The rediscovery of Ohwialuteola (Fabaceae, Papilionoideae) after 50 years and comparative analysis of Ohwia species in plastid genome sequence

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
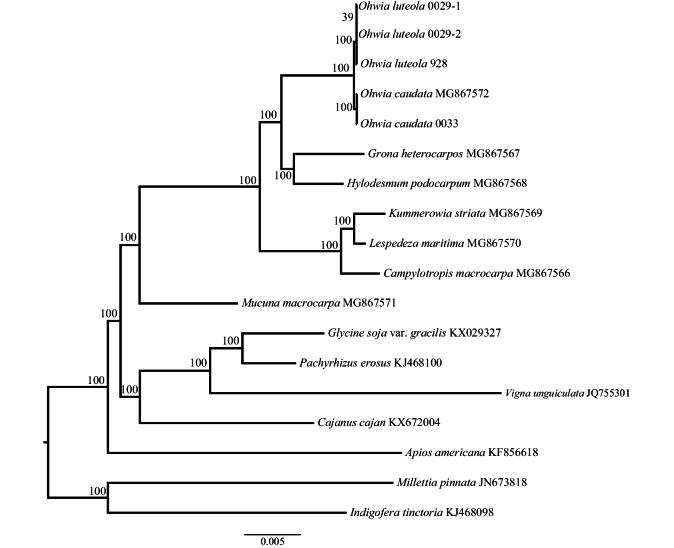 Figure 1
Figure 1 Figure 2
Figure 2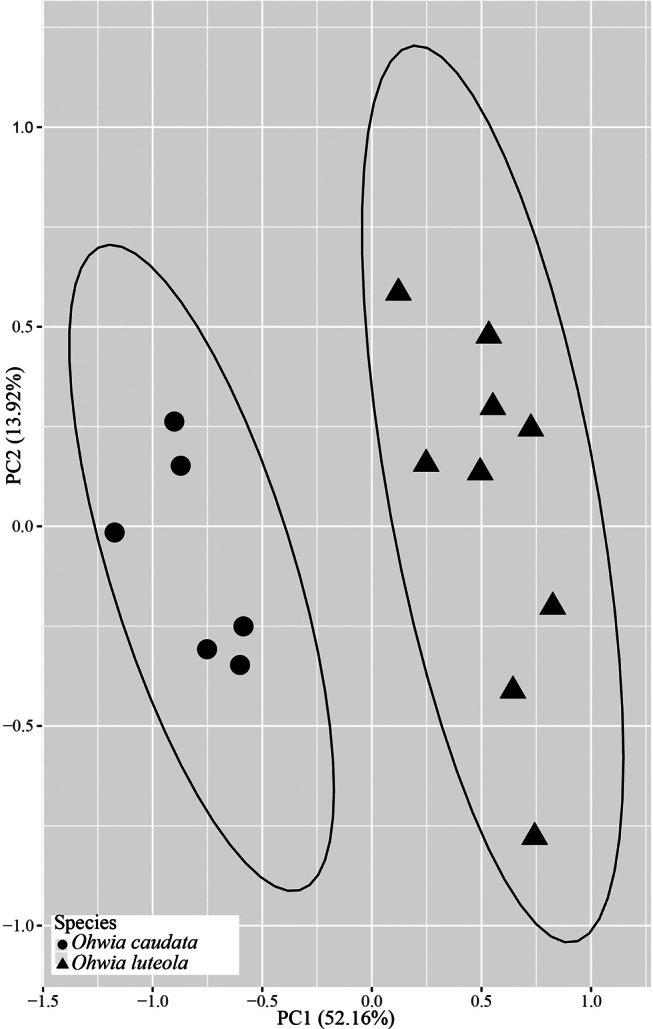 Figure 3
Figure 3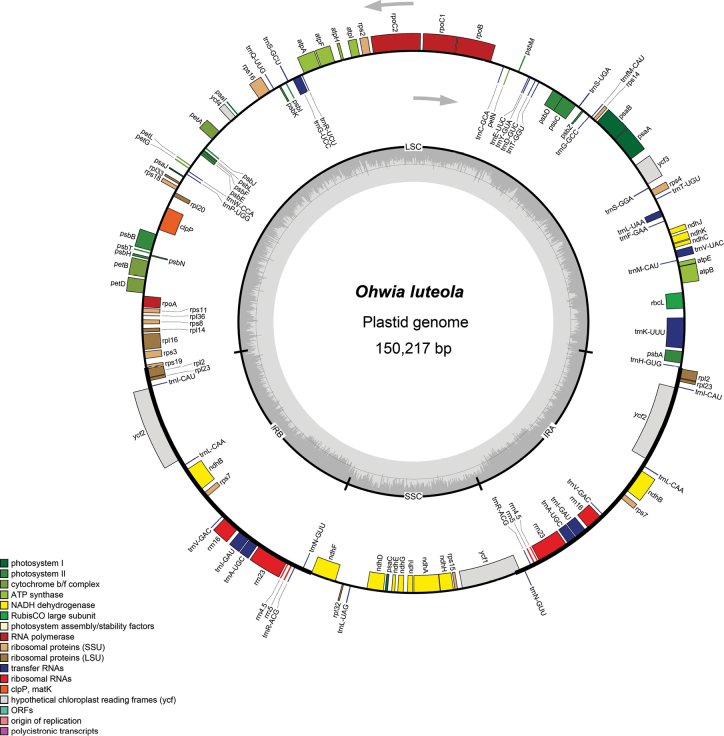 Figure 4
Figure 4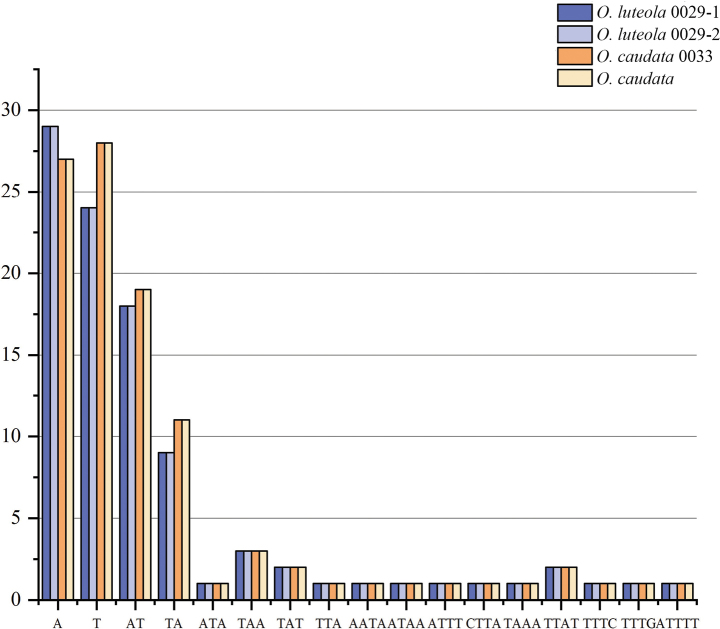 Figure 5
Figure 5 Figure 6
Figure 6| Species | Total size (bp) | Length of | Length of | Length of | GC content (%) | No. of genes |
|---|---|---|---|---|---|---|
| 150,217 | 83,227 | 18,442 | 24,274 | 35.1% | 128 | |
| 150,217 | 83,227 | 18,442 | 24,274 | 35.1% | 128 | |
| 150,250 | 83,242 | 18,480 | 24,264 | 35.1% | 128 | |
|
| 150,249 | 83,241 | 18,480 | 24,264 | 35.1% | 128 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant and Fungal Species Descriptions · Genomics and Phylogenetic Studies · Botanical Research and Chemistry
Introduction
Fabaceae (or Leguminosae), the third largest family of angiosperm, comprises more than 19,500 species in ca. 765 genera, 36 tribes, and 6 currently recognized subfamilies (Caesalpinioideae, Cercidoideae, Detarioideae, Dialioideae, Duparquetioideae, and Papilionoideae) (Azani et al. 2017). The legume plants have highly diversified in growth forms including trees, shrubs or herbs, sometimes climbing or decumbent, and ca. 88% of legume species have the ability to establish associations with nitrogen-fixing bacteria (Sprent et al. 2017; Zhang et al. 2020). Many legume species are economically and ecologically important (Yahara et al. 2013).
