Repetitive DNAs and differentiation of the ZZ/ZW sex chromosome system in the combtail fish Belontia hasselti (Perciformes: Osphronemidae)
Alan Moura de Oliveira, Geize Aparecida Deon, Alexandr Sember, Caio Augusto Gomes Goes, Weerayuth Supiwong, Alongklod Tanomtong, Fábio Porto-Foresti, Ricardo Utsunomia, Thomas Liehr, Marcelo de Bello Cioffi

TL;DR
This study explores the sex chromosome system in Java combtail fish and finds limited differentiation despite visible differences.
Contribution
The first satellitome study in Perciformes reveals low satellite DNA diversity and minimal sex chromosome differentiation in B. hasselti.
Findings
Thirteen satellite DNA monomers were identified, indicating low satDNA diversity in B. hasselti.
Repetitive DNA clusters were primarily found on autosomes, not sex chromosomes.
Comparative genomic hybridization found no female-specific repeats on the W chromosome.
Abstract
Java combtail fish Belontia hasselti (Cuvier, 1831), a member of the Osphronemidae family, inhabits lakes and rivers throughout Southeast Asia and Sri Lanka. Previous cytogenetic research revealed it possesses a diploid chromosome number of 48 chromosomes with a female-heterogametic ZZ/ZW sex chromosome system, where the W chromosome is distinguishable as the only metacentric element in the complement. Female-heterogametic sex chromosome systems seem to be otherwise surprisingly rare in the highly diverse order Perciformes and, therefore, B. hasselti provides an important comparative model to evolutionary studies in this teleost lineage. To examine the level of sex chromosome differentiation in B. hasselti and the contribution of repetitive DNAs to this process we combined bioinformatic analyses with chromosomal mapping of selected repetitive DNA classes, and comparative genomic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
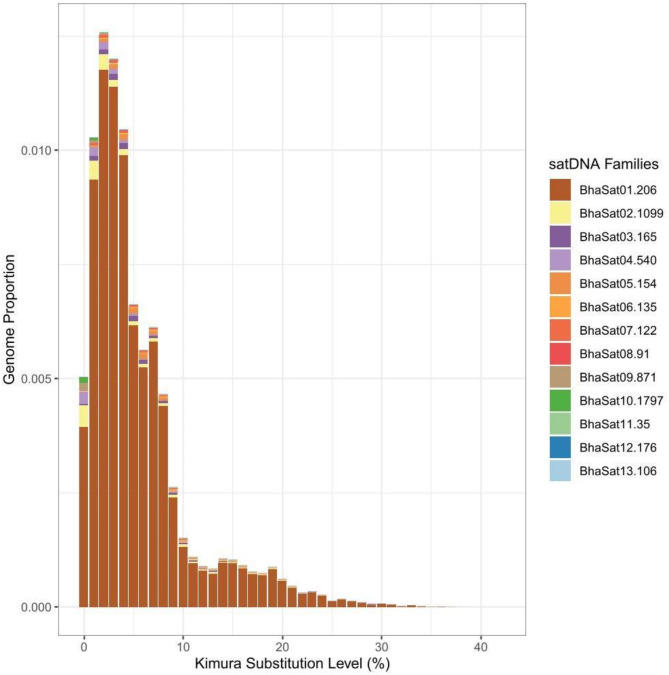 Figure 1
Figure 1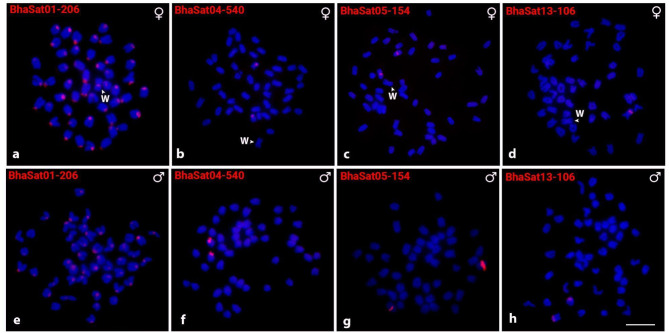 Figure 2
Figure 2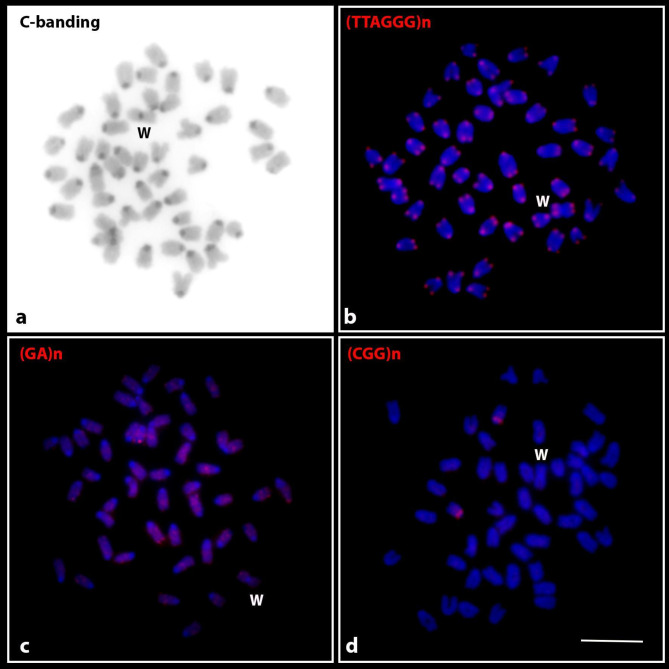 Figure 3
Figure 3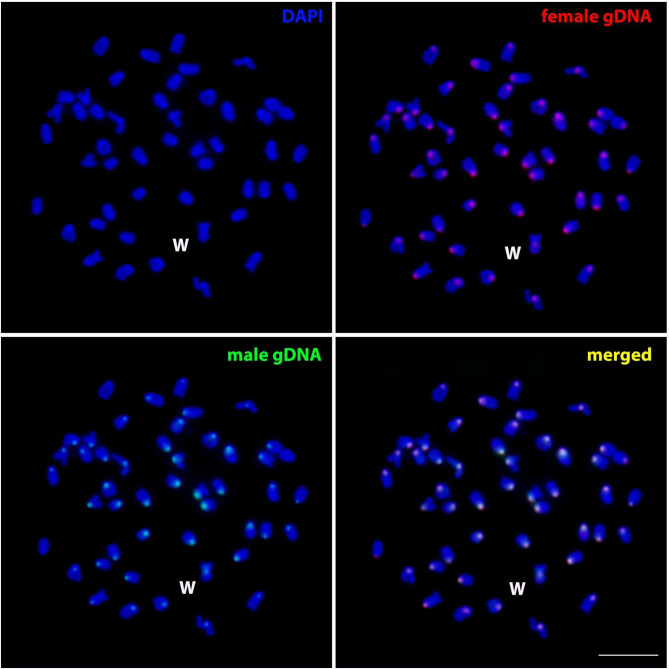 Figure 4
Figure 4- —Friedrich-Schiller-Universität Jena (1010)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetic and Clinical Aspects of Sex Determination and Chromosomal Abnormalities · Chromosomal and Genetic Variations · Genetic diversity and population structure
Introduction
