Mitigating stress: exploring how our feet change shape with size
Paige Treherne, Erin C.S. Lee, Michael J. Rainbow, Luke A. Kelly
TL;DR
This study examines how foot bones change shape with size to reduce joint stress in larger individuals.
Contribution
The paper identifies allometric scaling patterns in foot bones that help mitigate joint contact stress.
Findings
Articular surfaces on the calcaneus scale with positive allometry relative to bone volume.
Total bone surface area scales with negative allometry as bone size increases.
The calcaneus becomes more cube-like with increasing size, potentially reducing internal bone stress.
Abstract
If human skeletal shape increases proportionally with size (isometric scaling), we would expect exponential increases in joint contact stress as individuals become larger. However, if skeletal shape changes as a function of size (allometric scaling), this can mitigate increases in joint contact stress by changing the surface area (SA)-to-volume ratio. Here, we explored whether human foot bones scale with allometry and, if so, to identify the shape features that are associated with bone size. Computed tomography scans of the two largest foot bones (talus, calcaneus) were obtained from 36 healthy individuals. We implemented a scaling analysis for each joint articular surface and bone. We performed a Procrustes ANOVA to establish the shape features associated with bone size. In line with our hypothesis, articular surfaces on the calcaneus scaled with positive allometry relative to bone…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
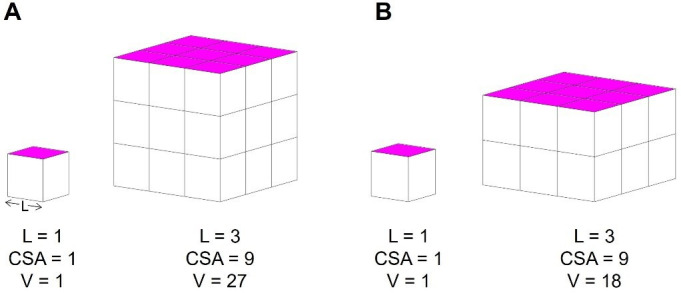 Figure 1
Figure 1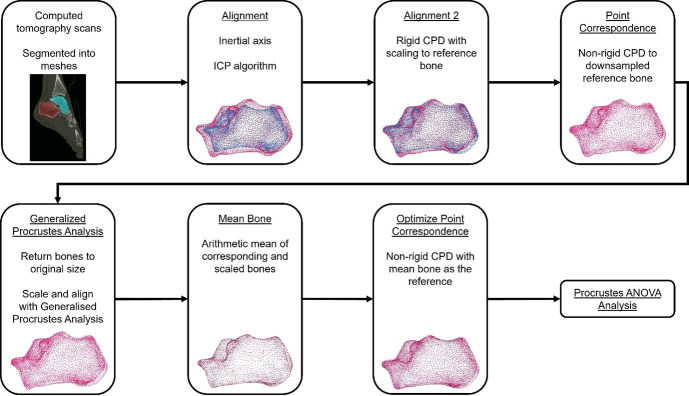 Figure 2
Figure 2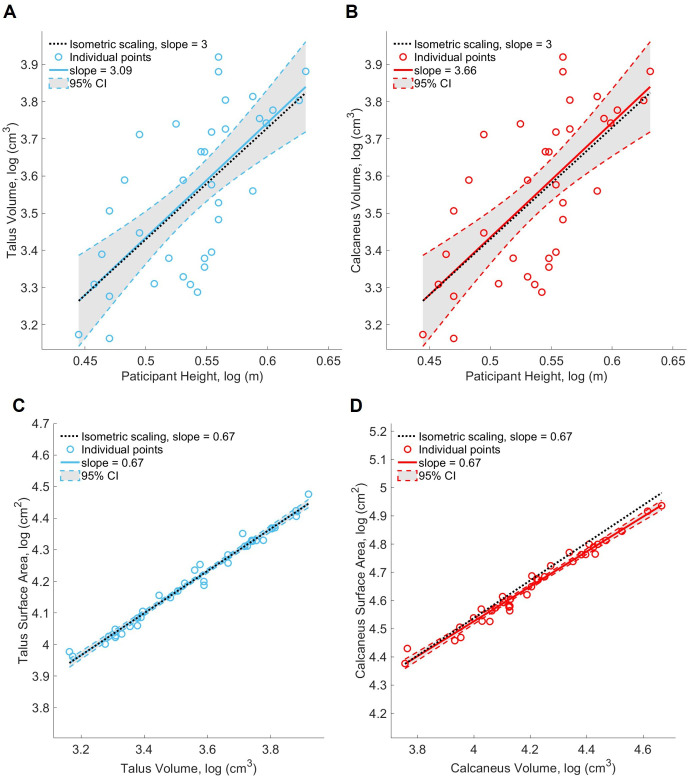 Figure 3
Figure 3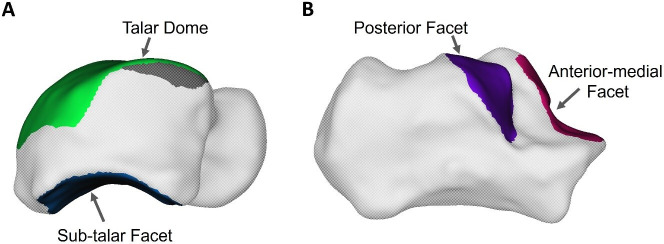 Figure 4
