A new Drosophila melanogaster research resource: CRISPR-induced mutations for clonal analysis of fourth chromosome genes
Bonnie M Weasner, Brandon P Weasner, Kevin R Cook, Michael J Stinchfield, Shu Kondo, Kuniaki Saito, Justin P Kumar, Stuart J Newfeld

TL;DR
Researchers created a new resource using CRISPR to study mutations in genes on the fourth chromosome of fruit flies.
Contribution
The study provides a comprehensive set of CRISPR-induced mutations for 62 fourth chromosome genes in Drosophila melanogaster.
Findings
CRISPR was used to generate mutations in 62 of 80 fourth chromosome protein-coding genes.
84 of the 119 mutations are likely null alleles due to frameshifts.
Mutations were tested for lethal, female-sterile, and visible phenotypes.
Abstract
As part of an ongoing effort to generate comprehensive resources for the experimental analysis of fourth chromosome genes in Drosophila melanogaster, the Fourth Chromosome Resource Project has used CRISPR mutagenesis with single guide RNAs to isolate mutations in 62 of the 80 fourth chromosome, protein-coding genes. These mutations were induced on a fourth chromosome bearing a basal FRT insertion to facilitate experimental approaches involving FLP recombinase-induced mitotic recombination. To permit straightforward comparisons among mutant stocks, most of the mutations were generated on isogenic fourth chromosomes, which were then crossed into a common genetic background. Of the 119 mutations, 84 are frameshift mutations likely to be null alleles, 29 are small, in-frame deletions, and 6 have yet to be characterized molecularly. The mutations were tested for recessive lethal,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
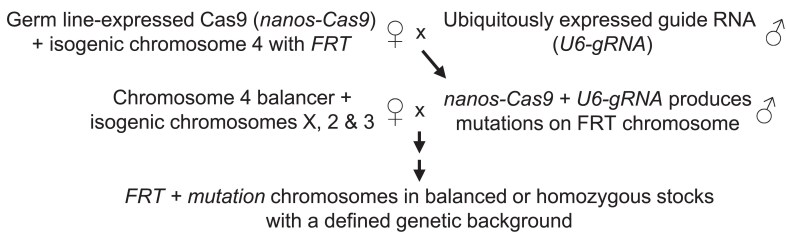 Fig. 1
Fig. 1| Gene | Mutagenized chromosomes recovered | Frameshift mutations | In-frame deletions | No mutation |
|---|---|---|---|---|
|
| ||||
|
| 3 | 2 | 0 | 0 |
|
| 7 | 3 | 1 | 0 |
|
| 2 | 1 | 1 | 0 |
|
| 5 | 1 | 0 | 0 |
|
| 3 | 3 | 0 | 0 |
|
| 4 | 3 Identical | 1 | 0 |
|
| 4 | 1 | 0 | 0 |
|
| 2 | 0 | 1 | 0 |
|
| 5 | 1 | 0 | 1 |
|
| 8 | 1 | 0 | 3 |
|
| 4 | 0 | 1 | 2 |
|
| 8 | 2 | 0 | 0 |
|
| 5 | 1 | 0 | 0 |
|
| 4 | 2 | 1 | 0 |
|
| 2 | 1 | 1 | 0 |
|
| 3 | 2 | 0 | 0 |
|
| 4 | 3 (2 identical) | 1 | 0 |
|
| 8 | 2 | 1 | 4 |
|
| 3 | 3 Identical | 0 | 0 |
|
| 1 | 1 | 0 | 0 |
|
| 4 | 1 | 2 | 0 |
|
| 11 | 4 Identical | 1 | 6 |
|
| 3 | 0 | 2 Identical | 0 |
|
| 3 | 0 | 0 | 0 |
|
| 6 | 2 | 1 | 3 |
|
| 2 | 0 | 2 | 0 |
|
| 5 | 4 (2 identical) | 0 | 0 |
|
| 1 | 1 | 0 | 0 |
|
| 5 | 5 (3 identical) | 0 | 0 |
|
| 3 | 1 | 0 | 0 |
|
| 7 | 2 | 0 | 2 |
|
| 5 | 3 Identical | 2 Identical | 0 |
|
| 2 | 0 | 1 | 1 |
|
| 3 | 1 | 1 | 1 |
|
| 4 | 2 | 0 | 2 |
|
| 3 | 0 | 2 Identical | 1 |
|
| 4 | 4 | 0 | 0 |
|
| 3 | 0 | 0 | 0 |
|
| 3 | 1 | 2 Identical | 0 |
|
| 3 | 0 | 3 Identical | 0 |
|
| 5 | 5 (2 identical) | 0 | 0 |
|
| 4 | 3 | 0 | 1 |
|
| 2 | 2 | 0 | 0 |
|
| 5 | 1 | 0 | 2 |
|
| 4 | 1 | 0 | 1 |
|
| 5 | 1 | 0 | 4 |
|
| 9 | 3 | 2 Identical | 1 |
|
| 5 | 3 (2 identical) | 0 | 2 |
|
| 6 | 6 (4 identical) | 0 | 0 |
|
| 2 | 0 | 1 | 1 |
|
| 5 | 2 Identical | 1 | 2 |
|
| 4 | 0 | 2 | 2 |
|
| 3 | 1 | 2 | 0 |
|
| 3 | 2 Identical | 1 | 0 |
|
| 8 | 4 (2 identical) | 4 (3 identical) | 0 |