Sex chromosomes, the most prevalent form of genetic sex determination in eukaryotes, evolve through progressive loss of recombination and sequence differentiation. This process displays variable pace among diverse clades. It might result in morphologically different (i.e. heteromorphic) sex chromosome counterparts while in some other cases the sex chromosomes remain (not necessarily due to short evolutionary time) undifferentiated and cytologically indistinguishable (i.e. homomorphic) [1]. Despite over a century of intense research into sex chromosome evolution in numerous eukaryotic lineages, many long-standing questions remain unresolved [1–6]. Teleost fishes provide a broad spectrum of important models to tackle them thanks to the immense variability of sex chromosome systems, their degree of differentiation, and the variability of these patterns at various taxonomic levels, including between populations of the same species in many cases [7–10]. Regarding the female-heterogametic ♂ZZ/♀ZW sex chromosome systems, besides the minority of the homomorphic ones described (e.g. in Seriola fishes [11–13] or several cichlids [14, 15]), a considerable portion of reported cases is highly heteromorphic, with the most studied examples concentrated in Neotropical ichthyofauna (e.g., genera Characidium, Parodon, Apareiodon, Megaleporinus, and Triportheus; e.g [16–21]). Given the long tracts of highly repetitive regions, only recently a more detailed genomic insight has been performed in Neotropical freshwater fishes with highly heteromorphic ZW sex chromosomes [22, 23]. To fully understand the range of variability regarding the sex chromosome evolution and sex-determining pathways in teleosts, more ZW systems should be investigated in various lineages. A particularly intriguing question is: what factors may contribute to their greater differentiation via repetitive DNA accumulation in comparison to established XY and XY-derived systems? In this context, the order Perciformes is particularly interesting, with a highly biased incidence of male-heterogametic systems [7]. In addition, some of the few ZW systems proposed to exist within this species-rich teleost order have been questioned [24] or speculative [25]. Thus, strictly speaking, besides the report of Epinephelus tauvina (Fabricius, 1775) [26], the only perciform fish to exhibit heteromorphic ZW sex chromosomes is the Java combtail fish Belontia hasselti (Cuvier, 1831) [27].
