Consensus furin cleavage sites in the cuticular collagens DPY-17 and SQT-3 are required for Q neuroblast left-right asymmetric migration in Caenorhabditis elegans
Vedant D. Jain, Celeste J. Gormly, Erik A. Lundquist

TL;DR
This study shows that specific cleavage sites in two collagens are essential for the asymmetric migration of Q neuroblasts in C. elegans.
Contribution
The study identifies the functional importance of consensus furin cleavage sites in DPY-17 and SQT-3 for Q neuroblast migration.
Findings
CFCS sites in DPY-17 and SQT-3 are required for their roles in Q neuroblast left-right asymmetric migration.
bli-4 mutants showed weak effects on migration, suggesting redundancy among proprotein convertase isoforms.
Apical secretion of cuticular collagens is necessary for Q neuroblast migration and may provide asymmetric guidance.
Abstract
Previous studies showed that the apically secreted cuticular collagens DPY-17 , SQT-3 , and DPY-14 control the left-right asymmetric migration of the Q neuroblasts in Caenorhabditis. elegans . Furthermore, apical secretion of DPY-17 and SQT-3 require the BLI-4 proprotein convertase of the subtilisin/kexin family and the consensus furin cleavage site (CFCS) in the N-terminus of DPY-17 and SQT-3 . Work here shows that the CFCS sites of DPY-17 and SQT-3 are required for their roles in Q neuroblast migration. bli-4 mutants had only weak effects on Q neuroblast migration, possibly due to redundancy among isoforms. These results suggest that apical secretion of cuticular collagens is required for Q neuroblast migration. These collagens might themselves provide left-right asymmetric guidance information, or might regulate another aspect of Q cell interaction with the cuticle, such as…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
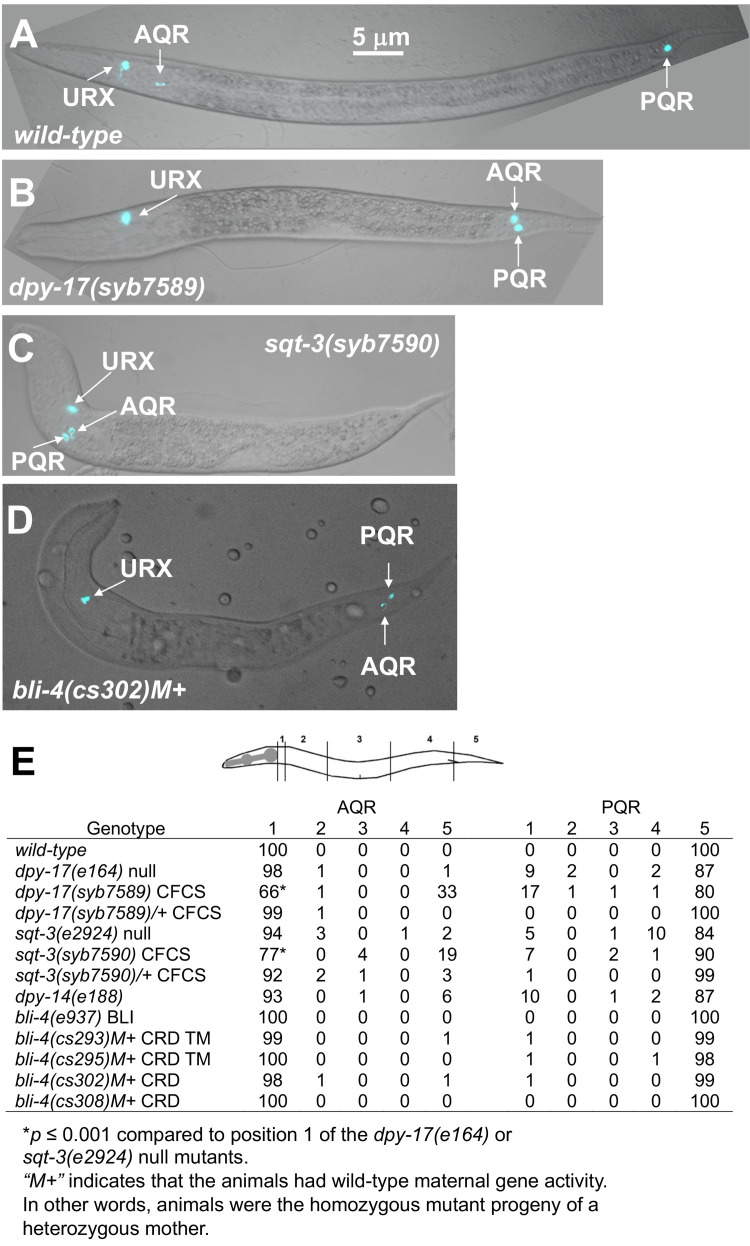 Figure 1
Figure 1|
Strain |
Genotype |
Origin |
|
|
|
CGC |
|
|
|
Josephson et al.,
|
|
|
|
Lang and Lundquist, 2021 |
|
|
|
This work/Birnbaum et al., 2023 |
|
|
|
Lang and Lundquist, 2021 |
|
|
|
This work/Birnbaum et al., 2023 |
|
|
|
Lang and Lundquist, 2021 |
|
|
|
This work/CGC |
|
|
|
This work/Birnbaum et al., 2023 |
|
|
|
This work/Birnbaum et al., 2023 |
|
|
|
This work/Birnbaum et al., 2023 |
|
|
|
This work/Birnbaum et al.