|
| 5 | 4 (3 identical) | 1 | 0 |
|
| 1 | 0 | 1 | 0 |
|
| 3 | 2 | 0 | 0 |
|
| 5 | 5 (4 identical) | 0 | 0 |
|
| 3 | 3 Identical | 0 | 0 |
|
| ||||
|
| 8 | 6 Identical | 2 Identical | 0 |
|
| 9 | 2 | 2 | 4 |
|
| 4 | 0 | 1 | 0 |
|
| 2 | 1 | 1 | 0 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
- —National Institutes of Health10.13039/100000002
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCRISPR and Genetic Engineering · Chromosomal and Genetic Variations · Plant Virus Research Studies
Introduction
The specific roles a gene plays in the normal functioning of a cell are often inferred from the phenotypes seen when its activity is reduced or eliminated, and loss-of-function mutations are important for such experimental approaches. While the functional significance of a gene can sometimes be determined from phenotypes seen in individuals composed entirely of homozygous mutant cells, it is often more informative to examine the phenotypes of homozygous mutant cells embedded in an otherwise heterozygous individual. Indeed, experimental approaches involving mitotic recombination to generate small clones of mutant cells are particularly important when homozygosity results in zygotic lethality.
Most current clonal analysis experiments in Drosophila involve the use of the GAL4–UAS–GAL80 cell-specific expression system to control FLP recombinase-induced mitotic recombination between FRT sequences positioned at the bases of chromosome arms. These methods permit clone formation to be targeted spatially and temporally, and they allow investigators to determine the contributions of individual genes to the development, homeostasis, and adaptation of specific tissues (Weasner et al. 2017). These methods have become especially important in exploring cell-to-cell communication. For example, they are used extensively in studying competition among cells (Kiparaki and Baker 2023).
Here, we describe the efforts of the Fourth Chromosome Resource Project to develop Drosophila stocks for the clonal analysis of loss-of-function phenotypes for the majority of fourth chromosome, protein-coding genes. This work was prompted by the paucity of research resources related to the fourth chromosome and by 2 experimental developments. First, Goldsmith et al. (2022) used CRISPR site-specific genome-editing technology to insert an FRT sequence at the base of the long arm of the fourth chromosome (the short arm carries no genes), which they then used to generate mitotic clones. Second, a set of stocks bearing guide RNA-expressing transgenes for most protein-coding genes on the fourth chromosome was generated at the National Institute of Genetics in Japan. We recognized that the easiest way to place mutations distal to this basal FRT was to use these guide RNAs to induce mutations directly on the FRT-bearing chromosome. To add experimental value to the stock collection, we have taken care to generate the mutations on fourth chromosomes derived from a single progenitor chromosome and to place the mutagenized chromosomes into a common genetic background. The new mutation-bearing stocks are now being distributed by stock centers in both Japan and the United States.