Belontiinae represents one of the four subfamilies within Osphoronemidae – a lineage that comprises altogether 136 species placed in 14 genera, distributed from Pakistan and India to Southeast Asia [28]. The Belontiinae subfamily includes a single genus Belontia, which involves two species: B. signata (Günther, 1861) found in Sri Lanka and B. hasselti found in southern Thailand, the Malay Peninsula, Sumatra, Java, and Borneo [29, 30]. Belontia hasselti is specifically adapted to survive in acidic blackwaters and it also represents a commercially valuable fish commonly found in the aquarium trade. The sole cytogenetic investigation undertaken in this species revealed a diploid chromosome number (2n) of 48 almost exclusively acrocentric chromosomes and the presence of a highly heteromorphic ZW sex chromosome system, where the female-limited W chromosome is the only metacentric element in the karyotype [27].
Various repetitive DNA classes have been found accumulated on highly heteromorphic sex chromosomes in teleosts, such as microsatellites (e.g [19, 31–35]), ribosomal RNA genes (e.g [20, 36–38]), transposable elements (e.g [18, 35, 39–41]), telomeric repeats [35], and especially in the recent years also satellite DNAs (satDNA) [42–48]. Repetitive DNAs are therefore intimately associated with sex chromosome evolution and may also directly contribute to master sex-determining gene establishment [49–51] or its relocation to other chromosomal pair in the process referred to as sex chromosome turnover (e.g., in salmonid fishes – reviewed in [52]).
Satellite DNA represents one of the most abundant and variable repetitive DNA fractions in the eukaryotic genomes, both in terms of sequence diversity and patterns of organization across chromosomes. The entire collection of satDNA monomers per species is being termed satellitome and, given its fast evolution, the satellitome properties may vary greatly even between closely related species [53–55]. Among teleosts, the link between satDNAs and sex chromosome evolution has thus far been traced mostly among Neotropical members of the orders Characiformes and Siluriformes ([42, 43, 45, 47, 48, 56, 57] among others) but analogous studies have also been undertaken in African annual killifishes [44, 46]. The satellitomes in the context of ZZ/ZW sex chromosome evolution in Neotropical fishes have been studied thoroughly in the genus Triportheus [43, 57] and Megaleporinus [42, 56]. In both cases, the W chromosomes are heterochromatin-rich and display a high concentration of satDNAs [42, 43, 56, 57].
In this study, we integrated cytogenetic and genomic methods to examine the patterns of evolution and differentiation in the ZW sex chromosome system of B. hasselti. To this end, we characterized the B. hasselti satellitome (being thus the first analyzed perciform species in this respect) and performed chromosomal mapping of the satDNA monomers alongside some other tandemly repeated sequences, namely telomeric repeats and microsatellites. Lastly, we also assessed the degree of sex chromosome differentiation using comparative genomic hybridization (CGH). Our results revealed the lowest satDNA monomer diversity among the teleosts analyzed to date and a low level of ZW sex chromosome differentiation.