Figure 4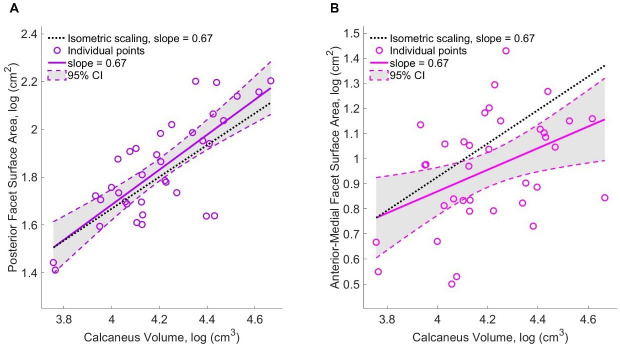 Figure 5
Figure 5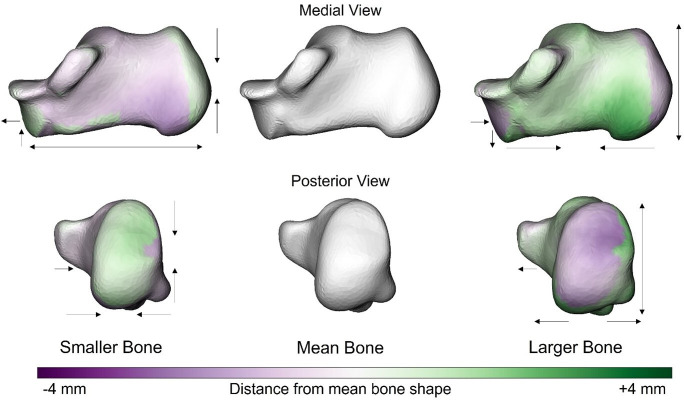 Figure 6
Figure 6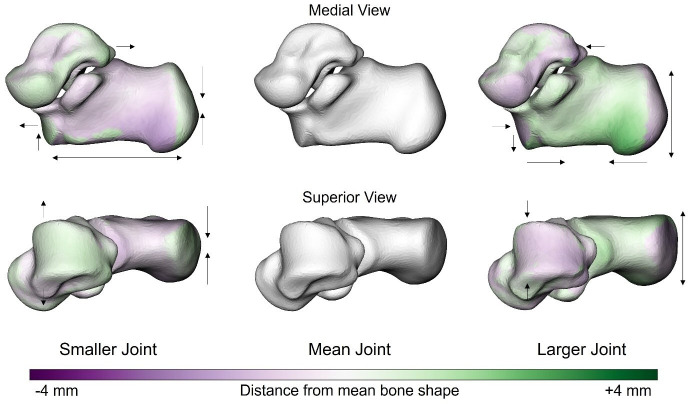 Figure 7
Figure 7|
slope |
slope 95% CI |
|
|
power achieved |
scaling strategy | |
|---|---|---|---|---|---|---|
|
talus |
0.67 |
0.64, 0.70 |
0.87 |
0.98 |
1.0 |
isometry |
|
sub-talar surface |
0.71 |
0.57, 0.85 |
0.55 |
0.77 |
1.0 |
isometry |
|
talar dome |
0.71 |
0.61, 0.81 |
0.46 |
0.85 |
1.0 |
isometry |
|
calcaneus |
0.62 |
0.59, 0.65 |
0.0037 |
0.98 |
1.0 |
negative allometry |
|
posterior facet |
0.73 |
0.52, 0.95 |
0.55 |
0.58 |
0.9999995 |
isometry |
|
anterior-medial facet |
0.43 |
0.11, 0.75 |
0.14 |
0.18 |
0.78 |
isometry |
- —Australian Research Councilhttp://dx.doi.org/10.13039/501100000923
- —Natural Sciences and Engineering Research Council of Canadahttp://dx.doi.org/10.13039/501100000038
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMorphological variations and asymmetry · Foot and Ankle Surgery · Lower Extremity Biomechanics and Pathologies
Introduction
Human feet exhibit a broad range of sizes and shapes [1–3]. But are bigger feet just a scaled-up version of smaller feet? If this were the case, our feet (and foot bones) would scale isometrically, increasing equally in all dimensions for every unit of size increase. Isometric scaling follows the square–cube law, as length (L) increases in all directions, surface area (SA) increases to the power of two and volume (V) increases to the power of three (SA ∝ L^2^, V ∝ L^3^) (figure 1A). However, isometric scaling in terrestrial animals without specific shape or postural adjustments may cause disproportionate increases in joint articular contact pressures because as overall size increases as a function of volume, SA grows at a slower rate [4,5].