Materials and methods
An extensive description of our experimental methods is provided in Supplementary File 1. It provides details of the screens isolating CRISPR-induced mutations on a single-origin fourth chromosome carrying the FRT transgene TI{TI}FRT101F (Goldsmith et al. 2022); the crosses placing the mutagenized chromosomes into a common, isogenic background; the construction of stocks necessary for these crosses; the molecular characterization of the mutations; and the preliminary characterization of associated phenotypes. Mutations in 4 genes were generated by alternative crosses, which are also outlined. Supplementary Table 1 gives a list of fourth chromosome genes. Supplementary Table 2 provides details of the guide RNA transgenes used in the screens. Supplementary Table 3 gives primers utilized to identify and characterize new mutations. Supplementary Table 4 is the Reagent Table listing the progenitor stocks used to develop the strains employed in the screens, the stocks used to characterize mutant phenotypes, and the sources of biological and chemical reagents.
All fly cultures were reared under standard conditions. All genomic coordinates cited in this report refer to D. melanogaster genome release 6. Gene models, annotations, and nomenclature reflect FlyBase release 2024_4. Information about the new mutations in publicly distributed stocks has been archived in FlyBase (Öztürk-Çolak et al. 2024). Information about the stocks may be obtained via the websites of the Bloomington Drosophila Stock Center at Indiana University (https://bdsc.indiana.edu/) and the Department of Drosophila Genomics and Genetic Resources at the Kyoto Institute of Technology (https://www.dgrc.kit.ac.jp/). A preliminary report of this work was presented by Stinchfield et al. (2024).
Results and discussion
As mentioned, our effort to generate CRISPR-induced mutations in fourth chromosome genes on an FRT-bearing chromosome depended on the availability of a set of guide RNA-expressing stocks from the Japan National Institute of Genetics. These stocks were generated as part of a large-scale project to construct guide RNA-expressing stocks for all D. melanogaster protein-coding genes, which will be described elsewhere. The transgenes in these stocks were constructed by the methods of Kondo and Ueda (2013), and they ubiquitously express guide RNAs with 20-base targeting sequences under the control of snRNA:U6:96Ab regulatory sequences. Stocks targeting 77 of the 80 protein-coding genes on the fourth chromosome were available for our efforts (those we used are listed in Supplementary Table 2). In addition, we used 1 guide RNA stock originating in the work of Zirin et al. (2020).
As outlined in Fig. 1 and shown in detail in Supplementary File 1, the guide RNA-expressing transgenes were combined in males with a transgene expressing the Cas9 nuclease in the germ line and a fourth chromosome carrying the FRT transgene TI{TI}FRT101F. All FRT*-*bearing chromosomes used in screens were derived from a single chromosome to minimize sequence variation across all screens. Putative mutation-bearing fourth chromosomes were isolated in balanced or homozygous stocks and tested for alterations at the guide RNA target sites by PCR amplification followed by Sanger sequencing. We generally found it necessary to sequence fewer than 10 mutagenized chromosomes per gene to identify 1 or more independent mutations. Table 1 summarizes the results of the 69 screens undertaken using this method and Supplementary Table 5 describes the mutations chosen for phenotypic characterization. In several instances, we recovered independent mutations with identical molecular lesions—reflecting the predictability of CRISPR mutagenesis and DNA repair (Table 1).
Conceptual outline of the CRISPR screens producing new fourth chromosome mutations. In males, a transgene expressing Cas9 in the germ line (nanos-Cas9) was combined with a transgene ubiquitously expressing a guide RNA targeting a specific fourth chromosome gene (U6-gRNA) and a transgene carrying an FRT sequence inserted near the base of an isogenized fourth chromosome. Many of the gametes produced by these males carried mutations in the targeted gene caused by Cas9-induced double-stranded DNA breaks. A series of crosses involving stocks with balancer chromosomes and isogenic X, second, and third chromosomes produced balanced or homozygous stocks with a defined genetic background. Supplementary File 1 provides a detailed description of the screens and the development of the stocks used.
