ICAT mediates the inhibition of stemness and tumorigenesis in acute myeloid leukemia cells induced by 1,25-(OH)2D3
YULIAN WANG, LIANLI ZHU, RONGHAO ZENG, YUNPING PU, BAIJIAN CHEN, YUWEI TAN, MING HONG, WEIJIA WANG

TL;DR
This study shows that vitamin D inhibits leukemia cell stemness and tumor growth, possibly by interacting with the ICAT protein.
Contribution
The study identifies ICAT as a mediator of 1,25-(OH)2D3's anti-leukemic effects on stemness and tumorigenesis.
Findings
1,25-(OH)2D3 inhibits proliferation and stemness of AML cells and promotes monocyte differentiation.
ICAT knockdown reduces the inhibitory effects of 1,25-(OH)2D3, while ICAT overexpression enhances them.
ICAT modulates the Wnt/β-catenin signaling pathway in response to 1,25-(OH)2D3 in AML cells.
Abstract
The role of 1,25-dihydroxyvitamin D3 (1,25-(OH)2D3) in cancer prevention and treatment is an emerging topic of interest. However, its effects on the stemness of acute myeloid leukemia (AML) cells are poorly understood. The proliferation and differentiation of AML cells (HL60 and NB4) were investigated by the CCK-8 assay, immunocytochemical staining, and flow cytometry. The abilities of HL60 and NB4 cells to form spheres were examined by the cell sphere formation assay. In addition, the levels of stemness-associated markers (SOX2, Nanog, OCT4, and c-Myc) in HL60 and NB4 cells were measured by western blotting and quantitative real-time polymerase chain reaction. Moreover, we obtained β-catenin-interacting protein 1 (ICAT)-knockout and ICAT-overexpressing HL-60 cells using gene editing and lentiviral infection techniques and investigated the role of ICAT in modulating the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
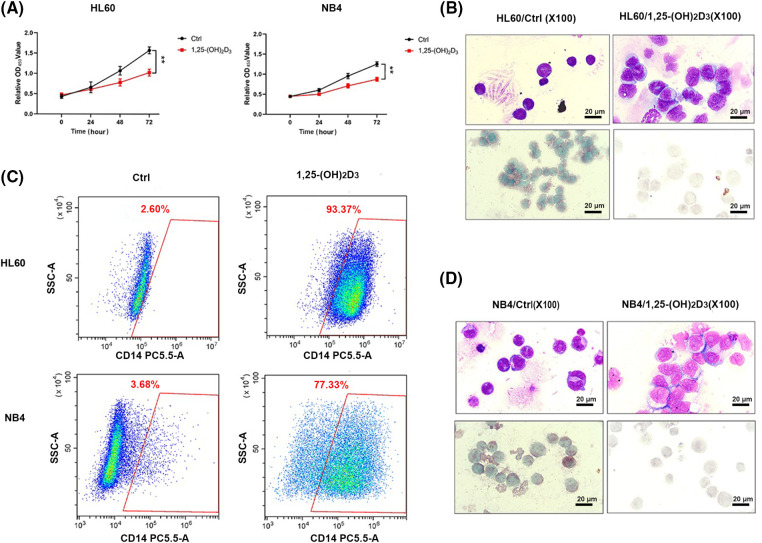 Figure 1
Figure 1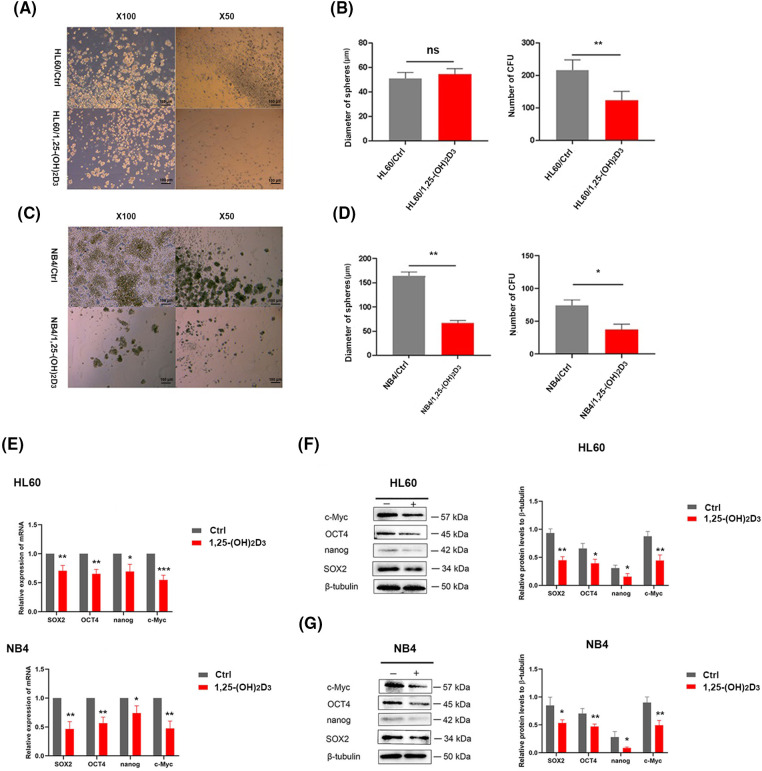 Figure 2
Figure 2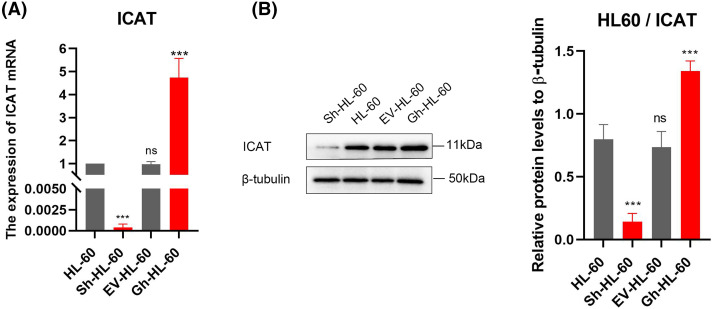 Figure 3
Figure 3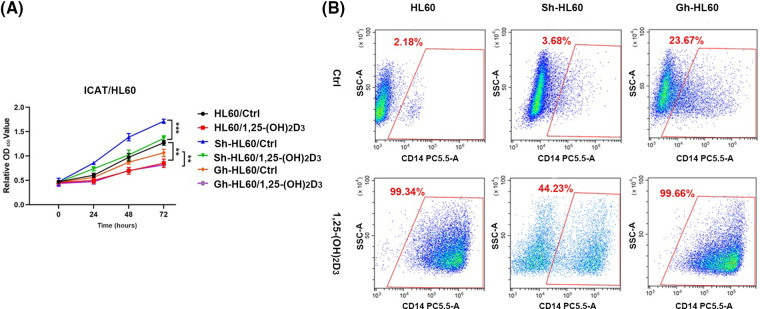 Figure 4
Figure 4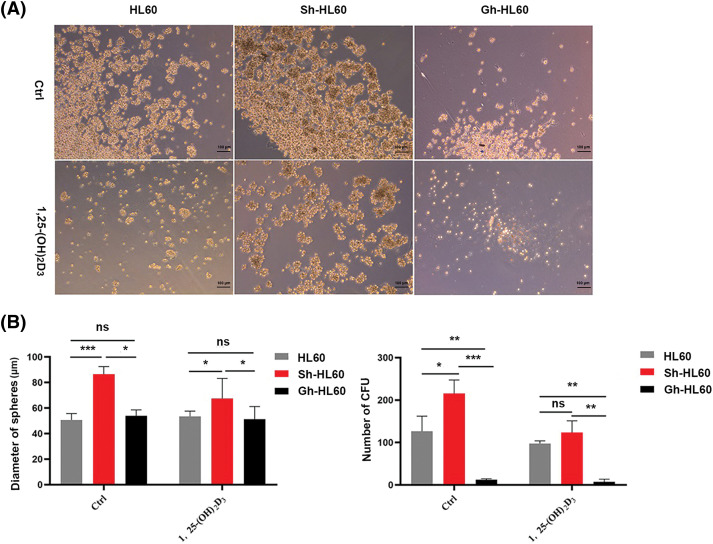 Figure 5
Figure 5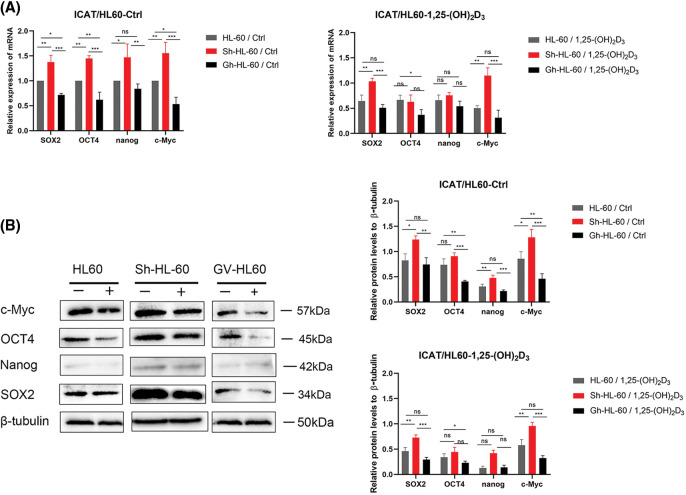 Figure 6
Figure 6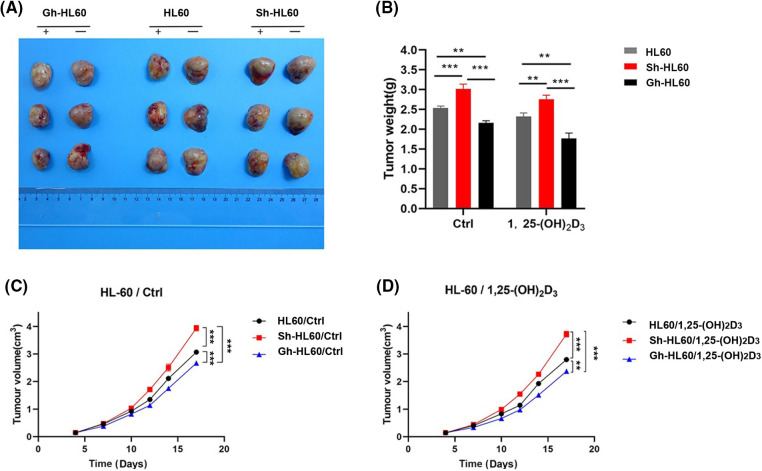 Figure 7
Figure 7| Gene | Primer sequences (5′→3′) |
|---|---|
| OCT4 | F: 5′-GACAGGGGGAGGGGAGGAGCTAGG-3′ |
| R: 5′-CTTCCCTCCAACCAGTTGCCCCAAAC-3′ | |
| nanog | F: 5′-CAGCCCAGATTCTTCCACCAGTCCC-3′ |
| R: 5′-CGGAAGCGTTCCCAGTCGGGTTCACC-3′ | |
| SOX2 | F:5′-GGGAAATGGGAGGGGTGCAAAAGAGG-3′ |
