritic structure which endows its neurons with higher computational capability, making their recurrent collateral connections crucial to increasing memory capacity and encoding.^7,71^ The proisocortical structure of the cingulum favours forming connectional bridges between the neocortical areas of both hemispheres and the allocortex of the left hippocampus.^72^ Higher-order auditory areas encircle and receive verbal information from the primary auditory cortex. At least two projections emerge from the higher-order auditory areas: one projection is thought to be important for sound localization and it targets the dlPFC; the second projection is involved in processing complex sounds, including linguistic function in humans, and terminates in vlPFC and mPFC.^73^ The arcuate fasciculus has a key role, interconnecting the superior temporal cortex to PTOj and prefrontal cortices.^73^ The cingulum is a collection of axons coursing deep within the cingulate cortices (ACC and PCC) and parahippocampal gyrus. It is the principal pathway carrying information from higher-order sensorial areas in the temporal lobe, prefrontal cortices and PTOj to the hippocampus.^73^ The fornix, via telencephalic and diencephalic structures, connect the cingulate cortices and mPFC to the hippocampus.^73^ This compact fibre bundle is essential for the bidirectional control and effective interplay between mPFC, cingulate cortices and hippocampus which is fundamental for schemas’ integration and update. Finally, to integrate the information of the two halves of the cerebral cortex, tracts that interconnect the two sides of the brain course through the corpus callosum.^73^
Limitations and future prospects
Our study has several strengths, including the use of one of the largest series of HCs and individuals with TLE and HS, the employment of normalized neuropsychological data derived from standardized neuropsychological testing, high-resolution MRI data acquired on the same scanner, robust methodology (Bonferroni corrected analyses) and the correction of our findings for the main predictable confounders (age, sex, handedness, history and duration of epilepsy, frequency of seizures, presence and side of HS and number of ASM). The human corticohippocampal verbal memory network here proposed is supported by animal models, human lesion and functional studies and anatomical and brain connectivity evidence. However, several limitations must be discussed. With the aim of discovering true associations between verbal learning and grey matter volumes of the cerebral cortex and hippocampal subfields, we restricted our analyses to HCs and individuals with TLE who underwent standardized neuropsychological evaluation and MRI data acquisition on the same scanner. This reduced any potential biases caused by different MRI acquisition protocols and variations related to diverse neuropsychological testing. Validation in future studies is encouraged. Second, given the clinical relevance of verbal learning impairments in patients with HS,^5^ we focused our hypothesis specifically on this cognitive domain. While this targeted approach allowed us to concentrate on a key area of cognitive dysfunction, it limited our ability to assess relationships with other cognitive functions, such as fluency, linguistic phonemic and semantic processing or executive control. Future studies should expand upon these findings to clarify the role of the identified cortical and hippocampal structures in supporting verbal memory within a broader cognitive framework that integrates language, memory processes and executive control. Further, even if widely used and validated, the subfield segmentation method we employed on 3T MRI data is still the best approximation available to investigate hippocampal subfields. Future studies may benefit from using 7T MRI images which will allow better discrimination between the different hippocampal subfields and quantification of their volumes.
Clinical implications
The current study demonstrated that verbal learning, in patients with TLE, is strongly related to the volume of distinct regions of the prefrontal, temporal and cingulate cortices and to the left DG, CA4 and CA3 hippocampal subfields. This finding is important. Firstly, a cortical–hippocampal network for verbal memory may be hypothesized for these patients, providing the basis for a better understanding of human verbal memory, with biomarkers that are important for both clinical and research perspectives. Secondly, this study may provide attractive targets for forthcoming modulating therapies. Up to now, direct stimulation of the lateral temporal cortex has been shown to successfully improve episodic memory encoding.^74^ Future studies targeting the prefrontal cortices and/or left PTOj may lead to interesting results. Thirdly, we and others previously showed that verbal memory MRI activation in the posterior hippocampus is protective against verbal memory decline after language-dominant ATLR, and that sparing the posterior 45% of the speech-dominant hippocampus mitigated the risk of verbal memory decline.^18,66,75^ Future work will analyse the impact of sparing or resecting posterior left DG, CA4 and CA3 on verbal memory.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bakker A, Kirwan CB, Miller M, Stark CEL. Pattern separation in the human hippocampal CA 3 and dentate gyrus. Science. 2008;319(5870):1640–1642.18356518 10.1126/science.1152882 PMC 2829853 · doi ↗ · pubmed ↗
- 2De Tisi J, Bell GS, Peacock JL, et al The long-term outcome of adult epilepsy surgery, patterns of seizure remission, and relapse: A cohort study. Lancet. 2011;378(9800):1388–1395.22000136 10.1016/S 0140-6736(11)60890-8 · doi ↗ · pubmed ↗
- 3Bauman K, Devinsky O, Liu AA. Temporal lobe surgery and memory: Lessons, risks, and opportunities. Epilepsy Behav. 2019;101:106596.31711868 10.1016/j.yebeh.2019.106596 PMC 6885125 · doi ↗ · pubmed ↗
- 4Rausch R, Kraemer S, Pietras CJ, Le M, Vickrey BG, Passaro EA. Early and late cognitive changes following temporal lobe surgery for epilepsy. Neurology. 2003;60(6):951–959.12654959 10.1212/01.wnl.0000048203.23766.a 1 · doi ↗ · pubmed ↗
- 5Baxendale S, Thompson P, Harkness W, Duncan J. Predicting memory decline following epilepsy surgery: A multivariate approach. Epilepsia. 2006;47(11):1887–1894.17116029 10.1111/j.1528-1167.2006.00810.x · doi ↗ · pubmed ↗
- 6Topçu Ç, Marks VS, Saboo KV, et al Hotspot of human verbal memory encoding in the left anterior prefrontal cortex. E Bio Medicine. 2022;82:104135.35785617 10.1016/j.ebiom.2022.104135 PMC 9254338 · doi ↗ · pubmed ↗
- 7Rolls ET . Cerebral cortex. Principles of Operation; 2016.
- 8Baddeley AD . Short-term memory for word sequences as a function of acoustic, semantic and formal similarity. Q J Exp Psychol. 1966;18(4):362–365.5956080 10.1080/14640746608400055 · doi ↗ · pubmed ↗