Results
Satellitome characterization
The iterations carried out using the Tandem Repeat Analyzer (TAREAN) software resulted in a total of 13 satDNA families for B. hasselti, hereafter named as BhaSatDNAs (Table 1). The vast majority of the satellites found are characterized as long (> 100 bp), with the lengths of their repeat units (monomers) varying between 35 bp and 1797 bp. The A + T content ranged from 38.70 to 68.70% with an average of 51.4%. Divergence ranged from 0.73 to 15.11%, with an average of 6.83% among the satDNAs. The repeat landscapes generated from the analyses are shown in Fig. 1.
Table 1. General features of Belontia. hasselti satellitome, including the monomer sizes in base pairs (bp), abundance, divergence, and A + T contentsatDNA familyMonomer size (bp)AbundanceDivergence (%)A + T (%)BhaSat01-206206 bp0,0817925175,7360,70%BhaSat02-10991099 bp0,0022100174,557,30%BhaSat03-165165 bp0,0010788536,1255,80%BhaSat04-540540 bp0,0009742444,1159,80%BhaSat05-154154 bp0,0007701635,3736,40%BhaSat06-135135 bp0,00047065710,3657,80%BhaSat07-122122 bp0,0003859056,4355,70%BhaSat08-9191 bp0,0002740079,4462,60%BhaSat09-871871 bp0,0002676070,7345,80%BhaSat10-17971797 bp0,0002493891,6453,40%BhaSat11-3535 bp0,00021594615,1154,30%BhaSat12-176176 bp9.01e-0511,9668,70%BhaSat13-106106 bp5.51e-057,3538,70%
Fig. 1. Repeat landscapes showing the abundance and divergence profiles for all satDNA monomers identified in the Belontia hasselti genome
Fluorescence in situ hybridization (FISH)
Only four out of 13 characterized BhaSatDNA monomers displayed positive FISH signals on B. hasselti chromosomes (Fig. 2). The BhaSat01-206 was shown to accumulate in the (peri)centromeric regions of all chromosomes, while the BhaSat04-540 and BhaSat05-154 monomers each located to a single chromosome pair (18 and 5 respectively), with no difference observed between male and female individuals. In contrast, the BhaSat13-106 exhibited sex-linked variation in the distribution pattern, where males possessed signals on both homologs of a single chromosome pair, while females displayed the signal on just one homolog. Though identifying the Z chromosome is challenging, it is nonetheless plausible to infer that this satDNA is associated with the Z chromosome, given this pattern was consistent across all individuals investigated (Fig. 2).
Fig. 2. Metaphase chromosome plates of Belontia hasselti after in situ hybridization experiments with BhaSatDNAs in female (a-d) and male (e-h) individuals. The satDNA family names are indicated in the upper left corner. The arrowhead indicates the W chromosome. Scale bar = 10 μm
C-banding and FISH with telomeric and microsatellite probes
Microsatellite mapping on the chromosomes of B. hasselti resulted in positive signals for the (GA)n and (CGG)n sequences. Detectable (GA)n arrays were scattered across several chromosomes except for the (peri)centromeric regions (Fig. 3c), while the (CGG)n motif was localized in the (peri)centromeric region of only a single chromosome pair (Fig. 3d). Constitutive heterochromatin was detected in all chromosomes, mostly in pericentromeric regions (Fig. 3a). Telomeric sequences were identified at the terminal regions of all chromosomes. No additional (interstitial) sites were observed.
Fig. 3. Female metaphase chromosomes of Belontia hasselti after C-banding (a) and fluorescence in situ hybridization with telomeric (b) and microsatellite sequence motifs (GA)n (c) and (CGG)n (d). The probe used is indicated in the top left corner. The W sex chromosome is marked. Scale bar = 10 μm
Patterns of comparative genomic hybridization (CGH)
Comparative genomic hybridization analysis of male and female individuals of B. hasselti revealed overlapping signals especially in the (peri)centromeric regions of all chromosomes. No regions marked exclusively by either of the genomic probes were observed on metaphases within our sampling (Fig. 4).