| R: 5′-TTGCGTGAGTGTGGATGGGATTGGTG-3′ | |
| c-Myc | F: 5′-TGGAAAACCAGCCTCCC-3′ |
| R: 5′-CGTAGTCGAGGTCATAGTTC-3′ | |
| GAPDH | F: 5′-AAATCCCATCACCATCTTCCAG-3′ |
| R: 5′-AGGGGCCATCCACAGTCTTCT-3′ |
- —Guangdong Basic and Applied Basic Research Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAcute Myeloid Leukemia Research · Histone Deacetylase Inhibitors Research · Cancer, Hypoxia, and Metabolism
Introduction
Acute myeloid leukemia (AML) is a type of blood and bone marrow cancer characterized by the rapid accumulation of abnormal white blood cells, known as myeloblasts. These cells interfere with the production of normal blood cells, leading to symptoms such as fatigue, infections, and easy bruising or bleeding. AML can affect people of all ages but is more common in older adults [1]. AML comprises a complex group of diseases. The WHO has proposed a classification of acute leukemia that incorporates genetic, epidemiological, and immunologic factors in addition to morphological factors. The latest classification also includes molecular analyses [2]. Current therapeutic strategies for AML include chemotherapy, hematopoietic stem cell transplantation, and immunotherapy [3]. However, conventional chemotherapy often fails to achieve long-term remission, and disease relapse is frequent [4]. Studies have illustrated that stemness, the ability of leukemia cells to self-renew and resist differentiation, is a major driver of primary drug resistance in AML [5]. Addressing both stemness and specific high-risk co-mutations could potentially circumvent resistance and enhance survival outcomes for individuals diagnosed with AML [5]. Therefore, targeting the stemness of leukemia cells is particularly crucial in the context of treatment resistance, prognosis, and relapse in AML.
1,25-(OH)2_D_3, also known as 1α,25-dihydroxyvitamin D_3_, is a multifunctional hormone and a key controller of human genetic activity. It influences the characteristics and functions of different types of cells by regulating the activity of numerous genes in a specific manner depending on the tissue and cell type [6]. Prior studies indicated that 1,25-(OH)2_D_3 can regulate innate and adaptive immunity as well as calcium and bone homeostasis [7–9]. Studies also reported that 1,25-(OH)2_D_3 can inhibit the stemness of cancer cells. Specifically, 1,25-(OH)2_D_3 has inhibitory effects on tumor cell proliferation and promotive effects on the differentiation of tumor cells, including colorectal cancer, melanoma, and leukemia cells [10–12]. In addition, 1,25-(OH)2_D_3 has been demonstrated to attenuate the stemness of tumor cells, including ovarian cancer cells, pancreatic cancer cells, breast cancer cells, and glioblastoma stem-like cells [13–17]. 