(A) Isometric scaling: the square–cube law. As an object increases uniformly in length, its cross-sectional surface area (CSA, pink) increases by the length to the power of two (l2). The volume increases by length to the power of three (l3). Therefore, SA grows at a rate of volume2/3, and (B) Allometric scaling: an object does not grow uniformly in all dimensions, allowing the shape to change as a function of size. Following allometric scaling, the SA can increase faster than isometric scaling (positive allometry).
Allometric scaling describes a shape–function relationship that deviates from the square–cube law (figure 1B). Allometric scaling is suggested as a strategy to offset increased gravitational and muscular forces as terrestrial animals increase in size [4,6]. For example, positive allometry describes a shape function relationship whereby the SA of a skeletal structure grows at a rate faster than would be predicted with the square–cube law for isometric scaling. This strategy has been shown to mitigate increasing joint contact stresses across a range of species of different sizes and within specific regions on the bones of the hindfoot complex [4,6–12].
The human hindfoot complex is unique in shape compared with other non-human primates with specific adaptations to enable habitual upright locomotion [13–15]. Compared to non-human primates, the human calcaneus is larger and more robust, presumably to accommodate increased ground reaction forces and muscular forces applied by the Achilles tendon [15–17]. The human talus is less ‘wedged’ with a flatter talar dome, which is hypothesized to allow for equally distributed load across the surface during locomotion with an upright tibia [18–20]. However, humans exhibit a wide range of sizes; the average heights of adult females around the world range from 150 to 168 cm, while the average heights of adult males range from 160 to 183 cm [21], and average adult body mass (combined female and male) from 58 to 81 kg [22]. Since we still complete similar bipedal movement patterns, it is likely that we may also need to change shape as a function of size to maintain healthy joint contact stress. This could particularly be the case for our feet, which support our entire body’s mass and are our primary interface with the earth we move across. Within the human population, substantial variations in hindfoot bone shape also exist [23–25]. However, it is unknown to what extent these shape differences are related to size.
Therefore, this study aims to explore whether human foot bones scale with allometry and, if so, identify allometry-associated shape features. First, we compared the relationships between bone volume and SA of the calcaneus, talus and their articular surfaces to the relationship expected from isometric scaling. We used geometric morphometrics to create a statistical shape model to assess whether shape features are related to size. We regressed this model against bone and joint volume using a Procrustes ANOVA. Based on McMahon’s hypothesis that animals scale with constant stress similarity [4], we predict that the bones and their articular surfaces will scale with positive allometry (SA growing faster than isometric scaling) and that there will be shape features at an individual bone and joint level related to size.
Methods
Participants
2.1.
After Institutional Review Board approval in accordance with the Declaration of Helsinki and informed consent, 36 healthy adults were included in this study (15 female, 21 male; height 156.0–188.0 cm; mass 54.0–109.0 kg; 19–51 years). Individuals were excluded if they had a history of lower limb injury using a modified version of the Identification of Functional Ankle Instability Questionnaire, as well as any known neurological impairment, musculoskeletal issues or cardiovascular conditions [26].
Computed tomography scans
2.2.