Fig. 4. Female metaphase spread of Belontia hasselti after comparative genomic hybridization. Female (red; Atto-550-dUTP-labeled) and male (green; Atto-488-dUTP-labeled) genomic DNA along with DAPI chromosome counterstaining (blue).The merged image with DAPI staining and both genomic probes (indicated in yellow) shows no biased or specific signals of either of the probes. The W chromosome is indicated. Scale bar = 10 μm
Discussion
By analysing the combtail fish B. hasselti, the present study provides the first satellitome analysis in the highly diverse teleost order Perciformes, with a special emphasis on the link between satDNA and sex chromosome evolution in this species. The genome proportion of satDNAs varies considerably among related species in diverse animal and plant lineages [55]. In teleost fishes, the largest satDNA libraries identified to date harbor 140 and 164 different satDNA monomers, presented in the two Anostomidae species, Megaleporinus elongatus (Valenciennes, 1850) and M. macrocephalus (Garavello & Britski, 1988), respectively [42, 58]. On the opposite side of the spectrum, the smallest satellitome to date (25 monomers) has been reported in the Asian arowana Scleropages formosus (Müller & Schlegel, 1840) [59]. In the present study, we provide the new lowest teleost satellitome, composed of only 13 satDNA monomers (BhaSatDNAs) in B. hasselti. The satDNA fraction builds up only 8.86% of the genome of this fish.
Eight out of thirteen BhaSatDNAs did not display visible signals after FISH, most probably due to their non-clustered organization and/or low abundance in the genome, hampering thereby the generation of signals above the method’s detection limit [60–62]. The remaining four BhaSatDNAs (BhaSat01-206, BhaSat04-540, BhaSat05-154, and BhaSat13-106) displayed a range of distinct detectable patterns on chromosomes after FISH (Fig. 2). The most abundant monomer, BhaSat01-206, was detected in the (peri)centromeric regions of all chromosomes (Fig. 2). This pattern together with this monomer being the most abundant satDNA in the B. hasselti genome suggest that this satDNA might play a role in centromere organization and/or function. In teleosts, the prime candidates for centromeric satDNA with these features have been so far identified e.g. in two Neotropical characiform species, Triportheus auritus (Valenciennes, 1850) (TauSat01-119; [43]), and Prochilodus lineatus (Valenciennes, 1837) (PliSat01-225; [63]), and further in African Nothobranchius killifishes [44, 64]. However, further evidence from other Neotropical species strongly suggests that less abundant satDNAs may also take over the centromere-related role [48, 65, 66]. In the case of BhaSat01-206 of B. hasselti, direct experimental testing of the interaction between this satDNA and core centromeric protein CENP-A needs to be undertaken in future studies.
The other two mapped satDNAs, BhaSat04-540 and BhaSat05-154, formed (peri)centromeric and interstitial clusters, respectively, on a single autosome pair (Fig. 2). Interestingly, the last monomer with identifiable signals, BhaSat13-106, presented sex-linked differences in the hybridization pattern, with a signal on both homologs of a single chromosome pair in males but on a single homolog in females. Due to its highly dynamic nature, satDNA evolution is frequently associated with the amplification and contraction of tandem arrays, along with the possible dissemination of the repeats to other genomic locations [54, 55, 60, 67, 68]. It is therefore reasonable to assume that the obtained pattern might reflect the presence/absence polymorphism due to the variable degree of repeat cluster size among homologs, and eventually entire cluster elimination from one of the homologs. The sex-related pattern is puzzling; although our findings align with the hypothesis that this satDNA may represent a marker exclusive to the Z chromosome, the difficulty in identifying the Z chromosome (since it has similar size to the autosomes) using chromosomal markers does not allow us to fully confirm this interpretation.