1,25-(OH)2_D_3 exerts anticancer effects by modulating multiple signaling pathways to inhibit tumor cell stemness. Previous research highlighted the pivotal role of the Wnt/β-catenin signaling in mediating the suppressive effects of 1,25-(OH)2_D_3 on the stemness of breast cancer and ovarian cancer cells [18,19]. In previous research studying the effects of 1,25-(OH)2_D_3 in AML differentiation, 1,25-(OH)2_D_3 was found to inhibit the entry of β-catenin protein into the nucleus by upregulating β-catenin–interacting protein 1 (ICAT) and then suppressing the Wnt/β-catenin signaling, inducing the differentiation of acute promyelocytic leukemia cells into a mononuclear line, with ICAT playing a pivotal role in this process [20]. Earlier investigations revealed that ICAT inhibits glioblastoma proliferation and suppresses colorectal cancer progression by targeting the Wnt/β-catenin signaling [21,22]. Nevertheless, the precise mechanism by which 1,25-(OH)2_D_3 and ICAT affect AML cell stemness remains largely unknown.
In this study, HL-60 and NB4 cells underwent treatment with 1,25(OH)2_D_3 to assess its impact on cell proliferation, differentiation, and the expression of markers associated with stemness. The investigation aimed to uncover how 1,25(OH)2_D_3 influences the stem cell characteristics of AML cells. Our data demonstrated that 1,25(OH)2_D_3 inhibited the stemness of HL-60 and NB4 cells, providing evidence supporting the use of 1,25-(OH)2_D_3 in leukemia treatment. Additionally, the experimental data from prior investigations revealed that ICAT protein expression was increased in HL-60 cells following exposure to 1,25(OH)2_D_3. Leveraging CRISPR/Cas9 gene-editing technology alongside lentivirus transfection, we successfully engineered HL-60 cell models featuring ICAT knockout or overexpression. Subsequently, we demonstrated that 1,25(OH)2_D_3 suppresses stemness through the Wnt/β-catenin signaling via a mechanism mediated by ICAT. This provides novel insights into the potential mechanisms of current antitumor therapy in leukemia.