A computed tomography (CT) scan (120 kV, 60 mA, General Electric Medical Systems, USA) of each participant’s right foot was captured with the participant lying prone with their ankle in a plantar-flexed orientation (average resolution: 0.356 × 0.356 × 0.625 mm). A plantar-flexed ankle and foot orientation was chosen as it improves the in-plane resolution of the foot [27]. Talus and calcaneus bones were segmented in Mimics 24.0 (Materialise, Leuven, Belgium) to create three-dimensional bone surface meshes [28,29]. The meshes were then smoothed in Geomagic Wrap 2021 (3D Systems, SC).
Data processing and analysis
2.3.
Size and allometry
2.3.1.
To assess how the talus and calcaneus volume scaled with body size (height), we fit a linear regression model of bone volume on body height in a logarithmic space, using log height – log bone volume plots. Isometric scaling on a log height – log volume plot is represented by a fit line with a slope of 3 (square–cube law, volume = length^3^). To test for allometry, the SA and volume of the talus and calcaneus were computed from the tessellated meshes of each bone and articular surface [30] in Matlab (Mathworks, Natick, MA). To assess whether the talus and calcaneus scale with isometry or allometry, log volume (Log V) – log surface area (Log SA) plots were computed. Isometric scaling (equal growth in all dimensions) on a Log V – Log SA plot is represented by a fit line with a 0.67 slope (square–cube law, SA = V^0.67^) [8,10]. Allometric scaling is found to be a deviation from either side of this line. Joint articular surface scaling was assessed using the same approach, comparing the joint articular SA to the corresponding bone volume.
Joint articular surfaces
2.3.2.
To obtain the joint articular surfaces of interest (talus sub-talar facet, talus talar dome, calcaneus posterior facet and calcaneus anterior-medial facet), each vertex within the anatomical boundaries of these surfaces were manually selected from the smoothed bones in Geomagic Wrap 2021 (3D Systems, SC). The articular surfaces were then isolated, and the boundaries of the surfaces were inspected to ensure they were appropriate.
Statistical shape models
2.3.3.
To quantify the relationship between shape and size across our sample of tali and calcanei, we created a series of statistical shape models (figure 2). These models were created at levels of the individual bones (talus, calcaneus) and at the level of the joint complex (sub-talar joint, combined talus and calcaneus). To initiate the alignment and point correspondence process, an initial reference bone was selected by visually inspecting all bones in the sample and choosing a reference bone that appeared to be close to an average size with an absence of irregular features.
To build a statistical shape model of the sub-talar joint complex, we initially generated three-dimensional surface meshes by segmenting CT scans. A series of alignment and point correspondence processes are undertaken to ensure robust mesh correspondence across all samples, including an iterative closest point (ICP) algorithm and rigid and non-rigid coherent point drift (CPD) processes. We subsequently performed a Procrustes ANOVA to objectively identify bone shape features that are related to size.
Alignment
2.3.3.1.
Initially, all bones were aligned to their inertial axes. Subsequently, bones were further aligned using an iterative closest point algorithm. A rigid coherent point drift (CPD) algorithm was then applied to align each bone in the sample to ensure optimal alignment to the reference bone while scaling the bones using relative centroid sizes (the square root of the sum of squared distances from each mesh vertex to the bone’s centroid) [31]. As the meshes at this point have a different number of vertices, this scaling factor is an initial guess that will be refined in subsequent steps. These rigid transformation steps are undertaken to facilitate optimal point correspondence in the following step without influences of alignment and size impacting the initial point correspondence.
Point correspondence
2.3.3.2.
Initial mesh correspondence was completed using a non-rigid CPD algorithm, reducing the mesh vertices in each bone in the sample to match the reference bone’s number of vertices while conserving individual bone shape [31]. The output bones (all with the same number of vertices) were unscaled using the scaling factor from the rigid CPD transformation to return the bones to their original size. We then performed Generalized Procrustes Analysis on the corresponding bones [32]. This process scales the bones to a common centroid size and further aligns the meshes by minimizing the sum squared distances between corresponding vertices. In Procrustes space, we determined a new reference mesh by calculating the arithmetic mean of all Procrustes meshes within the dataset, for each bone [33]. Subsequently, a second non-rigid CPD was performed to re-register all bones to the mean mesh and obtain point correspondence across the sample of bones. This process yielded three-dimensional shape coordinates, which are the set of aligned, scaled and corresponding mesh vertices that retain only shape information. Rigid and non-rigid CPD were performed in MATLAB [31].
Sub-talar joint complex
2.3.3.3.