Teleost species exhibiting a predominantly heteromorphic ZW sex chromosome system typically possess a significantly larger W chromosome compared to its Z counterpart, resulting from considerable accumulation and/or amplification of repetitive DNA, characterized by an extensive block of heterochromatin. A shrunk, largely degenerated W is also present in several lineages, however (reviewed in [7, 69]). Satellitome investigations conducted on the ZW system in fishes indicate fundamental role of this repetitive DNA class in sex chromosome differentiation. In the highly differentiated ZW system of M. macrocephalus, for instance, out of 22 satDNAs evidenced to accumulate on the W chromosome, 14 monomers were exclusively located only on this female-limited sex chromosome [42]. Previous formal analyses based on the updated list of fish sex chromosome systems provided significant support for the hypothesis that, in teleost fishes, female-heterogametic systems generally differentiate faster than male-heterogametic ones [7]. The observed patterns in B. hasselti, however, reveal an unusual case of significantly heteromorphic, enlarged W chromosome, yet characterized by a low degree of heterochromatinization, confined to the pericentromeric region ([27]; present study). Such a discrepancy between apparent heteromorphism (considerably enlarged W chromosome) and absence of large blocks of heterochromatin has been previously described among teleosts in Western mosquitofish Gambusia affinis (Baird & Girard 1853) [69]. More recent study in this fish indicated that despite this fact, the W chromosome is densely populated with microsatellites and transposable elements [70]. Further study is therefore required to expand the analysis of W-linked patterns in B. hasselti by other repetitive DNA classes. Thus far, our CGH assay additionally suggested that qualitative differences in repeat content between Z and W might not be pronounced in B. hasselti, as both male and female genomic probe co-hybridized equally across the chromosome complement, with predominant accumulations in the (peri)centromeric regions only (Fig. 4). These findings imply that either the region of differentiation between Z and W is physically small and cannot be detected at the resolution of CGH, or, alternatively, the lack of sex-specific signal might reflect insufficient differences in the amount and composition of accumulated repetitive DNA across the chromosome complement between sexes (cf. [44, 71]). In summary, together with the low level of constitutive heterochromatin and accumulated repeats evidenced herein and also by Chaiyasan et al. [27], the results indicate a low degree of Z-W sequence divergence in B. hasselti.
Another possible mechanism explaining the occurrence of the enlarged, yet non-heterochromatinized W chromosome might be amplification of gene copies, which creates tandemly repeated segments (amplicons). An increasing number of studies in diverse eukaryotic lineages suggest that this mechanism might represent a common feature accompanying sex chromosome differentiation (e.g [72–75]). A deeper gene content analysis of the W chromosome will determine whether this mechanism might hold also for B. hasselti.
We cannot further exclude the possibility that the addition of another chromosome material (via fusion or translocation event) might enlarge the B. hasselti W chromosome similarly as it was described in certain cichlid fish species of the Oreochromini tribe [76]. The absence of interstitial telomeric signals (ITS) does not corroborate this reasoning, however, it is also probable that telomeric sequences might have been lost prior to chromosome fusion or, alternatively, DNA repair mechanisms might have removed telomeric repeats from the fusion breakpoint.
The evident symmetry of both W chromosome arms regarding their length and the absence of ITS led us to assume yet another and perhaps slightly more plausible scenario for the origin of the W chromosome – through the isochromosome formation. This might have proceeded via aberrant centromere division during mitosis or meiosis, resulting in the separation of chromosome arms rather than chromatids. The symmetric location of repetitive DNA classes, mostly satDNAs, on both chromosome arms of a metacentric chromosome has served as a supporting evidence for this mechanism, proposed previously to originate sex chromosomes in Rumex acetosa [77] and B chromosomes in several Neotropical fish species [65, 78–81]. In favor of this possibility, none of the cytogenetic patterns previously reported for B. hasselti [27] and in the present study show clearly asymmetric distribution between the W chromosome arms. What is more, a slightly supportive pattern has been obtained by mapping the (CA)15 microsatellite motif which tentatively suggests symmetrical distribution across the W chromosome arms [27]. However, given the widespread distribution of this microsatellite throughout the entire chromosome complement, this pattern may equally likely reflect the exclusion of this microsatellite motif from centromeres. Studies involving whole chromosome painting probes and closely related species might provide important data for testing the above hypotheses.
Conclusion
The satellitome of B. hasselti, first one studied within the order Perciformes, turned out to encompass the lowest number of satDNA monomers described in teleosts to date. A subset of satDNA monomers provided detectable signals on the chromosomes and their patterns, together with those from mapping other repetitive sequences and the CGH method, reinforce the view that the ZW sex chromosome system in B. hasselti shows little differentiation at the level of repetitive DNA accumulation (although limitations of our approach have been discussed), despite its highly heteromorphic character with large euchromatic W chromosome. Belontia hasselti thus provides a promising model for investigating alternative hypotheses which could explain the unusual pattern of sex chromosome differentiation in the perciform genome with low satDNA diversity.