Materials and Methods
Cell culture
NB4 cells were purchased from Wuxi Newgain Biotechnology Co., Ltd. (Wuxi, China), and HL-60 cells were procured from the Chinese Academy of Sciences. HL60 cells were cultured in IMDM medium (SH30228.01, Cytiva, CO, USA), with 20% fetal bovine serum (Gibco, Grand Island, USA) and 1% penicillin-streptomycin (Gibco, Grand Island, USA). NB4 cells were cultured in RPMI-1640 medium (CGM112.05, Cellmax, Guangzhou, China) with 10% fetal bovine serum (Gibco, Grand Island, USA) and 1% penicillin-streptomycin (Gibco, Grand Island, USA). All cells were maintained in a 37°C, 5% CO_2_ cell culture incubator. 1,25-(OH)2_D_3 (D1530, Sigma-Aldrich, St. Louis, MO, USA) was dissolved in absolute ethyl alcohol and stored at −20°C at a concentration of 1 × 10^−3^ M/L. The concentration of 1,25-(OH)2_D_3 in the drug medium was 1 × 10^−7^ M/L.
CCK-8 assay
Cells were seeded at a density of 10^4^ cells in 96-well plates. Cells in the drug group were incubated with 1 × 10^−7^ M 1,25-(OH)2_D_3, and cells in the control group were untreated. All other conditions were identical between the groups. Twenty microliters of CCK-8 reagent (Beyotime, Shanghai, China) were added to each well at the specified time point (0, 1 day, 2 day, and 3 day), followed by incubation for 2.5 h, and the absorbance at 450 nm was measured in a spectrophotometer (Bio-Rad, Hercules, CA, USA) to calculate the relative growth rate of the cells.
Immunohistochemical assay
Cells from both the control and experimental groups were collected at 72 h for Wright–Giemsa staining (BioVision, Guangzhou, China). Esterase staining was performed using the Acid α-Naphthyl Acetate Esterase Staining Kit (G2390, Solarbio, Guangzhou, China) [23], and NaF inhibition assays were conducted to observe cellular morphology using a BX-63 upright microscope (Olympus, Tokyo, Japan).
CD14 detection by flow cytometry
Approximately 2 × 10^5^/mL cells were exposed to 1,25-(OH)2_D_3 (10^−7^ M, as a positive control) for 72 h. Cells were washed for twice by pre-cold 1× PBS. Then, incubated with anti-CD14 antibody (Becton Dickinson, NJ, USA) in the dark. After 30 min, cells were washed by pre-cold PBS and detected with flow cytometry (CytoFLEX S, Beckman-Coulter, CA, USA) [24].
Cell sphere formation assay
First, 0.5 × 10^3^ cells were seeded into ultra-low attachment 24-well plates. Starting from day 0, 200 µL of sphere-specific culture medium was added, and 200 µl of sphere-specific culture medium was added again on days 3, 6, 9, and 12. Photographic documentation and quantification of tumor spheres (diameter > 30 μm) were conducted on days 9, 12, and 15 [25].
Western blot blotting
Total proteins was extracted using RIPA lysis buffer (C1053, Applygen, Beijing, China) containing protease inhibitors (4693132001, Roche, Basel, Switzerland), and the protein concentration was determined by the bicinchoninic acid assay (BIV-K812-1000, BioVision). Proteins (30 µg/well) were separated by 12% SDS-PAGE and then transferred to a PVDF membrane (ISEQ00010, Millipore, Danvers, MA, USA). Membranes were sealed with 5% skim milk for 0.5 h and incubated with the following primary antibodies at 4°C overnight: anti-β-tubulin (AF1216, 1:5000, Beyotime), anti-SOX2 (ab97959, 1:2000, Abcam, Cambridge, UK), anti-OCT4 (ab179800, 1:2000, Abcam), anti-Nanog (4903S, 1:2000, CST, Danvers, MA, USA), and anti-c-Myc (GTX10825, 1:1000, GeneTex, SoCal, UK). Subsequently, the membranes were incubated for 1.5 h at 24°C with Alexa Fluor 488B-labeled anti-rabbit IgG (ab6721, 1:5000, Abcam). Immobilon Western Chemiluminescent HRP Substrate (WBKLS0500, Millipore, MA, USA) was used to detect the blots using a chemiluminescence imager (Fusion SoloS. EDGE, VILBER, Paris, France). The intensity was analyzed using Image Lab™ software (Bio-Rad).