The sub-talar joint complex statistical shape model was created by combining the final corresponding tali and calcanei for each participant. This allowed an analysis of the variations in shape across the entire sub-talar joint complex.
Statistical analysis
2.4.
Scaling
2.4.1.
Scaling relationships of the bone and their joint articular surfaces were determined to be allometric when the 95% confidence interval (CI) for the estimated Log V – Log SA did not include isometry (0.67) [12]. Scaling with positive allometry (slope CI > 0.67) means that the SA is growing at a rate faster than that of isometric scaling, which may have implications for potentially offsetting joint pressures as humans scale. Meanwhile, scaling with negative allometry (slope CI < 0.67) is when the SA grows at a rate slower than that of isometric scaling. For each slope, a two-tailed t‐test was used to calculate the p-value to assess whether it significantly deviated from isometric scaling.
Shape and size
2.4.2.
We applied a Procrustes ANOVA to examine the relationship between size (volume) and shape for the individual bones (talus, calcaneus) and the sub-talar joint complex (talus and calcaneus combined). Procrustes ANOVA generates multivariate regression models where size is the predictor and the three-dimensional shape coordinates are the response variables [34]. A Procrustes ANOVA was applied for each individual bone (talus, calcaneus) and the entire sub-talar joint complex, using the size variables (bone/joint volume) as the predictor variables. To visualize the results of the Procrustes ANOVA, the mean shape was warped to the fitted values (shape coordinates) predicted by each regression model across the spectrum of the size variable used, e.g. smallest to largest volume, highlighting the regions of each bone that change shape with increasing or decreasing bone size. Procrustes ANOVA analysis was performed with the R packages geomorph and RRPP [35–38].
Results
Scaling results
3.1.
Talus and calcaneus volumes scaled isometrically with body height (talus: slope Log height – Log V = 3.09, 95% CI = 1.92, 4.25, figure 3A; calcaneus: slope = 3.66, 95% CI = 2.70, 4.61, figure 3B). Body height explained 46% of the variation in talus volume and 64% of calcaneus volume. The SA and volume of the talus scaled isometrically (slope Log V – Log SA = 0.67, 95% CI: 0.64, 0.70, p = 0.87, figure 3C, table 1) while the calcaneus scaled with negative allometry (slope Log V – Log SA = 0.62, 95% CI: 0.59, 0.65, p = 0.0037, figure 3D, table 1). This indicates that as bone volume increased, the SA of the calcaneus grew slower than expected under isometric scaling.
Scaling plot (Log Height – Log V) of (A) the talus with individual participant points and line of best fit in blueScaling plot (Log Height – Log V) of (A) the talus with individual participant points and line of best fit in blue, (B) the calcaneus with individual participant points and line of best fit in red, with isometric scaling line in black (slope = 3). Scaling plots (Log V – Log SA) of (C) the talus with individual participant points and line of best fit in blue and (D) the calcaneus with individual participant points and line of best fit in red, with the isometric scaling line in black (slope = 0.67).
The articular surfaces on the talus (figure 4A) both scaled isometrically, following the same pattern of the entire talus bone (slope Log V – Log SA = 0.71 for both the sub-talar facet and talar dome, sub-talar facet 95% CI: 0.57, 0.85, talar dome 95% CI: 0.61, 0.81, electronic supplementary material, figure S1, table 1). The posterior sub-talar facet and the anterior-medial facet on the calcaneus (figure 4B) also scaled with isometry (posterior facet slope Log V – Log SA = 0.73, 95% CI: 0.52, 0.95, figure 5A, anterior-medial facet slope Log V – Log SA = 0.43, 95% CI: 0.11, 0.75, figure 5B, table 1).
Articular surfaces of (A) the talus, talar dome in green and sub-talar facet in blue, and (B) calcaneus, posterior facet in purple and anterior-medial facet in pink.
Scaling plot (Log V – Log SA) of calcaneus articular surfaces, (A) the posterior facet (PF) with individual participant points and line of best fit in purple, (B) the anterior-medial facet (AMF) with individual participant points and line of best fit in pink. The isometric scaling line (slope = 0.67) is in black across all graphs.
Relationship between bone shape and bone size
3.2.
