Hidden diversity of Trypanosomatidae (Protozoa: Kinetoplastea) in bats from an urban park in Brazil
Mariana Alves Lima, Marcela Elisa Vaz, Jennifer Emanuelle Ferreira, Ana Cristina Vianna Mariano da Rocha Lima, Débora Cristina Capucci, Sônia Aparecida Talamoni, Felipe Dutra-Rêgo, José Dilermando Andrade Filho, Vinícius Belo, Vinícius Belo, Vinícius Belo, Vinícius Belo

TL;DR
This study explores the diversity of trypanosomatid parasites in bats from an urban park in Brazil, revealing new insights into their potential role in disease transmission.
Contribution
The study identifies novel trypanosomatid species and mixed infections in urban bats, expanding understanding of their ecological and epidemiological significance.
Findings
Nine bats were found to carry trypanosomatids, including Leishmania infantum and new Trypanosoma species.
Mixed infections with Leishmania infantum and Leishmania braziliensis were detected in one bat species.
DNA integrity was confirmed using cytb and gamma-actin gene amplification.
Abstract
Trypanosomatids are obligate flagellated parasites, with the genera Leishmania and Trypanosoma acting as etiological agents of significant diseases such as leishmaniasis and Chagas disease. Although ecological studies have increasingly highlighted the role of bats as potential reservoirs of these parasites, the diversity of trypanosomatids in urban bat populations remains poorly understood. This study investigates the occurrence and diversity of Trypanosomatidae in bats from Mangabeiras Municipal Park (MMP), an urban park in Belo Horizonte, Minas Gerais, Brazil, a region of ecological interest due to the prior detection of Leishmania in sand flies. A total of 56 bats representing seven species were captured, and 149 biological samples (blood, tissues, and organs) were analyzed using NNN/LIT culture medium. Contamination was reported in 32.2% of the samples, while 67.8% yielded negative…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Fig 1
Fig 1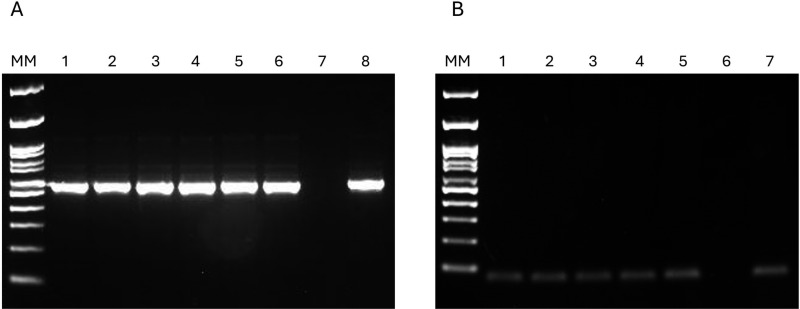 Fig 2
Fig 2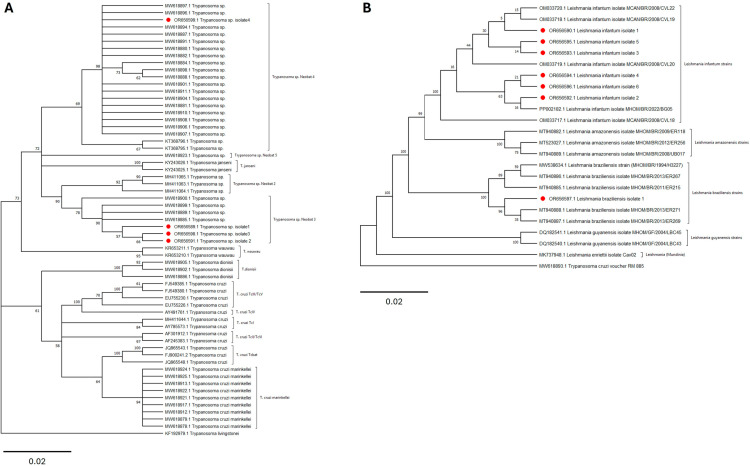 Fig 3
Fig 3- —http://dx.doi.org/10.13039/501100003593Conselho Nacional de Desenvolvimento Científico e Tecnológico
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTrypanosoma species research and implications · Vector-borne infectious diseases · Research on Leishmaniasis Studies
Introduction
The order Trypanosomatida comprises obligate flagellated endoparasites, with the genera Leishmania and Trypanosoma being particularly noteworthy. Leishmania is responsible for visceral (VL) and cutaneous leishmaniasis (CL), while Trypanosoma includes parasites associated with Chagas disease in the Americas (Trypanosoma cruzi) and sleeping sickness in Africa (Trypanosoma brucei) [1]. Trypanosomatids are transmitted to vertebrate hosts through various mechanisms. Leishmania is transmitted via the bite of sand flies during blood feeding [2], while T. cruzi is primarily transmitted through the release of triatomine feces and urine (Hemiptera: Reduviidae: Triatominae) during hematophagy, although other transmission routes have also been reported [3]. Trypanosomatids are among the most extensively studied protists and, based on their life cycle, can be subdivided into two non-taxonomic groups: monoxenous (involving a single host) and heteroxenous (involving two or more hosts, one of which is a vector). The phylogenomic structure of these parasites, typically analyzed through multiple protein-coding genes, has revealed a complex and heterogeneous relationship among the genera [4].