The crosses to establish new mutations in balanced or homozygous stocks simultaneously placed the mutagenized fourth chromosomes into a defined genetic background so that, to the extent practical, the mutation-bearing stocks would differ at the sequence level only by the induced mutations. As shown in Supplementary File 1, the isogenic background consists of second and third chromosomes derived from the genetic background used for the DrosDel deletions (Ryder et al. 2007), the Dp(1;Y)BSC chromosomes (Cook et al. 2010) and their progenitor insertions (Ryder et al. 2004), and an X chromosome marked with y^1^ and w^1118^ derived from a heavily used Bloomington stock. The Y chromosomes in the final stocks are derived from the guide RNA stocks and cannot be assumed to be uniform. Fourth chromosomes bearing dominant mutations with visible phenotypes (e.g. ci^D^ or Gat^eya^) or transgenes expressing fluorescent reporter proteins (e.g. P{ActGFP}unc-13^GJ^) were used to balance the new, fourth chromosome mutations. Supplementary Table 6 lists the stocks that have been placed into public distribution to date at the Drosophila stock centers in Bloomington and Kyoto.
We encountered idiosyncratic lethality and sterility in the early crosses of 3 screens. While these problems were unrelated to the mutageneses per se and are likely attributable to cryptic mutations in the guide RNA stocks, Table 1 shows that we were able to recover mutations using the same guide RNA stocks in alternative crosses (described in Supplementary File 1). Supplementary Table 5 describes the mutations and Supplementary Table 6 lists the stocks. Note that these screens and subsequent stock constructions did not rigorously control the genetic backgrounds and that future screens by the Fourth Chromosome Resource Project will use variations of this approach and will, likewise, not control for genetic background in a disciplined way.
The predominant outcome of the mutageneses was the induction of small deletions (Supplementary Table 5). In any mutagenesis, deletions that result in frameshifted translation lead to the production of truncated proteins, while in-frame deletions lead to the loss of a small number of protein residues. Frameshifted alleles that eliminate substantial portions of proteins are likely to be amorphic, while in-frame deletion alleles may or may not affect protein function depending on the consequences of removing particular amino acids. Our goal was to recover at least 1 truncation mutation for each gene screened. We were largely successful as we recovered truncation mutations in 52 of the 72 genes for which we undertook screens (Table 1). Since the median position of the truncations was 14% of normal protein length (Supplementary Table 5), the mutations likely shorten proteins enough to result in null phenotypes. For 51 of the 52 genes with truncation mutations, all predicted protein isoforms are affected (2 of the 7 predicted Crk isoforms are unaffected by Crk^E^ even though, as described in the following, we showed it is a loss-of-function allele). The Fourth Chromosome Resource Project will revisit the genes for which we recovered no truncation alleles. We identified mutations in 3 genes from their lethality in complementation tests even though we have yet to identify the molecular lesions (Table 1).
While loss-of-function phenotypes have been characterized for many fourth chromosome genes, they are unknown for others. To evaluate, at least superficially, the effects of disrupting fourth chromosome genes, we assessed the viability, female fertility, and gross morphology of flies in which the new alleles were homozygous and in flies in which the new alleles were combined with preexisting mutations or chromosomal deficiencies that delete the targeted genes (the deficiencies are described in Supplementary Table 7, control crosses validating the deficiency and preexisting mutation stocks are shown in Supplementary Table 8, and the crosses evaluating the new mutations are detailed in Supplementary Table 9). Our results are summarized in Supplementary Table 10, which shows that the phenotypes of mutation homozygotes and hemizygotes are consistent. Homozygotes occasionally have more severe phenotypes than hemizygotes as one might expect if linked, deleterious mutations—preexisting or generated in the course of the crosses—were homozygosed along with the targeted mutations. Our results are presented in the context of previous phenotypic studies in Supplementary Table 11. This analysis allowed us to define the loss-of-function phenotypes for 35 genes that had never been characterized: 12 are necessary for viability while 23 have no overt loss-of-function phenotypes. It also allowed us to confirm and extend the previous characterizations of 29 genes and provide evidence that 3 in-frame deletions from our screens are loss-of-function alleles.
