Selective benefit of the sucrose TonB-dependent receptor, SucA, in Caulobacter crescentus
Erin NewRingeisen, Jacy Jordahl, Lisa Bowers

TL;DR
This paper investigates the function of the SucA protein in Caulobacter crescentus, showing when it provides a growth advantage and how it works.
Contribution
The study identifies specific conditions where the SucA TBDR provides a measurable fitness benefit and confirms its reliance on the proton motive force.
Findings
SucA provides a significant growth advantage under specific environmental conditions.
SucA activity depends on the proton motive force, consistent with canonical TBDR function.
Abstract
Gram-negative bacteria have outer membrane proteins called TonB-dependent receptors (TBDRs) that facilitate energy-dependent transport of substrates. Caulobacter crescentus is a gram-negative bacterium with a large set of TBDRs, yet the function of many of these TBDRs remains uncharacterized. This study focuses on SucA, a TBDR that transports sucrose. Previous studies showed that sucA expression was induced in the presence of sucrose, yet did not provide a measurable fitness advantage under the conditions tested. This work identifies conditions where sucA does confer a significant growth advantage and provides evidence that SucA activity relies on the proton motive force, a feature of canonical TBDRs.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
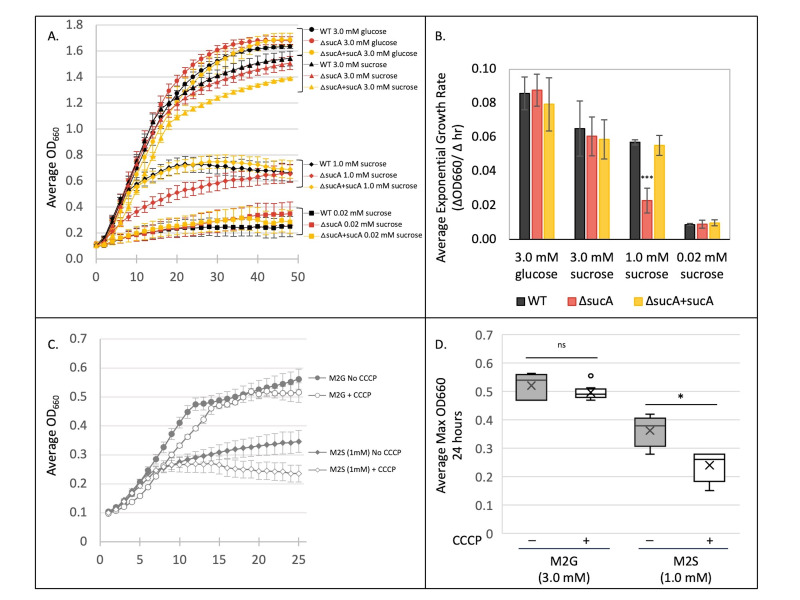 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTracheal and airway disorders · Neonatal Respiratory Health Research · Congenital heart defects research
Description
Caulobacter crescentus is a free-living, oligotrophic, gram-negative bacterium commonly found in soil and aquatic environments (Curtis and Brun, 2010; Wilhelm, 2018) . Like most free-living oligotrophic bacteria, it likely encounters a variety of nutrients that are scarce or fleeting in its natural environment (Hentchel et al., 2019) . To respond to changing nutrient availability, gram-negative bacteria have evolved different mechanisms for nutrient uptake across the outer membrane (OM). Outer membrane porins (OMPs) allow the passive diffusion of solutes with molecular mass < ~600 Da but for substrates that are too large or too scarce to diffuse through OMPs, Caulobacter relies on TonB-dependent receptors (TBDRs) for their active uptake (Noinaj et al., 2010) .
TBDRs are located in the OM of gram-negative bacteria and bind substrates with high affinity. Substrate binding causes a conformational change, allowing the TBDR to interact with TonB, a protein that bridges the periplasmic space (Schauer et al., 2008). TonB physically couples the TBDR with a cytoplasmic membrane (CM) motor protein complex (ExbB-ExbD), which is powered by the proton motive force (pmf) (Wiener, 2005; Noinaj et al., 2010; Silale and Van Den Berg, 2023) . This interaction uses energy derived from the pmf to drive transport of the substrate across the OM. Different TBDRs have been shown to transport a wide variety of substrates including vitamin B 12 complex, iron chelators, transition metals, carbohydrates, and aromatic compounds (Menikpurage et al., 2019; Mazzon et al., 2014; Neugebauer et al., 2005; Blanvillain et al., 2007; Eisenbeis et al., 2008; Lohmiller et al., 2008; Modrak et al., 2018; Balhesteros et al., 2017; Presley et al., 2014; Fujita et al., 2019). While most gram-negative bacteria encode a few TBDR genes (Blanvillain et al., 2007) , Caulobacter crescentus is predicted to encode 67 different TBDRs (Nierman et al., 2001) , most of which are yet to be characterized. Furthermore, many of these TBDRs are conserved throughout the α-proteobacteria, which suggests they may confer a selective benefit in some environments, yet they are often dispensable for growth under typical laboratory growth conditions (Blanvillain et al., 2007; Christen et al., 2011; Déjean et al., 2013) .