In general, trypanosomatids exhibit ecological associations with a wide range of hosts, including humans [5]. However, enzootic cycles associated with these pathogens are primarily linked to mammals in wild areas [6,7]. The high susceptibility of bats to infection by various trypanosomatids has been extensively studied [8,9], and in some cases, bats have been suggested as potential reservoirs, particularly for protozoa of the genus Trypanosoma and possibly Leishmania [8,10,11]. Given the diversity and potential for mixed infections in bat populations, the use of sensitive and specific detection methods, such as PCR, is essential for analyzing biological samples that may present low and heterogeneous parasite loads [12–14]. The significance of these animals as natural hosts for various pathogens is further amplified by their ecological habits, which span vast geographical regions and their adaptability to diverse environments, including urban areas [15], potentially facilitating transmission, particularly to humans.
Despite growing evidence of trypanosomatid infections in bats, most studies have focused on rural or sylvatic settings [14,16,17], with limited investigations into urban environments where bats frequently interact with humans and domestic animals. Recent studies have increasingly underscored the role of urban areas in harboring diverse trypanosomatid species, both in endemic and non-endemic regions, providing critical insights for guiding health authorities in epidemiological surveillance efforts [13,18]. Urban parks, in particular, may serve as hotspots for pathogen transmission due to the overlap of human, domestic, and wildlife populations. This study investigates the occurrence and diversity of trypanosomatids in bats from Mangabeiras Municipal Park (MMP), an urban park in Belo Horizonte, Minas Gerais, Brazil, providing insights into the role of urban environments in shaping the dynamics of these parasites.
Previous studies have reported the presence of Leishmania in sand flies in this park, suggesting a potential role of urban protected areas in maintaining trypanosomatid transmission cycles [19]. Belo Horizonte is endemic for CL and VL, with 416 human cases of VL and 663 cases of CL reported between 2020 and 2024 [20]. These findings highlight the need to investigate the role of bats in urban trypanosomatid transmission cycles and compare these dynamics with rural and sylvatic scenarios observed in other regions of Brazil and worldwide.
Materials and methods
Study area
Mangabeiras Municipal Park (MMP), located in the south-central region of Belo Horizonte, Minas Gerais, is considered one of the largest urban parks in Latin America, covering an area of 245.2 hectares. According to the Köppen-Geiger climate classification, the region has a subtropical highland climate (Cwb), characterized as humid temperate with dry winters [21]. MMP primarily features Atlantic Forest vegetation, with areas of Cerrado, especially on the slopes, fostering rich biodiversity. Recent studies have assessed the presence of hemoparasites and ectoparasites in Nasua nasua (coatis) [22] and the detection of Leishmania in sand flies [19], suggesting the establishment of an epidemiological cycle in the area.
Capture, processing, and collection of biological samples
Bats were captured using mist nets (12 m × 3 m) placed in an area accessible to park visitors. The sample collection site included a children’s recreational area (Fig 1) surrounded by Semideciduous Seasonal Forest vegetation (19°57’03.3“S 43°54’30.4”W). Six sampling events were conducted in May, June (2), July (2), and December 2022, using 10 mist nets deployed from 6:00 PM to midnight, with inspections every 30 minutes.
Bat collection site within Mangabeiras Municipal Park (MMP), located in Belo Horizonte, Minas Gerais, Brazil.Images under a CC BY license, with permission from Débora Cristina Capucci, copyright 2024.