Ohwia H. Ohashi, is a small genus within the tribe Desmodieae of subfamily Papilionoideae containing two species, i.e., O.luteola (H.Ohashi & T.Nemoto) H. Ohashi and O.caudata (Thunb.) Ohashi (Huang and Ohashi 2010). Members of this genus are characterized by their shrub or subshrub growth habit, featuring pinnately trifoliolate leaves, persistent stipules, and winged petioles. Ohwialuteola is endemic to Yunnan Province (China) and characterized by corolla pale yellow, while O.caudata widely distributed in East Asia, and corolla greenish or yellowish white (Huang and Ohashi 2010). Ohwialuteola was described in 1998 based on a single number collection from northeastern Yunnan Province (China) in 1972 (Ohashi and Nemoto 1998), but additional specimens of O.luteola have not been recorded for more than 50 years.
In a recent exploration of Zhangjiajie city (Hunan Province, China), we collected an unknown Ohwia species with similar morphological characteristics to O.caudata. However, they have smaller leaflets with obtuse apex, which are obviously different from O.caudata. After having a determination of the material by Hiroyoshi Ohashi, one of the original authors of Ohwialuteola as Desmodiumluteolum H.Ohashi & T.Nemoto, we made a morphological comparison of our material with the images of the type of O.luteola (KUN) and habitat description and confirmed that they belong to O.luteola. Therefore, the purpose of our research described here was to provide an insight into the taxonomic status of the O.luteola by comparing morphological features and analyzing the plastome.
Material and methods
Morphology observation and measurement
In total, 9 individuals of the O.luteola and 6 individuals of O.caudata were examined and herbarium voucher specimens deposited in the herbariums of the Department of Biology, Jishou University (JIU) and the Kunming Institute of Botany (KUN). Fourteen morphological characters were selected for the morphometric analysis. The characters include terminal leaflet length; terminal leaflet width; lateral leaflet length; lateral leaflet width; petiole width; terminal leaflet petiole length; lateral leaflet petiole length; number of inflorescences per branch; number of flower nodes per inflorescence; flower stipe length; wing base (1-slightly auriculate, 2-distinctly auriculate); terminal leaflet length/width ratio; lateral leaflet length/width ratio. We performed a principal component analysis (PCA) using R v.4.0.2 (R Core Team 2020) to project and visualize trends in morphological variability across our samples.
DNA extraction and sequencing
Total genomic DNA was extracted from silica gel-dried materials and herbarium material (three individuals of O.luteola and one individual of O.caudata) using the Plant Genomic DNA Kit (TianGen Biotech, Beijing, China) following the manufacturer’s protocol. DNA libraries were constructed with paired-end reads (PE150) were generated using an Illumina NovaSeq 6000 platform. Library construction and sequencing were carried out at Novogene Co., Ltd. in Beijing, China. Approximate 4 Gb of raw-reads were obtained for each sample.