SucA provides a selective benefit to cells growing with sucrose as the sole carbon source.
SucA is a Caulobacter TBDR that transports sucrose. Previous studies determined its expression is strongly induced in the presence of sucrose, yet it does not confer a growth advantage for cells under typical laboratory growth conditions when sucrose is supplied at 3.0 mM in the growth media (Modrak et al., 2018) . We wondered if SucA might confer a growth advantage at a lower concentration of sucrose, since natural aquatic environments have been reported to have sucrose concentrations in the µM range (Sogin et al., 2022) . To test this, we compared the growth rate of three isogenic strains: WT Caulobacter (NA1000), an in-frame unmarked sucA deletion strain ( ΔsucA ), and the complemented deletion strain with sucA expressed from its native promoter on a low-copy plasmid ( * ΔsucA+pSucA * ) . The growth of these three strains was measured over 48 hours in minimal media containing either glucose (3.0 mM) or sucrose (0.02 mM, 1.0 mM or 3.0 mM) as the sole carbon source ( Figure 1A ). Maximum cell density depends on carbon content and may not reveal subtleties in cell division rates; therefore, we analyzed fitness of the three strains by examining the exponential growth rate ( Figure 1B ). As expected, all three strains had equal exponential growth rates when glucose (3.0 mM) was the sole carbon source and at the highest concentration of sucrose (3.0 mM) where sucrose would be expected to enter the cells via facilitated diffusion. At the lowest concentration of sucrose tested (0.02 mM), there was not significant growth for any of the three strains in 48 hours (with this low amount of carbon, cells could at most perform one doubling). However, at the intermediate sucrose concentration (1.0 mM), the cells expressing sucA (WT and ΔsucA
- sucA) had a significantly greater exponential growth rate compared to cells lacking sucA ( ΔsucA ) ( Figure 1A and 1B). Typical TBDRs are known to have a high affinity for their substrate, often with a K d in sub-µM range (Neugebauer et al., 2005; Blanvillain et al., 2007) so it was surprising to observe a significant difference in exponential growth rate between 3.0 mM sucrose and 1.0 mM sucrose. However, after 48 hours, the cells lacking sucA ( ΔsucA ) did eventually reach the same maximum optical density as the cells expressing *sucA * ( Figure 1A ). Thus, there may be other slower uptake mechanisms for sucrose.
SucA activity relies on the proton motive force.
A canonical feature of TBDRs is their reliance on the pmf generated across the CM to drive the transport of nutrients across the OM (Bradbeer and Woodrow, 1976) . The proton gradient can be disrupted by the non-specific proton ionophore carbonyl cyanide-m-chlorophenylhydrazone (CCCP). In order to test whether SucA-mediated transport of sucrose across the OM is dependent upon the pmf of an intact CM, growth of WT Caulobacter cells was monitored in the presence and absence of a non-lethal concentration of CCCP. If SucA activity depends on the pmf, its activity should be reduced in the presence of CCCP and this effect would be most obvious at lower concentrations of the substrate. WT Caulobacter cultures (NA1000) containing either 3.0 mM glucose (M2G) or 1.0 mM sucrose as the sole carbon source were incubated with or without 6.25 µM CCCP and the culture density was measured for 24 hours ( Figure 1C ). For cells grown in M2G, there was no significant change in optical density after 24 hours with and without CCCP ( Figure 1C and 1D); thus, we determined that cell viability and glucose uptake was not affected at this concentration of CCCP. However, the growth of cells in M2S with 1.0 mM sucrose was significantly inhibited by the addition of CCCP ( Figure 1C and 1D). The inhibitory effect of CCCP was first observed in the M2S cultures after around 10 hours of incubation. In the M2S cultures, the cells treated with CCCP transitioned to stationary phase earlier than the cells without CCCP, and reached a lower optical density after 24 hours. Thus, CCCP diminishes the selective advantage of SucA, evidence that SucA-mediated transport depends on the energy from the pmf, typical of a canonical TBDR.
In other environmental bacteria, some TBDRs that transport carbohydrates are dispensable for growth in nutrient media but essential for growth and virulence on plant leaves (Blanvillain et al., 2007; Déjean et al., 2013) . Similarly, in studies shown here, the Caulobacter crescenuts sucA gene is dispensable for growth at a relatively high sucrose concentration (3.0 mM) but confers a significant growth advantage at a reduced sucrose concentration (1.0 mM). Furthermore, this growth advantage provided by the sucA gene depends on an intact pmf. Together, these findings contribute to a more nuanced understanding of the role and selective advantage of TBDRs and highlight the importance of laboratory growth conditions when studying the function and selective benefit of nutrient uptake systems.