After capture, the bats underwent a clinical examination, were classified according to age and sex, and identified using the classification keys of Vizotto and Taddei [23] and Gardner [24]. Captured animals classified as threatened according to the IUCN [25], as well as pregnant or lactating females, those with offspring, or those carrying identification bands, were immediately released.
Bats used in the study were anesthetized for blood collection via intracardiac puncture and subsequently euthanized for the collection of spleen, heart, liver, ear skin, and wing membrane samples. The collected material was stored in tubes containing 1X PBS supplemented with antibiotics (Penicillin 5,000 U/mL and Streptomycin 5,000 μg/mL) and antifungal agents (5-Fluorocytosine) for trypanosomatid isolation in culture medium, and in tubes containing RNAlater for molecular analysis.
Culture medium
Biological samples of blood, tissues, and organs were macerated in sterile Petri plates using a scalpel blade and maintained at 25°C (±1°C) in biphasic NNN/LIT medium supplemented with 20% heat-inactivated sterile fetal bovine serum, antibiotics, and antifungal agents (as described above). Cultures were inspected weekly for two months for the observation of flagellated forms. Cultures showing fungal or bacterial contamination were discarded after macroscopic and/or microscopic evaluation using slide and coverslip examination.
Molecular detection of Trypanosomatidae
DNA was extracted using the Gentra Puregene Tissue Kit® (Qiagen) for tissue samples, while the Blood GenomicPrep Mini Spin Kit (GE Healthcare) was used for blood samples. To assess the quality and integrity of the extracted genomic material, DNA samples were subjected to PCR targeting the cytochrome B (cytB) gene for blood samples, using the primers cytb1 (5’-CCATCCAACATCTCAGCATGATGAAA-3’) and cytb2 (5’-GCCCCTCAGAATGATATTTGTCCTCA-3’) [26], and gamma-actin for tissues and organs, using the primers γ-fw (5’-ACAGAGAGAAGATGACGCAGATAATG-3’) and γ-rv (5’-GCCTGAATGGCCACGTACA-3’) [27]. For both reactions, 5 μL (10 ng/μL) of capybara (Hydrochoerus hydrochaeris) DNA, previously extracted by the research group, was used as a positive control.
The Nested-PCR technique, targeting the V7V8 region of the 18S gene (SSU rRNA), was used for the molecular detection of Trypanosomatidae. Reactions were performed using the primers TRY927F (5’-GAAACAAGAAACACGGGAG-3’) and TRY927R (5’-CTACTGGGCAGCTTGGA-3’), amplifying 927 bp in the first reaction, and the primers SSU561F (5’-TGGGATAACAAAGGAGCA-3’) and SSU561R (5’-CTGAGACTGTAACCTCAAAGC-3’), flanking 561 bp in the second reaction [28,29].
In all reactions, positive controls consisted of reference strains of Leishmania amazonensis (IFLA/BR/67/PH8), Leishmania braziliensis (MHOM/BR/75/M2903), Leishmania infantum (MHOM/BR/74/PP75), and Leishmania guyanensis (MHOM/BR/75/M4147). A negative control was also included, consisting of the PCR reagent mix without the addition of DNA template.
The amplified products were purified using the QIAquick PCR Purification Kit (Qiagen), and the amplicons were used for DNA sequencing under previously described conditions [30]. The obtained sequences were analyzed using FinchTV (Geospiza, Inc.) and MEGA X [31], and were compared with sequences deposited in GenBank using the BLAST tool. All sequences obtained were deposited in GenBank under the accession numbers OR656589 - OR656599.
Ethical statements
This study was approved by the Animal Ethics Committee (CEUA) of the Oswaldo Cruz Foundation (protocol LW-3/23) and the Pontifical Catholic University of Minas Gerais (protocol 14/2022). Licenses for the collection of bats were granted by SISBIO (numbers 80543-4 and 82980-1). Specimens were deposited at the Natural Science Museum of the Pontifical Catholic University of Minas Gerais (MCN/CA-09/2022).
Results
A total of 56 bats from the Phyllostomidae family were captured, representing seven species. The Stenodermatinae subfamily was the most represented, with 46 individuals (82.1%), followed by Carolliinae (6 individuals, 10.7%) and Glossophaginae (4 individuals, 7.1%). Artibeus lituratus was the most abundant species, with 28 individuals (50% of the total), followed by Sturnira lilium (N = 12, 21.4%) and Carollia perspicillata (N = 6, 10.7%) (Table 1).