|
|
|
|
This work/Birnbaum et al., 2023 |
- —National Institute of Neurological Disorders and Stroke (United States)https://ror.org/01s5ya894
- —National Institute of General Medical Sciences (United States)https://ror.org/04q48ey07
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
Topics14-3-3 protein interactions
Description
The bilateral Q neuroblasts, sisters of the V5 seam cells are born during embryogenesis and migrate in the L1 larva (Sulston 1976; Chapman
- et al.* 2008; Middelkoop and Korswagen 2014). On the right side, QR migrates anteriorly over the V4 seam cell, and on the left side, QL migrates posteriorly over the V5 seam cell (Chapman
- et al.* 2008). An identical pattern of cell division, cell death, and neuronal differentiation produces three neurons on each side: AQR, AVM, and SDQR on the right, and PQR, PVM, and SDQL on the left. QR descendants migrate anteriorly. AQR migrates the farthest to the deirid ganglion near the pharynx ( Figure 1 ). QL descendants migrate posteriorly. PQR migrating the farthest posteriorly to the phasmid ganglion posterior to the anus ( Figure 1 ).
The transmembrane receptor molecules UNC-40 /DCC, PTP-3 /LAR, MIG-21 , CDH-3 and CDH-4 control initial Q migration (Middelkoop
- et al.* 2012; Sundararajan and Lundquist 2012; Sundararajan
- et al.* 2014; Ebbing
- et al.* 2019). UNC-40 /DCC and PTP-3 /LAR act redundantly in parallel in QL to promote posterior migration, and mutually inhibit each other's activity in QR, causing anterior migration. Initial migration defects cause subsequent errors in migration of AQR and PQR (Chapman
- et al.* 2008).
Mutations in the cuticle collagen genes * dpy-14 , dpy-17 * , and * sqt-3 * cause initial Q migration defects similar to * unc-40 * and * ptp-3 * , as evidenced by reversal of AQR and PQR migration (Lang and Lundquist 2021; Lundquist 2024) . AQR is sometimes in the normal posterior position of PQR, and PQR is sometimes in the normal anterior position of AQR ( Figure 1 ) . DPY-17 and SQT-3 likely act together in a collagen trimer and are mutually required for each other secretion (Novelli
- et al.* 2006; Birnbaum
- et al.* 2023). AQR/PQR migration defects in double mutants of each combination resemble defects in single mutants, suggesting that they all act in a common pathway, possibly in a DPY-14 / DPY-17 / SQT-3 collagen trimer (Lang and Lundquist 2021; Lundquist 2024) . The proprotein convertase of the subtilisin/kexin family (PCSK) BLI-4 is required for proper apical secretion of DPY-17 and SQT-3 to form the cuticle (Birnbaum
- et al.* 2023). Cleavage of DPY-17 and SQT-3 at an N-terminal dibasic consensus furin cleavage site (CFCS) by BLI-4 is required for secretion (Birnbaum
- et al.* 2023). Mutations in
bli-4 * cause improper DPY-17 and SQT-3 secretion and cuticle defects. Mutations of the CFCS sites in * dpy-17 * and * sqt-3 * cause similar defects (Birnbaum
- et al.* 2023). DPY-14 also has a predicted N-terminal CFCS site (…STAGKSGY
R * AK * R * AWQFGSWV…) but its function has not been investigated.