Materials and methods
Chromosomal Preparation and C-banding
Ten individuals (five of each sex) of B. hasselti were collected from the To Daeng peat swamp forest in Narathiwat Province, Thailand. Metaphase chromosomes were obtained from kidney using the colchicine treatment as described in [82] and the general protocol following Supiwong et al. [83]. This species is classified as Least Concern (LC) on the IUCN Red List, thus no permissions were necessary for specimen collection under the local legislation. All the procedures followed ethical protocols approved by the Institutional Animal Care and Use Committee of Khon Kaen University, based on the Ethics of Animal Experimentation of the National Research Council of Thailand (ACUC-KKU-90/60). Constitutive heterochromatin distribution was visualized by C-banding according to standard protocol [84].
DNA extraction and genome sequencing
The phenol-chloroform-isoamyl alcohol method [85] was applied to extract genomic DNAs (gDNAs) from female and male individuals. The samples were subsequently sequenced on the BGISEQ-500 platform at BGI (BGI Shenzhen Corporation, Shenzhen, China).
Characterization of Belontia hasselti satellitome
The characterization of B. hasselti satellitome was carried out using the TAREAN software [86], available on the Galaxy platform (https://repeatexplorer-elixir.cerit-sc.cz/galaxy). Firstly, raw reads were quality-filtered using the Trimmomatic software [87] with Q < 20, using the options LEADING:3 TRAILING:3 SLIDINGWINDOW:4:20 MINLEN: 100 CROP: 101 bp parameters. Then, 2 × 500,000 reads were randomly selected for clustering in TAREAN. After this, the putative satellite DNAs identified by TAREAN were filtered from the original library using the DeconSeq software [88] and a new sampling of 2 × 500,000 reads was carried out, with subsequent clustering by TAREAN.
The filtering and re-clustering process was repeated until no putative satellite DNA was recovered by TAREAN. After this, we filtered out multigene families from the catalog of satDNAs. For this, any sequence that exhibited at least 50% similarity to any multigene DNA family (e.g., ribosomal DNA classes) was discarded. In addition, a homology search was carried out using the RepeatMasker software, implemented in a custom Python script (https://github.com/fjruizruano/satminer/blob/master/rm_homology.py) to check for possible redundancies, comparing the similarity between each satDNA sequence. This process used the previously proposed parameters [60], where sequences with over 95% similarity were considered variants of the same satellite, those with between 80% and 95% similarity were considered part of the same satellite DNA family, and those with between 50% and 80% similarity were considered superfamilies. The catalogs were deposited on GenBank with the accession numbers PQ062508–PQ062520.
Abundance and diversity of SatDNAs
The abundance and divergence of B. hasselti satDNA monomers were calculated using the RepeatMasker software [89] using the “cross_match” tool. For this, a sample of 2 × 5,000,000 reads from each library were used. The abundance of each satDNA was obtained by dividing the number of mapped reads by the sum of the total nucleotides. In this way, the satDNAs were ranked in decreasing order of abundance, renamed with the abbreviation of the species name (Bha), together with the term “Sat” followed by the catalog number and the size of the repeated unit (monomer size), as suggested by Ruiz-Ruano et al. [60]. Also, through RepeatMasker, using the Kimura-2 parameter based on the calcDivergenceFromAlign.py script [89], the genetic distance between the BhaSatDNAs was calculated and then represented in a Repeat landscape.
Primer design and DNA amplification by polymerase chain reaction
After isolating the catalog of satellite DNAs, specific primers were manually designed, corresponding to eleven of the thirteen families of satDNAs isolated from B. hasselti. BhaSat11-35, due to its small motif size, was synthesized directly conjugated with Biotin-16-dUTP. As for BhaSat08, due to self-ringing problems, primers were not synthesized. For the eleven satDNA sequences, amplifications were carried out using polymerase chain reaction (PCR), with the reaction conditions and thermal profiles optimized according to Štundlová et al. [44]. The following cycle was used: (i) initial denaturation of 95 ºC for 5 min, (ii) 34 cycles with a denaturation step of 95 ºC for 40 s; an annealing step varying from 52 ºC to 61 ºC for 40 s, depending on the satDNA; and an extension step of 72 ºC for 45 s, (iii) final extension of 72 ºC for 10 min. Once this stage had been completed, the PCR products were checked in a 2% agarose gel to ensure the amplification and integrity of the satDNAs. Finally, these products were quantified using a NanoDrop 2000/2000c spectrophotometer (Thermo Fisher Scientific, Waltham, USA).