Table 1: Species composition and distribution of bats captured by sampling month in Mangabeiras Municipal Park, 2022, Belo Horizonte, Minas Gerais, Brazil.
A total of 149 samples, including blood (N = 32), tissues (ear skin and wing membrane, N = 18 each), and organs (liver, N = 40; heart, N = 41) from 52 individuals, were analyzed in NNN/LIT culture medium. Fungal or bacterial contamination was observed in 48 samples (32.2%), while the remaining 104 samples (67.8%) showed no trypanosomatid growth after two months of incubation.
The detection of trypanosomatids through the V7V8 Nested-PCR revealed 11 positive samples (5.9%) from nine animals out of a total of 186 samples (Fig 2A). The findings include: Artibeus lituratus presenting DNA of Leishmania infantum (N = 3) and Trypanosoma sp. Neobat 3 (N = 1); Anoura caudifer carrying DNA of Trypanosoma sp. Neobat 4 (N = 1); Carollia perspicillata and Glossophaga soricina presenting DNA of Leishmania infantum (N = 1 each); Sturnira lilium presenting DNA of Trypanosoma sp. Neobat 3 (N = 1); and Platyrrhinus lineatus (N = 1) showing a mixed infection with DNA of Leishmania infantum and Leishmania braziliensis (Table 2).
Detection and validation of trypanosomatid DNA in bat samples.(A) Representative agarose gel electrophoresis (2%) of V7V8 Nested PCR products showing samples positive for trypanosomatid DNA. MM: Molecular weight marker (100 bp ladder); Lanes 1-6: Positive bat samples; Lane 7: Negative control; Lane 8: Positive control (L. braziliensis reference strain). (B) Verification of genomic DNA integrity through the amplification of endogenous control genes. The PCR products of gamma-actin (70 bp) gene were used as endogenous controls to confirm the quality and integrity of the extracted genomic material. MM: Molecular weight marker (100 bp ladder); Lanes 1-5: Gamma-actin gene amplification products from tissue and organ samples; Lane 6: Negative control; Lane 7: Positive control (capybara DNA, Hydrochoerus hydrochaeris).
Table 2: Detection of trypanosomatids in bat species from Mangabeiras Municipal Park, Belo Horizonte, Minas Gerais, Brazil, through V7V8 Nested-PCR.
Of the PCR-positive samples, seven (63.6%) yielded negative results in culture medium analysis, one (9.1%) showed contamination with fungi or bacteria, hindering evaluation for two months, and three (27.3%) were not collected. The endogenous amplification of cytb and gamma-actin genes demonstrated the integrity of the extracted genomic material (Fig 2B).
The phylogenetic analysis of the sequences confirmed the taxonomic positions of the isolates, placing them within the Trypanosoma wauwau clade (Fig 3A) and the Leishmaniinae subfamily (Fig 3B).
Maximum likelihood phylogenetic trees of Trypanosoma and Leishmania isolates obtained from bats sampled in Mangabeiras Municipal Park, Brazil, based on 18S SSU rDNA sequences.(A) Phylogenetic tree of Trypanosoma sp. Neobat sequences generated in this study, constructed using the Kimura 2-parameter (K2) model with 1,000 bootstrap replicates. (B) Phylogenetic tree showing the taxonomic positions of Leishmania infantum and Leishmania braziliensis sequences obtained in this study, constructed using the Kimura 2-parameter (K2) model with 1,000 bootstrap replicates. Bootstrap values (≥50%) are displayed at the nodes. Isolates from this study are highlighted in red circles, and reference sequences were retrieved from GenBank. Scale bars represent the number of substitutions per site.
Discussion
In this study, the diversity of bat trypanosomes in MMP, one of the largest urban parks in Latin America, located in Belo Horizonte, Minas Gerais, Brazil, was highlighted through standard capture efforts. Molecular data revealed a predominance of Leishmania infantum DNA (50%), the etiological agent of VL. Additionally, sequences from two divergent kinetoplastid taxa were identified: one similar to Trypanosoma sp. Neobat 3 (25%) and another to Trypanosoma sp. Neobat 4 (12.5%), both considered hidden bat parasites. Leishmania braziliensis sequences were also detected in bats (12.5%). This study expands knowledge regarding the occurrence of trypanosomatids circulating in urban bats.