Quantitative real-time polymerase chain reaction (qRT-PCR)
The experimental and control groups of cells were subjected to total RNA extraction with TRIzol reagent (15596026, Life Technologies, South San Francisco, CA, USA). Subsequently, cDNA synthesis was performed following the manufacturer’s instructions using the HiScript® III RT SuperMix for qPCR (+gDNA wiper) kit (R223-01, Vazyme, Nanjing, China). The sequences of primers are shown in Table 1. qRT-PCR was conducted using PowerUp™ SYBR™ Green Master Mix (A25742, Applied Biosystems, Waltham, MA, USA) and the ABI PCR System (Applied Biosystems). The thermocycling parameters were as follows: 50°C for 120 s; 95°C for 120 s; and 40 cycles of 95°C for 15 s and 60°C for 60 s. The dissolution curve protocol was 95°C for 15 s, 60°C for 60 s, and 95°C for 15 s. Relative gene expression levels were calculated by the 2^−ΔΔCq^ method [26]. GAPDH was applied as a normalization control.
CRISPR/Cas9 gene-editing and lentivirus infection techniques
The human CTNNBIP1 (ICAT) gene was knocked out in HL-60 cells using CRISPR/Cas9 [27]. Homozygous ICAT gene knockout (Sh-HL-60) was confirmed by PCR and by sequencing electroporated single clones. The lentiviral vector LVEFS>KozakHumanCTNNBIP1CDS[NM_020248.3]CMV>EGFP/T2A/Puro was constructed, validated by PCR, and sequenced. 293T cells were infected with ICAT lentivirus, a control vector, and a helper plasmid for lentivirus packaging. The ICAT overexpression virus and control virus were then used to infect HL-60 cells, resulting in stably transfected ICAT-overexpressing cells (Gh-HL-60) and empty vector cells (EV-HL-60), as determined by screening with puromycin. Harvested cells were used for subsequent experiments.
In vivo tumorigenesis assay
The animal experiments in the current study were approved and supervised by the Institutional Animal Care and Use Committee of Zhongshan People’s Hospital (No. K-2023-214). 54 (eighteen in each group) nude mice (6-week-old male NOD/SCID), which were purchased from GemPharmatech Co., Ltd., Nanjing, China, were maintained in pathogen-free facilities. All the animals were weighed and randomly into groups according to their weight using StudyDirector™ (3.1.399.19, Studylog System, Inc., S. San Francisco, CA, USA) Select the Matched distribution method for grouping. Approximately 106 Sh-HL60, Gh-HL60, and HL60 cells were injected subcutaneously into the right flank of mice (eighteen mice in each group) for tumorigenesis. When the average tumor volume reached 100 mm^3^, mice were randomly assigned to two groups with 3 mice per group: the treatment (1,25-(OH)2_D_3) and control (PBS) groups. Mice in the treatment group received 0.5 μg/kg 1,25-(OH)2_D_3 intraperitoneally twice a day for 2 weeks. Tumor size was measured twice a week. Tumour volumes were calculated according to the formula (width2 × length)/2. The mice were humanely killed, and the weight of the excised tumor was measured.
Statistical analysis
All data in this study were generated from three independent experiments. Statistical analyses were conducted with the GraphPad Prism 7.0 software. Differences between the two groups were assessed by the two-tailed Student’s t-test. Differences between before and after treatment were assessed by one-way analysis of variance (ANOVA). p < 0.05 indicated statistical significance.
Results
Antiproliferative and monocytic differentiation-inducing effects of 1,25-(OH)2D3 I AML cells
We used the CCK-8 assay to validate the suppressive effects of 1,25-(OH)2_D_3 on the proliferation of HL-60 and NB4 cells (Fig. 1A). We also employed an immunohistochemical assay to elucidate the monocytic lineage differentiation induced by 1,25-(OH)2_D_3 in NB4 and HL-60 cells (Fig. 1B,D). The expression of the monocytic differentiation marker CD14 was remarkably upregulated after treatment (Fig. 1C). The results indicated that 1,25-(OH)2_D_3 significantly inhibits cell proliferation in HL-60 and NB4 cells while inducing their differentiation toward the monocytic lineage.
*Antiproliferative and monocytic differentiation-inducing effects of 1,25-(OH)2D3 on AML cells. (A) The proliferation of HL-60 and NB4 cells following 1,25-(OH)2D3 treatment for 0, 24, 48, and 72 h. The results are presented as the mean ± SD of triplicate experiments. *p < 0.01. (B) Representative images of Wright–Giemsa staining, esterase staining, and NaF inhibition experiments in untreated and drug-treated HL-60 cells. (C) Representative flow cytometry data depicting the percentage of cells that were positive for the surface antigen CD14 in untreated and drug-treated HL-60 and NB4 cells. (D) Representative images of Wright–Giemsa staining, esterase staining, and NaF inhibition experiments in untreated and drug-treated NB4 cells.
Inhibition of stemness by 1,25-(OH)2D3 in AML cells
We also conducted a cell sphere formation assay to further determine the effects of 1,25-(OH)2_D_3 on the stemness of HL-60 and NB4 cells. 1,25-(OH)2_D_3 suppressed sphere formation by these cells, as evidenced by reductions in the number (Fig. 2A,B) and diameter of spheres (Fig. 2C,D). Regarding stemness marker gene expression, the expression of OCT4, Nanog, SOX2, and c-Myc was inhibited by 1,25-(OH)2_D_3 at the protein (Fig. 2E) and mRNA levels (Fig. 2F,G). These results indicated that 1,25-(OH)2_D_3 significantly decreased the stemness of HL-60 and NB4 leukemia cells.