Work described here shows that the CFCS sites of DPY-17 and SQT-3 are required for Q neuroblast migration. * dpy-17 ( syb7589 ) * and * sqt-3 ( syb7590 ) * CFCS mutants displayed AQR and PQR migration defects that resembled null alleles, with AQR sometimes in the tail and PQR sometimes in the head ( Figure 1 ). The penetrance of PQR defects of CFCS mutants were not significantly different than nulls ( Figure 1 ). However, AQR defects were significantly stronger in the CFCS mutants. This suggests that the CFCS mutants could have a dominant interfering effect on other factors necessary for AQR migration, possibly by interfering with their secretion ( *e.g. * another unidentified cuticle collagen that might be involved). Indeed, * sqt-3 ( syb7590 )/+ * heterozygotes displayed dominant Dpy and right-hand roller phenotypes, and dominant AQR and PQR migration defects ( Figure 1 ). * dpy-17 ( syb7589 )/+ * heterozygotes were weakly Dpy but did not display AQR/PQR migration defects. These results indicate that the * sqt-3 ( syb7590 ) * mutant is dominant and might have dominant-negative effects on cuticle formation and on AQR/PQR migration. DPY-17 and SQT-3 are mutually required for each other's secretion (Novelli
- et al.* 2006; Birnbaum
- et al.* 2023). In
dpy-17 * mutants, much SQT-3 remains in the cytoplasm in a pattern consistent with the endoplasmic reticulum (Birnbaum
- et al.* 2023). In
sqt-3 * mutants, DPY-17 protein level is severely reduced (Birnbaum
- et al.* 2023). This suggests that SQT-3 is required for the stability of DPY-17 and possibly other proteins, and might explain the dominant-negative effect of
sqt-3 ( syb7590 ) * but not * dpy-17 ( syb7589 ) * .
The classic * bli-4 ( e937 ) * viable blistered cuticle mutant had no defects in AQR or PQR migration ( Figure 1 ). * bli-4 * encodes at least four groups of isoforms, with viable * bli-4 ( e937 ) * affecting only the BLI isoform group (Birnbaum
- et al.* 2023). Mutations predicted to affect all isoforms ( i.e.
bli-4 * nulls such as * bli-4 ( cs281 * )) are embryonic lethal and could not be scored for AQR/PQR migration, which occurs in the L1 larva. Mutations that specifically affect the cysteine-rich domain (CRD) isoforms with and without a transmembrane domain (TM) result in larvally-arrested animals with defective cuticles (Birnbaum
- et al.* 2023). These mutants had variable defects in AQR and PQR migration (0-2%), less penetrant than
dpy-17 * and * sqt-3 * mutants ( Figure 1 ). Possibly, different isoforms of BLI-4 can act with redundancy on DPY-17 and SQT-3 CFCS cleavage. The astacin metalloprotease DPY-31 is thought to process SQT-3 at a C-terminal site (Novelli
- et al.* 2004; Novelli
- et al.* 2006), but
dpy-31 * mutants had no effect on AQR/PQR migration (Lang and Lundquist 2021) . This is consistent with other results that show that some aspects of SQT-3 function do not require DPY-31 (Novelli
- et al.* 2004; Novelli
- et al.* 2006; Birnbaum
- et al.* 2023).
These results indicate that factors affecting cleavage and apical secretion of DPY-17 and SQT-3 to form the apical extracellular matrix cuticle are required for their roles in Q neuroblast migration. It is unclear how the cuticle might be providing left-right asymmetric guidance information to the Q neuroblasts. Possibly, cuticle structure affects the structure of the underlying basal extracellular matrix basement membrane upon which the Q neuroblasts migrate. EPI-1 /lamininA is required for the ability of the Q descendants to migrate but does not affect direction (Lang and Lundquist 2021) . EMB-9 /Collagen IV A1 mutants have low penetrance PQR directional migration defects (0-2%) (Lang and Lundquist 2021) , suggesting a possibly role of basement membrane collagen IV in QL direction. The Q cells are the daughters of the lateral epidermal seam cells and are in contact with the cuticle when born before they migrate between the basement membrane and epidermis (Sulston and Horvitz 1977; Chapman
- et al.* 2008; Middelkoop and Korswagen 2014). Possibly, DPY-14 , DPY-17 , and SQT-3 in the cuticle provide the left-right asymmetric guidance information required for initial Q neuroblast migration. Alternately, DPY-14 , DPY-17 , and SQT-3 might affect Q cell attachment to the cuticle, which might influence migration. This is consistent with DPY-17 acting with MUA-3 /Fibrillin1 in muscle cell and organ attachment to the cuticle. In any case, these data indicate that collagens in an apical extracellular matrix can influence neuroblast migration.