Leishmania infantum was previously detected in Evandromyia edwardsi from MMP [19], and the proximity of this urban park to residential neighborhoods where dogs are present underscores an epidemiological scenario involving this parasite in the urban area of Belo Horizonte. In this study, four bat species (Artibeus lituratus, Carollia perspicillata, Glossophaga soricina, and Platyrrhinus lineatus) were found carrying L. infantum DNA in different biological samples, such as heart, liver, and spleen. All these bats have been previously associated with L. infantum in Brazil [32], reinforcing their role in the transmission cycle.
The MOTUs designated as Trypanosoma spp. Neobat comprise a complex of unnamed trypanosomes divided into T. sp. Neobat 1-6 [33–36]. Each MOTU likely represents a valid species requiring formal taxonomic description. However, as these flagellates appear to be incapable of growing in conventional culture media for trypanosomatids [37], taxonomic classification remains incomplete. Therefore, combined strategies such as parasitological and molecular assays are essential for determining bat positivity.
To date, T. spp. Neobat gene sequences have been detected in phyllostomid bats from Panama, Colombia, and Brazil. The finding of T. sp. Neobat 3 in samples from A. lituratus in Belo Horizonte confirms previous reports from the state of Rio de Janeiro, where the parasite was also detected in this bat [35], reinforcing the putative role of the genus Artibeus as a reservoir of this trypanosomatid [33–35]. Additionally, T. sp. Neobat 3 was identified for the first time in S. lilium, a bat known to be a putative reservoir of T. dionisii [35,38], expanding knowledge on putative hosts.
Trypanosoma sp. Neobat 4, a member of the T. wauwau complex, was long considered a specialized parasite of the Carollia genus, particularly C. perspicillata, where the parasite was originally described [35]. However, T. sp. Neobat 4 has been identified in bats belonging to other genera, such as Anoura caudifer, Glossophaga soricina, and Platyrrhinus recifinus, disproving the previous hypothesis of host specificity to Carollia [38]. In the present study, sequences 100% similar to T. sp. Neobat 4 were confirmed in a blood sample from A. caudifer, indicating that other bats may, in fact, play a role in the transmission cycle of this parasite.
Leishmania braziliensis was previously found in Evandromyia edwardsi collected in caves from MMP, highlighting a putative transmission cycle involving this sand fly and other mammals [19]. In this study, P. lineatus was found to be positive for L. braziliensis in a blood sample, reinforcing previous findings [39]. Several Brazilian bats have been reported carrying L. braziliensis, especially insectivorous species such as Cynomops planirostris [40], Eumops glaucinus [41], Molossus molossus [42], M. rufus [41], and Lasiurus cinereus [40]. Although sand fly ingestion by bats is uncertain, and the probability of accidental transmission by grooming cannot be ruled out, the presence of Leishmania in frugivorous species [39,41], nectarivorous species [42,43], and hematophagous species [43] suggests alternative transmission routes, in addition to the traditional sand fly bite.
Knowledge gaps remain regarding the biological and ecological properties of MOTUs T. spp. Neobats, such as their vectors, transmission cycles, and interactions with hosts. Although sand flies are not considered natural vectors of Trypanosoma, naturally infected females with parasites of this genus have been reported in Brazil. Among several sand fly species recorded in the study area [19], Nyssomyia whitmani has been associated with Trypanosoma sp. infection in Bahia [44]. Furthermore, representative species of the Shannoni series of the Psathyromyia genus have been found naturally infected with Trypanosoma rangeli and other unidentified trypanosomatids in Rondônia and Acre states, respectively [45]. However, a previous study on the sand fly fauna in the study area did not detect Trypanosoma infections in these species. While sand flies may play a role in the transmission of certain Trypanosoma species, more data are needed to establish their role in the transmission of T. spp. Neobats in urban environments.
In addition to sand flies, triatomine bugs, which are vectors of Trypanosoma cruzi, may also be involved in the transmission cycles of other trypanosomatids in bats. Indeed, the only known vector of T. cruzi marinkellei consists of triatomines from the genus Cavernicola, which are associated with bat colonies in caves, hollow trees, and palm trees [46,47]. Moreover, although the vector of T. cruzi Tcbat remains unknown, this genotype is most closely related to T. cruzi TcI, a group that is preferentially associated with didelphids and Rhodnius spp. in arboreal niches [48]. Similarly, the T. rangeli lineage TrE has only been found in bats and in the triatomine Rhodnius pictipes from Central and Amazonian regions of Brazil [49].