*Inhibition of stemness by 1,25-(OH)2D3 in AML cells. (A) Stemness of HL-60 cells treated with or without 1,25-(OH)2D3 (Ctrl) was analyzed by the sphere formation assay (×50 and ×100 magnification). (B) Quantification and statistical analysis of the data in A. (C) Stemness of NB4 cells treated with or without 1,25-(OH)2D3 (Ctrl) was analyzed by the sphere formation assay (×50 and ×100 magnification). (D) Quantification and statistical analysis of the data in C. (E) qRT-PCR of SOX2, OCT4, Nanog, and c-Myc expression. (F) Protein expression of SOX2, OCT4, Nanog, and c-Myc after treatment with or without 1,25-(OH)2D3 for 72 h in HL-60 cells was analyzed by western blotting. (G) Protein expression of OCT4, SOX2, c-Myc, and Nanog after treatment with or without 1,25-(OH)2D3 for 72 h in NB4 cells was verified by western blot. nsp > 0.05, *p < 0.05, **p < 0.01, **p < 0.001.
Validation of stable HL-60 strains with ICAT gene knockout or overexpression
HL-60 cells stably transfected with ICAT gene knockout and ICAT gene overexpression constructs were successfully generated and verified by qRT-PCR and western blotting, as presented in Fig. 3A,B. The sequencing results of ICAT gene knockout and overexpression vector construction are provided in the Appendix A (Fig. A1 and Table A1).
*Stable HL-60 cells were identified with both ICAT gene knockout and overexpression. (A) Quantitative real-time PCR was conducted to assess ICAT expression in these stable HL-60 transfectants. (B) Western blotting analysis was performed to examine ICAT expression in the stable HL-60 cells. Results represent the mean ± SD of triplicate experiments. nsp > 0.05, **p < 0.001.
Role of ICAT in the 1,25-(OH)2D3-mediated inhibition of AML cells proliferation and induction of monocytic differentiation
The results of the CCK-8 assay indicated 1,25-(OH)2_D_3 inhibited the proliferation of HL-60 cells, and this effect was lessened by ICAT knockout (Fig. 4A). Flow cytometry revealed 1,25-(OH)2_D_3 induced mononuclear differentiation, and this effect was weakened by ICAT knockout and increased by more than 20% by ICAT overexpression (Fig. 4B). These findings indicate that 1,25-(OH)2_D_3 may influence HL-60 cell differentiation through ICAT expression.
*ICAT expression’s involvement in the inhibition of AML cell proliferation and the promotion of monocytic differentiation by 1,25-(OH)2D3 was investigated. (A) HL-60, Sh-HL-60, and Gh-HL-60 cell proliferation was monitored at 0, 24, 48, and 72 h post 1,25-(OH)2D3 treatment. Data represent the mean ± SD of triplicate experiments. **p < 0.01, **p < 0.001. (B) Flow cytometry analysis demonstrated the percentage of CD14-positive cells among treated and untreated HL-60, Sh-HL-60, and Gh-HL-60 cells.
Enhancement of the 1,25-(OH)2D3-induced inhibition of AML cells sphere formation by ICAT expression
We performed cell sphere formation experiments to examine the role of ICAT expression in the inhibition of HL-60 cell sphere formation by 1,25-(OH)2_D_3. Our results indicated that, in untreated control cells, knockdown of ICAT alone enhanced sphere formation by HL-60 cells, whereas overexpression of ICAT alone inhibited their spheroidization. Meanwhile, in cells incubated with 1,25-(OH)2_D_3, ICAT overexpression enhanced its inhibitory effect, whereas 1,25-(OH)2_D_3 could not effectively inhibit the spheroidization of HL-60 cells after ICAT knockout, as evidenced by the changes in the number and diameter of spheroids (Fig. 5A,B).
*ICAT expression enhanced the inhibitory effect of 1,25-(OH)2D3 on AML cell sphere formation. (A) Stemness of HL-60, Sh-HL-60, and Gh-HL-60 cells treated with or without 1,25-(OH)2D3 (Ctrl) was analyzed by the sphere formation assay (×100 magnification). (B) Quantification and statistical analysis of the data in A. Results represent the mean ± SD of triplicate experiments. nsp > 0.05, *p < 0.05, **p < 0.01, **p < 0.001.
Downregulation of stemness markers by 1,25-(OH)2D3 through ICAT expression in AML cells
We assessed stemness-related gene expression (SOX2, Nanog, OCT4, c-Myc) in AML cells using qRT-PCR. ICAT knockout lessened the inhibitory effects of 1,25-(OH)2_D_3 on SOX2 and c-Myc expression, whereas these effects were enhanced by ICAT overexpression. In addition, ICAT overexpression enhanced the effects of 1,25-(OH)2_D_3 on OCT4 expression (Fig. 6A). The results of western blotting aligned with those of qRT-PCR (Fig. 6B), indicating that ICAT promoted the downregulation of stemness markers in AML cells treated with 1,25-(OH)2_D_3. This suggests that 1,25-(OH)2_D_3 inhibits AML cell stemness by modulating ICAT, and ICAT alone can suppress stemness characteristics.