Probe labeling and fluorescence in situ hybridization
Once the amplifications had been carried out, the PCR products were labeled using the nick-translation technique via the Jena Bioscience Kit (Jena, Germany), in which the Atto-550-dUTP nucleotide was incorporated into the satDNA sequence according to the manufacturer’s protocol. Microsatellite sequences directly labeled with Cy3 at the 5' end during the synthesis (VBC Biotech, Vienna, Austria) [(GA)n, (TA)n, (GAG)n, (CGG)n, (CAT)n, (TAC)n, (C30)n, (A30)n, (GC)n] and telomeric repeats (TTAGGG)n isolated according to [90] and labeled with Atto-550-dUTP through nick translation (Jena Bioscience) were also used as probes. Fluorescence in situ hybridization experiments were carried out on the chromosomes of females and males of the B. hasselti species following the protocol described in [91].
Assessment of genetic composition of sex chromosomes by comparative genomic hybridization
The genomic DNA of male and female B. hasselti was labeled with Atto-488-dUTP and Atto-550-dUTP, respectively, using the nick-translation kit from Jena Bioscience. Unlabeled C0t-1 DNA, prepared from male gDNA following the protocol of [92], was added to the probe to reduce the hybridization of shared abundant repeats. Thus, the final probe mix for one slide consisted of 3 µg of male-derived C0t-1 DNA and 500 ng of each labeled male- and female-derived gDNAs. After precipitation with 100% ethanol, the probes were air-dried and the pellets resuspended in a hybridization buffer (20 µl per slide), which contained 50% formamide, 2× SSC, and 10% dextran sulfate. The FISH experiments for CGH followed the methodology described in [93].
Microscopy and image analysis
A minimum of 30 metaphase spreads were examined per individual to ascertain the FISH results. Images were taken with a CoolSNAP camera attached to an Olympus BX50 microscope (Olympus Corporation, Ishikawa, Japan), and then processed using Image-Pro Plus 4.1 software from Media Cybernetics (Silver Spring, Maryland, USA).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Charlesworth D. When and how do sex-linked regions become sex chromosomes? Evolution. 2021; 10.1111/evo.1419610.1111/evo.1419633592115 · doi ↗ · pubmed ↗
- 2Abbott JK, Nordén AK, Hansson B. Sex chromosome evolution: historical insights and future perspectives. Proc R Soc B. 2017. 10.1098/rspb.2016.2806.10.1098/rspb.2016.2806 PMC 544393828469017 · doi ↗ · pubmed ↗
- 3Nirchio M, Oliveira C, Cioffi MB, Rossi FMC, Valdiviezo J, Paim FG et al. Occurrence of sex chromosomes in fish of the genus Ancistrus with a new description of multiple sex chromosomes in the Ecuadorian endemic Ancistrus clementinae (Loricariidae). Genes. 2023; 10.3390/genes 1402030610.3390/genes 14020306 PMC 995696036833233 · doi ↗ · pubmed ↗
- 4Plohl M, Meštrović N, Mravinac B. Satellite DNA Evolution. In Genome Dynamics; 2012. pp. 126–152.10.1159/00033712222759817 · doi ↗ · pubmed ↗
- 5Garrido-Ramos MA, Satellite DNA: an evolving topic. Genes. 2017. 10.3390/genes 8090230.10.3390/genes 8090230 PMC 561536328926993 · doi ↗ · pubmed ↗
- 6Müller S, Du K, Guiguen Y, Pichler M, Nakagawa S, Stöck M, et al. Massive expansion of sex-specific SN Ps, transposon-related elements, and neocentromere formation shape the young W-chromosome from the mosquitofish Gambusia affinis. BMC Biol. 2023. 10.1186/s 12915-023-01607-0.10.1186/s 12915-023-01607-0PMC 1018665737189152 · doi ↗ · pubmed ↗
- 7Bolger AM, Lohse M, Usadel B, Trimmomatic. A flexible trimmer for illumina sequence data. Bioinformatics. 2014. 10.1093/bioinformatics/btu 170.10.1093/bioinformatics/btu 170PMC 410359024695404 · doi ↗ · pubmed ↗