We observed no relationship between talus shape and volume (size) (p = 0.07, R^2^ = 0.04, Z = 1.5). The shape features of the calcaneus were related to calcaneus volume (p = 0.003, R^2^ = 0.06, Z = 2.7). As the calcaneus increased in size, the bone appeared to become relatively taller (superior-inferiorly), wider (medio-laterally) and shorter (antero-posteriorly). Collectively, these shape features appear to make the calcaneus become more ‘cube-like’ as it becomes larger (figure 6). In addition, the inferior aspect of the cuboid facet is elongated while becoming more curved.
Anatomical shape variations of the calcaneus linked to calcaneal volume. Green indicates where the bone is larger and outside the mean shape, while purple shows where the bone sits inside the mean bone. The arrows focus on the location and direction of change (pointing at the bone indicates decreased size of a feature, whereas an arrow pointing away from the bone indicates increased size of a feature).
Shape features of the sub-talar joint complex were also related to size (combined calcaneus and talus volume, p = 0.001, R^2^ = 0.06, Z = 3.1). As the joint complex becomes larger, we qualitatively observe similar shape changes in the calcaneus to those observed at the individual bone level. The calcaneus became relatively taller, wider and shorter with increasing joint volume (figure 7). Additionally, the joint-level analysis indicated shape–size relationships at the talus, with a relative shortening of the posterior process and the talar dome growing in width less than what would be expected for isometric scaling, appearing to become narrower as the joint-complex increases in size.
Anatomical shape variations of the sub-talar joint complex linked to sub-talar joint volume (combined calcaneus and talus volume). Green indicates where the bone is larger and outside the mean shape, while purple shows where the bone sits inside the mean bone. The arrows focus on the location and direction of change (pointing at the bone indicates decreased size of a feature, whereas an arrow pointing away from the bone indicates increased size of a feature).
Discussion
Human tarsal bones vary widely in shape and size. We sought to understand if the tarsal bones change shape as a function of size to inform our understanding of ankle and sub-talar joint contact mechanics during locomotion. Our hypothesis was not supported for the talus, which exhibited isometry and had no distinct shape–size relationships at the articular surface or individual bone level. However, the calcaneus and the sub-talar joint complex did show distinct shape–size relationships at the levels of the articular surface, individual bone and the sub-talar joint complex. The nature of these relationships varied, with differing scaling patterns and shape–size relationships occurring across each structural level. Despite this variability, it appears that shape features of the calcaneus dominate the shape–size adaptations of the sub-talar joint complex.
We observed deviations in scaling behaviour between the calcaneus bone and its joint articular surfaces, indicating region-specific scaling strategies in the sub-talar joint complex. We found the calcaneus scales with negative allometry; however, the posterior facet and anterior-medial facet scale with isometry, indicating that the SA of the joint is growing at a relatively faster rate than the calcaneus. The divergent scaling strategies of the joint articular surfaces and the calcaneus are largely consistent with the theory that the joint SA needs to grow at a faster rate to maintain relatively constant joint contact stresses across a range of skeletal sizes; however, we see this relative to the scaling of the bone rather than observing positive allometry [4].
Clear shape–size relationships were observed for the calcaneus but not for the talus. Given that the talus scaled isometrically, this result is unsurprising. Interestingly, as the calcaneus grows in size, it becomes taller (supero-inferiorly), wider (medio-laterally) and relatively shorter (antero-posteriorly). The medial calcaneal tubercle (the origin of the plantar fascia) becomes enlarged (figure 6). Collectively, these shape features appear to transition the calcaneus to a more cube-like (robust) shape as it increases in size. A similar pattern is observed across non-human primates, where more terrestrial species, such as gorillas, exhibit a wider tubercle compared with more arboreal apes, like orangutans [17]. This trend extends to subspecies of gorillas, with more terrestrial subspecies having relatively mediolaterally and anteroposteriorly shorter calcanei [40]. This adaptation is thought to increase the contact area with the ground, helping to dissipate the increased loads transmitted through the calcaneus [40]. This observation aligns with our finding of negative allometric scaling in this bone, as becoming more cube-like would lead to relatively lower growth in SA, as would be expected under isometric conditions [4]. The calcaneus is encumbered with large magnitudes of repetitive forces applied from the ground due to gravity and also from muscular forces (e.g. Achilles tendon) [41]. Given that gravitational and muscular forces scale with size [4,6], a more cube-like shape may improve the capacity of the bone to manage increasing internal bone stresses [4,8]. Increased calcaneal height, in particular, is suggested to counteract the bending stresses imposed on the calcaneus during locomotion due to the large Achilles tendon force and the forces applied by the intrinsic foot muscles and plantar fascia [12,42].