*Downregulation of stem cell markers by 1,25-(OH)2D3 through ICAT expression in AML cells. (A) qRT-PCR of SOX2, OCT4, Nanog, and c-Myc expression in HL-60, Sh-HL-60, and Gh-HL-60 cells after treatment with or without 1,25-(OH)2D3 for 72 h. (B) Protein expression of SOX2, OCT4, Nanog, and c-Myc after treatment with or without 1,25-(OH)2D3 for 72 h in HL-60, Sh-HL-60, and Gh-HL-60 cells was analyzed by western blotting. Results represent the mean ± SD of triplicate experiments. nsp > 0.05, *p < 0.05, **p < 0.01, **p < 0.001.
Inhibition of xenograft growth by 1,25-(OH)2D3 and ICAT expression in vivo
We conducted an in vivo tumor growth study using HL-60, Sh-HL-60, and Gh-HL-60 xenograft mouse models. 1,25-(OH)2_D_3 treatment significantly reduced tumor size and weight in mice. In the untreated control group, overexpression of ICAT reduced the volume and weight of xenograft tumors in the HL-60 model, whereas knockdown of ICAT increased tumor volume and weight (Fig. 7A–C). In the 1,25-(OH)2_D_3 treatment group, overexpression of ICAT enhanced the tumor-suppressing effect of 1,25-(OH)2_D_3, whereas this effect was attenuated by ICAT knockout (Fig. 7B,D).
*Inhibition of xenograft tumor by 1,25-(OH)2D3 and ICAT expression in vivo. (A) Photographs of tumors xenografts. (B) The effects of 1,25-(OH)2D3 and ICAT expression on tumor weight. (C) The corresponding tumor volumes in the untreated group at the indicated times are presented. (D) The corresponding tumor volumes after 1,25-(OH)2D3 injection on the indicated days. **p < 0.01, **p < 0.001.
Discussion
Cell stemness is defined by its quiescent state, pluripotent nature, and ability for long-term self-renewal [22]. It plays critical roles in leukemia initiation, progression, and relapse [28], but the key determinants of leukemia cell stemness are poorly understood [29]. Therefore, targeting leukemia cell stemness could be a promising strategy for improving the efficacy of leukemia treatment [30]. 1,25-(OH)2_D_3 is the most biologically active vitamin D metabolite [31]. Upon binding with its receptor, it initiates the translocation of the receptor complex to the nucleus, where it interacts with the genome, regulating the expression of over 1200 genes [32]. Studies have illustrated that immune cells, such as monocytes, dendritic cells, lymphocytes, and macrophages, express the vitamin D receptor. 1,25-(OH)2_D_3 can activate immune cells, such as T and B cells, macrophages, and dendritic cells, to regulate immunity and increase the production of antimicrobial peptides and neutralizing antibodies [33–35]. In addition, studies have revealed that 1,25-(OH)2_D_3 acts as an efficient anticancer agent through several signaling pathways [36–40]. Recent data suggest that 1,25-(OH)2_D_3 plays a regulatory function in both normal and cancerous stem cells, exerting an inhibitory effect on stem-like properties across diverse tumor types including hepatocellular carcinoma, breast, colorectal, prostate, and gastric cancer cells [36,41–45]. However, the effects of 1,25-(OH)2_D_3 on the stemness of AML cells has not been well studied. In this study, we investigated the effects of 1,25-(OH)2_D_3 on the stemness of AML cells. The experimental results provided evidence that 1,25-(OH)2_D_3 may directly inhibit the stemness of AML cells, and the compound suppressed the proliferation of NB4 and HL-60 cells and induced their differentiation into monocytes. This is consistent with previous studies suggesting that 1,25-(OH)2_D_3 can stimulate myeloid stem cells to preferentially differentiate into monocytes and macrophages [46]. In addition, the present results revealed that 1,25-(OH)2_D_3 can also suppress sphere formation by HL-60 and NB4 cells and suppress the expression of the stemness markers SOX2, Nanog, OCT4, and c-Myc in HL-60 and NB4 cells. The results indicated that 1,25-(OH)2_D_3 can inhibit the stemness of AML cells, as previously observed in breast cancer, colorectal cancer, and other tumors.