Methods
Bacterial strains and growth conditions
The *Caulobacter crescentus * strains in this study were NA1000 (referred to here as WT), LB235 (Modrak et al., 2018) (a non-polar sucA deletion strain which is referred to here as Δ sucA ), and LB236 (Modrak et al., 2018) (the sucA deletion strain complemented with sucA on a low copy plasmid under its own promoter, referred to here as Δ *sucA *
- p SucA ). All strains were incubated at 30°C in M2 minimal medium (Ely, 1991) , supplemented with either 3.0 mM D-glucose (M2G), 3.0 mM sucrose (M2S), 1.0 mM sucrose, or 0.02 mM sucrose ** . ** Kanamycin was added to all cultures of LB236 (Δ *sucA *
- p *SucA) * at 25 μg/ml.
**Growth Assay: ** NA1000 , Δ sucA , and Δ *sucA *
- p SucA were grown in M2G overnight and then spun down and the spent media was removed. Cells were then washed in 1 mL of M2 minimal medium supplemented with either 3.0 mM D-glucose, 3.0 mM sucrose, 1.0 mM sucrose or 0.02 mM sucrose and then resuspended in 1 mL of the same media to a starting OD 660 = 0.05.
For the samples treated with carbonyl cyanide-m-chlorophenylhydrazone, CCCP was added with a final concentration of 6.25 µM after cells were washed and resuspended. Five technical replicates for each sample were loaded into a 96-well plate with 200 μL of culture per well. The plates were incubated at 30°C with aeration and the optical density (OD 660 ) was recorded every hour with a BioTek Epoch-2 microplate reader running Gen5 TS 2.09 software. The first derivative of the growth curve (change in OD 660 vs time) was used to identify the period of exponential growth and exponential growth rate was calculated for each sample (ΔOD/ΔT). Growth curves were conducted in three independent biological experiments, each with five technical replicates. (*p-value <0.05)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Balhesteros Heloise Shipelskiy Yan Long Noah J. Majumdar Aritri Katz Benjamin B. Santos Naara M. Leaden Laura Newton Salete M. Marques Marilis V. Klebba Phillip E. 2017315 Ton B-Dependent Heme/Hemoglobin Utilization by Caulobacter crescentus Hut A Journal of Bacteriology 19960021-919310.1128/jb.00723-16PMC 533166628031282 · doi ↗ · pubmed ↗
- 2Blanvillain Servane Meyer Damien Boulanger Alice Lautier Martine Guynet Catherine DenancéNicolas Vasse Jacques Lauber Emmanuelle Arlat Matthieu 2007221 Plant Carbohydrate Scavenging through Ton B-Dependent Receptors: A Feature Shared by Phytopathogenic and Aquatic Bacteria P Lo S ONE 221932-6203 e 224e 22410.1371/journal.pone.000022417311090 PMC 1790865 · doi ↗ · pubmed ↗
- 3Bradbeer C Woodrow M L 1976101 Transport of vitamin B 12 in Escherichia coli: energy dependence Journal of Bacteriology 12810021-91939910410.1128/jb.128.1.99-104.1976135757 PMC 232830 · doi ↗ · pubmed ↗
- 4Christen B Abeliuk E Collier JM Kalogeraki VS Passarelli B Coller JA Fero MJ Mc Adams HH Shapiro L 2011830 The essential genome of a bacterium.Mol Syst Biol 752852810.1038/msb.2011.5821878915 PMC 3202797 · doi ↗ · pubmed ↗
- 5Curtis Patrick D. Brun Yves V. 201031 Getting in the Loop: Regulation of Development in Caulobacter crescentus Microbiology and Molecular Biology Reviews 7411092-2172134110.1128/mmbr.00040-0920197497 PMC 2832345 · doi ↗ · pubmed ↗
- 6Déjean Guillaume Blanvillain‐BaufuméServane Boulanger Alice Darrasse Armelle de Bernonville Thomas Dugé Girard Anne‐Laure Carrére Sébastien Jamet Stevie Zischek Claudine Lautier Martine SoléMagali Büttner Daniela Jacques Marie‐Agnès Lauber Emmanuelle Arlat Matthieu 2013227 The xylan utilization system of the plant pathogen Xanthomonas campestris pv campestris controls epiphytic life and reveals common features with oligotrophic bacteria and animal gut symbionts New Phytologist 19830028-646X 89991510.1111/nph · doi ↗ · pubmed ↗
- 7Eisenbeis Simone Lohmiller Stefanie Valdebenito Marianne Leicht Stefan Braun Volkmar 200881 Nag A-Dependent Uptake of N -Acetyl-Glucosamine and N -Acetyl-Chitin Oligosaccharides across the Outer Membrane of Caulobacter crescentus Journal of Bacteriology 190150021-91935230523810.1128/jb.00194-0818539735 PMC 2493260 · doi ↗ · pubmed ↗
- 8Ely Bert 1991[17] Genetics of Caulobacter crescentus Methods in Enzymology 0076-687937238410.1016/0076-6879(91)04019-k 1658564 · doi ↗ · pubmed ↗