The calcaneus and talus do not function in isolation but rather as an articular complex (sub-talar joint). Therefore, we sought to understand how these two bones change shape together as a function of size. Unlike the individual bone analysis, we observed specific shape features in both the talus and calcaneus that were related to size. The shape features observed in the calcaneus as part of the sub-talar joint complex were similar to the features observed at the individual bone level. The observed shortening of the talar posterior process and widening of the talar dome appear to be in response to the relative increase in width and decrease in length of the calcaneus with increasing size (figure 7). Given that the shape features of the calcaneus are similar at a bone and sub-talar joint complex level, it appears that the shape changes in this bone are driving the shape features across the entire complex. The talus sits within the ankle joint mortise and has no direct muscular attachments or direct interaction with the ground. Therefore, being constrained by the surrounding bones (calcaneus, tibia, navicular) may provide less opportunity for the talus to change shape without concomitant changes in the other surrounding bones.
Some methodological limitations should be considered when contextualizing our findings. We have not included sex as a covariate in our statistical model due to sample size or age due to data not being available for all participants. We chose to quantify size using bone and joint complex volume (combined talus and calcaneus volume) to isolate how bones change shape as a function of their own size. While hindfoot bone volume and body height had a significant relationship, there is still substantial variation present, which may limit our ability to infer the relationship between shape changes and loads associated with body size. However, the steeper scaling slope of the calcaneus compared with the talus may suggest that the calcaneus adapts more rapidly to increases in body size, reflecting its primary role in load bearing. Future studies that incorporate function, including the contribution of soft tissues and direct or indirect measures of joint contact mechanics may improve our understanding of the shape, size and function relationships within the human foot.
In summary, we observed articular surface, bone and joint complex-level shape–size relationships. These relationships are variable and appear to reflect the competing demands of balancing healthy joint contact mechanics and internal bone stress. These findings have important implications for the development and progression of degenerative joint conditions such as osteoarthritis. Further research exploring how joint posture and mobility interact with shape and size to determine joint contact mechanics have the potential to enhance the management of these conditions via surgical and conservative means.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Jurca A, Žabkar J, Džeroski S. 2019 Analysis of 1.2 million foot scans from North America, Europe and Asia. Sci. Rep. 9, 19155. (10.1038/s 41598-019-55432-z)31844106 PMC 6914786 · doi ↗ · pubmed ↗
- 2Schuster RW, Cresswell A, Kelly L. 2021 Reliability and quality of statistical shape and deformation models constructed from optical foot scans. J. Biomech. 115, 110137. (10.1016/j.jbiomech.2020.110137)33267964 · doi ↗ · pubmed ↗
- 3Schuster RW, Cresswell AG, Kelly LA. 2023 Foot shape is related to load-induced shape deformations, but neither are good predictors of plantar soft tissue stiffness. J. R. Soc. Interface 20, 20220758. (10.1098/rsif.2022.0758)36651181 PMC 9846431 · doi ↗ · pubmed ↗
- 4Mc Mahon TA. 1975 Using body size to understand the structural design of animals: quadrupedal locomotion. J. Appl. Physiol. 39, 619–627. (10.1152/jappl.1975.39.4.619)1194153 · doi ↗ · pubmed ↗
- 5Bertram JEA, Biewener AA. 1990 Differential scaling of the long bones in the terrestrial carnivora and other mammals. J. Morphol. 204, 157–169. (10.1002/jmor.1052040205)2348461 · doi ↗ · pubmed ↗
- 6Dick TJM, Clemente CJ. 2017 Where have all the giants gone? How animals deal with the problem of size. P Lo S Biol. (ed A Hedenström), 15, e 2000473. (10.1371/journal.pbio.2000473)28076354 PMC 5226675 · doi ↗ · pubmed ↗
- 7Malda J, de Grauw JC, Benders KEM, Kik MJL, van de Lest CHA, Creemers LB, Dhert WJA, van Weeren PR. 2013 Of mice, men and elephants: the relation between articular cartilage thickness and body mass. P Lo S One (ed. J Orgel), 8, e 57683. (10.1371/journal.pone.0057683)23437402 PMC 3578797 · doi ↗ · pubmed ↗
- 8Mc Mahon TA. 1975 Allometry and biomechanics: limb bones in adult ungulates. Am. Nat. 109, 547–563. (10.1086/283026) · doi ↗