The mechanisms by which bats acquire these parasites remain poorly understood. Bats may acquire T. cruzi either through blood meals from infected triatomines or by ingesting these arthropods. Triatomines and bats may interact during nocturnal foraging, when both are active, or during the day, when bats roost in trees or caves, potentially allowing triatomines to feed on them [50]. This underscores the potential involvement of multiple vectors, such as sand flies and triatomines, in the zoonotic transmission cycles of Trypanosoma species, particularly in urban areas where bats, sand flies, and triatomines may ecologically overlap. Understanding these interactions is essential for elucidating the dynamics of zoonotic transmission and the role of bats as reservoirs for trypanosomatid parasites.
Interestingly, a mixed infection was detected in P. lineatus, with L. infantum detected in the heart and L. braziliensis in the blood. Although L. braziliensis seems to exhibit tropism for dermal tissues, the visceralization of Viannia species is considered common and transient, at least in animal models [51–55]. The occurrence of mixed natural infections with two or more Leishmania species is apparently rare but has been reported in bats [56] and rodents [57]. Moreover, mixed infections with Trypanosoma spp. have also been described [58,59], highlighting that the gregarious habits of bats may facilitate the transmission of these parasites.
Despite the limitations of this study, such as sampling in a single location within MMP and the parasitological monitoring of tissue and organ samples presenting a high level of fungal and bacterial contamination, this study demonstrates the occurrence of Trypanosomatidae species carried by bats in urban areas of Belo Horizonte. The cycles of L. braziliensis and L. infantum appear to be established in MMP, with the confirmation of sand flies and bats carrying these parasites. Further studies are needed to ascertain the role of A. caudifer, A. lituratus, and S. lilium in the transmission cycle of T. spp. Neobat and the involvement of putative vectors, such as kissing bugs.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kaufer A, Ellis J, Stark D, Barratt J. The evolution of trypanosomatid taxonomy. Parasit Vectors. 2017;10(1):1–7. doi: 10.1186/s 13071-017-2204-7 28595622 PMC 5463341 · doi ↗ · pubmed ↗
- 2Bruschi F, Gradoni L. The leishmaniases: old neglected tropical diseases. Springer; 2018.
- 3Shikanai Yasuda MA. Emerging and reemerging forms of Trypanosoma cruzi transmission. Mem Inst Oswaldo Cruz. 2022;117:e 210033. doi: 10.1590/0074-02760210033 35584508 PMC 9113729 · doi ↗ · pubmed ↗
- 4Kostygov AY, Albanaz ATS, Butenko A, Gerasimov ES, Lukeš J, Yurchenko V. Phylogenetic framework to explore trait evolution in Trypanosomatidae. Trends Parasitol. 2024.10.1016/j.pt.2023.11.00938065790 · doi ↗ · pubmed ↗
- 5Schneider A, Ochsenreiter T. Failure is not an option - mitochondrial genome segregation in trypanosomes. J Cell Sci. 2018;131(18):jcs 221820. doi: 10.1242/jcs.221820 30224426 · doi ↗ · pubmed ↗
- 6Brenière SF, Waleckx E, BarnabéC. Over Six Thousand Trypanosoma cruzi Strains Classified into Discrete Typing Units (DT Us): Attempt at an Inventory. P Lo S Negl Trop Dis. 2016;10(8):e 0004792. doi: 10.1371/journal.pntd.0004792 27571035 PMC 5003387 · doi ↗ · pubmed ↗
- 7Jansen A, Xavier S das C, Roque A. Trypanosoma cruzi transmission in the wild and its most important reservoir hosts in Brazil. Parasites & Vectors. 2018;11(1):1–25.30189896 10.1186/s 13071-018-3067-2PMC 6127949 · doi ↗ · pubmed ↗
- 8de Araújo GR, de MelloÉM, de Oliveira VNGM, Dos Santos TR, Nunes RV, de Andrade HM, et al. Bats as hosts of Leishmania (Leishmania) infantum in Minas Gerais, an endemic area for visceral leishmaniasis. Vet Parasitol Reg Stud Reports. 2022;32:100740. doi: 10.1016/j.vprsr.2022.100740 35725103 · doi ↗ · pubmed ↗