Several decades ago, Benjamin Hochman and colleagues attempted to determine the number of fourth chromosome loci needed for viability or female fertility via mutagenesis and complementation analyses [reviewed in Hochman (1976)]. They identified 37 loci, but they did not know how close they were to isolating mutations in all vital loci. They were aware that factors such as complex intragenic complementation patterns and the low mutability of some genes make it difficult to arrive at a definitive count of genes using their approach. Our analysis of protein-coding genes (Table 2) shows 39 genes needed for viability, 1 gene needed strictly for male fertility, 33 genes with no overt phenotypes, and 7 genes for which there is no information. We have seen no evidence that any fourth chromosome genes have phenotypes affecting only external morphology or female fertility. (The absence of genes devoted exclusively to female fertility is not surprising: Perrimon et al. (1986) estimated that the X chromosome, which has more than 20 times as many genes as the fourth chromosome, carries only 15 genes needed solely for female reproduction.) There is a growing consensus that noncoding RNA genes usually have subtle loss-of-function phenotypes (Liu et al. 2017; Schor et al. 2018; Camilleri-Robles et al. 2024). The lack of obvious phenotypes associated with deletion of the antisense RNA gene asRNA:CR44031 (Supplementary Tables 8 and 11) and increased male-to-male courtship observed as the only apparent effect associated with deletion of the long noncoding RNA gene lncRNA:sphinx (Dai et al. 2008) are consistent with this view. If we assume that the remaining 25 noncoding RNA genes are unlikely to have lethal or sterile phenotypes, our relatively complete inventory agrees well with the count of Hochman and colleagues, suggesting that their studies were closer to identifying mutations in all vital loci than Hochman concluded in his most comprehensive review (Hochman 1976). Completion of the Fourth Chromosome Resource Project will facilitate a comprehensive assessment of loss-of-function phenotypes (including phenotypes more subtle than we assessed in our tests), and it will then be possible to determine how the Hochman vital complementation groups, most of which are still represented in extant stocks, correspond to molecularly defined genes.
We have confirmed the presence of the basal FRT-bearing transgene TI{TI}FRT101F in all the final stocks by PCR amplification. Although there is no reason to suspect the mutagenesis procedure methodically corrupted the FRT-bearing transgene, we have not systematically verified that the mutation-bearing chromosomes are capable of participating in FLP-mediated mitotic recombination. Nonetheless, Stinchfield et al. (2024) successfully recovered mitotic clones with fourth chromosomes from 2 stocks (Pur-alpha^B^ and zfh2^51A^).
To assess whether the mutagenesis procedure produced off-target mutations on the fourth chromosome, we sampled stocks for the presence of unexpected mutations producing lethal, female-sterile, or visible phenotypes (Supplementary Table 12). We tested twelve chromosomes with targeted mutations that were lethal (Actbeta^B^, apolpp^B^, Arf4^A^, ATPsynbeta^C^, bt^A^, CG31998^C^, CG31998^D^, CG33978^B^, ci^C^, ey^B^, pho^F^, sv^F^, toy^C^) for the presence of additional deleterious mutations by combining these chromosomes via crosses with a panel of deficiencies and mutations that together delete or disrupt all fourth chromosome genes. None of these chromosomes showed evidence of additional mutations. Similarly, we tested 5 chromosomes with targeted mutations that produced no overt phenotypes in combination with deficiencies or preexisting mutations yet showed appreciable recessive lethality or sterility in stocks (CG32017^K^, Gyf^F^, Hcf^A^, Pur-alpha^B^, Slip1^A^). None of these chromosomes had additional mutations. The poor viability in these and similar stocks may come from sublethal effects of the targeted mutations combined with sublethal effects of other mutations that were induced during the mutagenesis or that arose spontaneously in subsequent culture. The conclusions we can draw from all these crosses are limited: we did not test all mutated chromosomes, and it was impossible to test for off-target mutations in every fourth chromosome gene because some genes are deleted only by the same deficiencies as targeted genes. Nevertheless, these results indicate that off-target mutations were not common.