Methods
Standard * C. elegans * culture and genetic techniques at 20°C were utilized (Brenner 1974; Fay 2013) . A Pgcy-32::cfp transgene ( * lqIs244 ) * was used to visualize AQR and PQR (Chapman
- et al.* 2008; Josephson
- et al.* 2016). A five-position scale was used to assess AQR and PQR position along the body as previously described (Josephson
- et al.*
- (see diagram in Figure 1E ): position 1-the normal position of AQR in the deirid ganglion in the anterior; position 2-posterior to the normal position of AQR and anterior to the vulva; position 3 adjacent to the vulva; position 4-the birthplace of the Q neuroblasts in the posterior; and position 5-the normal final position of PQR in the tail in the phasmid ganglion behind the anus. Significance of difference of proportional data between genotypes was determined using Fisher's Exact test. Wormbase (Sternberg et al., 2024) was used for
C. elegans * informatics.
Reagents
The following * C. elegans * strains and genotypes were utilized:
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Birnbaum Susanna K. Cohen Jennifer D. Belfi Alexandra Murray John I. Adams Jennifer R. G. Chisholm Andrew D. Sundaram Meera V. 2023918 The proprotein convertase BLI-4 promotes collagen secretion prior to assembly of the Caenorhabditis elegans cuticle PLOS Genetics 1991553-7404 e 1010944 e 101094410.1371/journal.pgen.101094437721936 PMC 10538796 · doi ↗ · pubmed ↗
- 2Brenner S 197451 THE GENETICS OF CAENORHABDITIS ELEGANS Genetics 7711943-2631719410.1093/genetics/77.1.714366476 PMC 1213120 · doi ↗ · pubmed ↗
- 3Chapman Jamie O. Li Hua Lundquist Erik A. 2008121 The MIG-15 NIK kinase acts cell-autonomously in neuroblast polarization and migration in C. elegans Developmental Biology 32420012-160624525710.1016/j.ydbio.2008.09.01418840424 PMC 2642615 · doi ↗ · pubmed ↗
- 4Ebbing Annabel Middelkoop Teije C. Betist Marco Bodewes Eduard Korswagen Hendrik C. 201911 Partially overlapping guidance pathways focus the activity of UNC-40/DCC along the anteroposterior axis of polarizing neuroblasts Development 1477-912910.1242/dev.180059 PMC 737676131488562 · doi ↗ · pubmed ↗
- 5Fay David S. 20131230 Classical genetic methods Worm Book 1551-850715810.1895/wormbook.1.165.1PMC 412749224395816 · doi ↗ · pubmed ↗
- 6Josephson Matthew P. Chai Yongping Ou Guangshuo Lundquist Erik A. 2016210 EGL-20/Wnt and MAB-5/Hox Act Sequentially to Inhibit Anterior Migration of Neuroblasts in C. elegans PLOS ONE 1121932-6203 e 0148658 e 014865810.1371/journal.pone.014865826863303 PMC 4749177 · doi ↗ · pubmed ↗
- 7Lang Angelica E. Lundquist Erik A. 2021219 The Collagens DPY-17 and SQT-3 Direct Anterior–Posterior Migration of the Q Neuroblasts in C. elegans Journal of Developmental Biology 912221-37597710.3390/jdb 901000733669899 PMC 8006237 · doi ↗ · pubmed ↗
- 8Lundquist E 2024826 The DPY-14 cuticle collagen regulates left-right asymmetric neuronal migration in Caenorhabditis elegans.Micro Publ Biol 202410.17912/micropub.biology.00130239258272 PMC 11384304 · doi ↗ · pubmed ↗