In our previous studies examining the role of ICAT in AML differentiation, we found that the entry of β-catenin protein into the nucleus was inhibited by upregulating ICAT expression, and then inhibition of the Wnt/β-catenin signaling pathway induced the differentiation of HL-60 acute promyelocytic leukemia cells into a mononuclear line [20]. ICAT is a 9-kDa polypeptide that plays a negative role in the Wnt/β-catenin signaling by inhibiting β-catenin nuclear signaling via binding to β-catenin and competing with the transcription factor T cell factor [47,48]. ICAT is a key protein in the Wnt/β-catenin signaling that negatively modulates β-catenin co-transcriptional activity [48,49]. As an important molecule of the Wnt signaling pathway, ICAT sustains stemness by maintaining multipotency in certain cell types [50]. Activation of the Wnt/β-catenin signaling is one of the important mechanisms of tumorigenesis [51]. To further elucidate the function of ICAT in the 1,25-(OH)2_D_3-mediated inhibition of AML cell stemness, we knocked out and overexpressed the ICAT gene in HL-60 cells and analyzed its role by western blotting, qRT-PCR, and cell sphere formation assays. In untreated cells, knockout of ICAT alone enhanced the stemness of HL-60 cells, while upregulation of ICAT inhibited the stemness of these cells. In the 1,25-(OH)2_D_3 treatment group, ICAT overexpression enhanced the stemness-inhibiting effect of 1,25-(OH)2_D_3 in AML cells, whereas ICAT knockout weakened this inhibitory effect. These results suggested that 1,25-(OH)2_D_3 suppresses stemness through the Wnt/β-catenin signaling through a mechanism mediated by ICAT protein. Moreover, we validated these results in vivo in NOD/SCID mice. Previous studies illustrated that 1,25-(OH)2_D_3 suppresses cancer cells proliferation and promotes their differentiation, primarily by antagonizing TGF-β, Wnt/β-catenin, and EGF signaling pathways [6]. However, 1,25-(OH)2_D_3 interacts with the vitamin D receptor which binds to specific DNA sequences known as vitamin D response elements, thereby controlling the expression of interconnected downstream genes [52].
Recent research has shown that decreased ICAT expression correlates with unfavorable disease-free and overall survival outcomes in AML, indicating that ICAT is closely involved in AML progression [50]. In the current study, our initial focus was on understanding the role of ICAT in the suppression of AML cell stemness by 1,25-(OH)2_D_3. Our findings revealed that 1,25-(OH)2_D_3 reduces AML cell stemness by targeting the Wnt/β-catenin signaling, a process facilitated by ICAT protein. Nevertheless, there were certain limitations in our research. Notably, while the dosage of 1,25-(OH)2_D_3 administered to nude mice was based on existing literature, the delay in tumor extraction resulted in larger tumor sizes.
Conclusions
In summary, our study revealed that 1,25-(OH)2_D_3 suppresses the stem-like properties of AML cells, likely by modulating the Wnt/β-catenin signaling via ICAT. These findings offer fresh insights into how 1,25-(OH)2_D_3 may exert its anti-leukemic effects. Additionally, our observations suggest that ICAT plays a role in curtailing the stemness of AML cells, with its expression tightly linked to this characteristic. Therefore, further studies on the role and regulation of ICAT might reveal a novel therapeutic target for overcoming treatment resistance, improving prognosis, and preventing relapse in patients with AML.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Thomas GE, Egan G, García-Prat L, Botham A, Voisin V, Patel PS, et al. The metabolic enzyme hexokinase 2 localizes to the nucleus in AML and normal haematopoietic stem and progenitor cells to maintain stemness. Nat Cell Biol. 2022;24(6):872–84. doi:10.1038/s 41556-022-00925-9; 35668135 PMC 9203277 · doi ↗ · pubmed ↗
- 2Arber DA, Orazi A, Hasserjian R, Thiele J, Borowitz MJ, Le Beau MM, et al. The 2016 revision to the World Health Organization classification of myeloid neoplasms and acute leukemia. Blood. 2016;127(20):2391–405. doi:10.1182/blood-2016-03-643544; 27069254 · doi ↗ · pubmed ↗
- 3Thol F, Ganser A. Treatment of relapsed acute myeloid leukemia. Curr Treat Options Oncol. 2020;21(8):66–76. doi:10.1007/s 11864-020-00765-5; 32601974 PMC 7324428 · doi ↗ · pubmed ↗
- 4Mattes K, Gerritsen M, Folkerts H, Geugien M, van den Heuvel FA, Svendsen AF, et al. CD 34+ acute myeloid leukemia cells with low levels of reactive oxygen species show increased expression of stemness genes and can be targeted by the BCL 2 inhibitor venetoclax. Haematologica. 2020;105(8):e 399–e 403. doi:10.3324/haematol.2019.229997; 31727766 PMC 7395293 · doi ↗ · pubmed ↗
- 5Wang F, Morita K, Di Nardo CD, Furudate K, Tanaka T, Yan Y, et al. Leukemia stemness and co-occurring mutations drive resistance to IDH inhibitors in acute myeloid leukemia. Nat Commun. 2021;12(1):2607. doi:10.1038/s 41467-021-22874-x; 33972549 PMC 8110775 · doi ↗ · pubmed ↗
- 6Fernández-Barral A, Bustamante-Madrid P, Ferrer-Mayorga G, Barbáchano A, Larriba MJ, Muñoz A. Vitamin D effects on cell differentiation and stemness in cancer. Cancers. 2020;12(9):2413–31. doi:10.3390/cancers 12092413; 32854355 PMC 7563562 · doi ↗ · pubmed ↗
- 7Bikle DD. Vitamin D regulation of immune function. Curr Osteoporos Rep. 2022;20(3):186–93. doi:10.1007/s 11914-022-00732-z; 35507293 PMC 9065668 · doi ↗ · pubmed ↗
- 8Sîrbe C, Rednic S, Grama A, Pop TL. An update on the effects of vitamin D on the immune system and autoimmune diseases. Int J Mol Sci. 2022;23(17):9784–812. doi:10.3390/ijms 23179784; 36077185 PMC 9456003 · doi ↗ · pubmed ↗