In summary, the work described here has provided a new resource for examining the cellular functions of most protein-coding genes on the fourth chromosome. It complements the other resources being generated by the Fourth Chromosome Resource Project (Stinchfield et al. 2024) including UAS constructs for fourth chromosome gene misexpression and phenotypic rescue, GAL4 knock-ins for expressing UAS constructs in the normal patterns of fourth chromosome genes, UAS constructs for the human homologs of Drosophila fourth chromosome genes for creating “humanized” flies with human gene expression substituted for fly gene expression, GFP knock-ins into fourth chromosome genes for expressing fluor-tagged proteins in native gene patterns, and UAS-RNAi transgenes for GAL4-directed, tissue-specific knockdown of fourth chromosome genes. Our hope is that these resources will alleviate many of the difficulties that have historically hampered investigations of fourth chromosome gene functions.
Supplementary Material
jkaf006_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Camilleri-Robles C, Amador R, Tiebe M, Teleman AA, Serras F, Guigó R, Corominas M. 2024. Long non-coding RN As involved in Drosophila development and regeneration. NAR Genom Bioinform. 6(3):lqae 091. doi:10.1093/nargab/lqae 091.39157585 PMC 11327875 · doi ↗ · pubmed ↗
- 2Cook RK, Deal ME, Deal JA, Garton RD, Brown CA, Ward ME, Andrade RS, Spana EP, Kaufman TC, Cook KR. 2010. A new resource for characterizing X-linked genes in Drosophila melanogaster: systematic coverage and subdivision of the X chromosome with nested, Y-linked duplications. Genetics. 186(4):1095–1109. doi:10.1534/genetics.110.123265.20876560 PMC 2998296 · doi ↗ · pubmed ↗
- 3Dai H, Chen Y, Chen S, Mao Q, Kennedy D, Landback P, Eyre-Walker A, Du W, Long M. 2008. The evolution of courtship behaviors through the origination of a new gene in Drosophila. Proc Natl Acad Sci USA. 105(21):7478–7483. doi:10.1073/pnas.0800693105.18508971 PMC 2396706 · doi ↗ · pubmed ↗
- 4Goldsmith SL, Shimell M, Tauscher P, Daly SM, Shimmi O, O'Connor MB, Newfeld SJ. 2022. New resources for the Drosophila 4th chromosome: FRT 101F enabled mitotic clones and Bloom syndrome helicase enabled meiotic recombination. G 3 (Bethesda). 12(4):jkac 019. doi:10.1093/g 3journal/jkac 019.35084488 PMC 8982423 · doi ↗ · pubmed ↗
- 5Hochman B . 1976. The fourth chromosome of Drosophila melanogaster. In: Ashburner M, Novitski E, editors. The genetics and biology of Drosophila. Academic Press. p. 903–928.
- 6Kiparaki M, Baker NE. 2023. Ribosomal protein mutations and cell competition: autonomous and nonautonomous effects on a stress response. Genetics. 224(3):iyad 080. doi:10.1093/genetics/iyad 080.37267156 PMC 10691752 · doi ↗ · pubmed ↗
- 7Kondo S, Ueda R. 2013. Highly improved gene targeting by germline-specific Cas 9 expression in Drosophila. Genetics. 195(3):715–721. doi:10.1534/genetics.113.156737.24002648 PMC 3813859 · doi ↗ · pubmed ↗
- 8Liu SJ, Horlbeck MA, Cho SW, Birk HS, Malatesta M, He D, Attenello FJ, Villalta JE, Cho MY, Chen Y, et al 2017. CRISP Ri-based genome-scale identification of functional long noncoding RNA loci in human cells. Science. 355(6320):eaah 7111. doi:10.1126/science.aah 7111.PMC 539492627980086 · doi ↗ · pubmed ↗