Plastid genome assembly, annotation, and comparison
Plastomes were assembled using GetOrganelle (Jin et al. 2020) based on the clean reads. The plastome of O.caudata (MG867572) was selected as a reference (Jin et al. 2019). We detected the boundaries of large single-copy (LSC), small single-copy (SSC), and two inverted repeats (IRs) using RepeatFinder v.1.0.1 (Volfovsky et al. 2001). The final annotation was conducted in GENEIOUS v. 11.1.4 (Kearse et al. 2012). A circular plastome map was drawn in OGDraw v.1.3.1 (Greiner et al. 2019). SSRs are tandem repeats of one to six nucleotide long DNA motifs with high variability, multi-allelic nature, codominant inheritance, repeatability, relative abundance, and other traits that hold great promise in evolutionary and population genetics studies. The MISA program (http://pgrc.ipk-gatersleben.de/misa/) was used to identify the SSR, with a minimum number of repeat units of 10, 5, 4, 3, 3, and 3 for mono-, di-, tri-, tetra-, penta-, and hexa-nucleotides, respectively.
Phylogenetic analyses
We failed to obtain a complete plastid genome of the sample of ‘O.caudata 928’ (isotype) because the DNA of this sample was extracted from herbarium material collected over 50 years ago. To determine the phylogenetic position of O.caudata, a total of 34 plastid CDS were extracted using GENEIOUS v.11.1.4. The outgroups and other Leguminosae species were selected based on the work of Jin et al. (2019). Voucher information and GenBank accession numbers were provided in Appendix 1. Sequences were aligned with MAFFT (Katoh and Standley 2013). The concatenated plastid CDS dataset is deposited in DRYAD (https://doi.org/10.5061/dryad.4qrfj6qn5). Maximum likelihood (ML) analysis was performed using RAxML-HPC v.8.2.4 (Stamatakis 2014), with the GTR + I + G model and run for 1000 bootstrap iterations. The phylogenetic trees were visualized using FigTree v.1.4.2 (Rambaut 2014).
Results and discussion
The aligned plastid CDS matrix contained 34,582 sites. The ML tree is shown in Fig. 1. Our results showed that O.luteola from Hunan Province and isotype from Yunnan Province were clustered together and strongly supported O.luteola sister to O.caudata (BS = 100%, Fig. 1). This sister relationship is also supported by morphological characters. Morphological synapomorphies of O.luteola and O.caudata included pinnately 3-foliolate, stipules persistent, calyx campanulate and 4-lobed (Huang and Ohashi 2010).
Maximum likelihood (ML) analysis based on the 34 plastid protein-coding genes. ML bootstrap (BS) values are given above the branches.
Morphologically, most leaflets of O.caudata are lanceolate or oblong (Fig. 2D) (Ohashi 2005), terminal leaflets have a length/width ratio of up to 6.7, and leaflets apex acuminate. In contrast, O.luteola has oblong-elliptic leaflets, the terminal leaflet length/width ratio ranges from 2.9 to 3.6, and the leaflets apex is acute (Fig. 2H). The wings of O.luteola are distinctly auriculate at the base, and the inner side is indistinctly rugose (Fig. 2E) (vs. wings slightly auriculate at the base and inner side distinctly rugose). Also, it differs by its hilum at the center of the axis and not over 3/5 length of seeds (Fig. 2G) (vs. hilum off-center and over 1/2 length of seeds). More importantly, O.luteola grows on limestone along the river, and O.caudata usually grows under the forest. It is noteworthy that the corolla of O.luteola is described as pale yellow (Ohashi and Nemoto 1998), and the flowers observed from fresh materials collected in Hunan province are greenish-white to yellowish-white. Additionally, the flowers of the specimen turn yellow after drying. Morphological traits from 15 specimens were explored using PCA (Fig. 3). The first two principal components identified by PCA accounted for 66.08% of the variation across all characters. The PCA results showed that individuals of O.luteola and O.caudata formed distinct clusters.
Comparison of Ohwialuteola and O.caudataA–DO.caudata (A wings B ovary C seed D Branch) E–HO.luteola (E wings F ovary G seed H Branch).
Principal components plots based on fourteen morphological characters.
We sequenced, assembled, and annotated three plastomes representing O.luteola (two individuals) and O.caudata (one individuals). The features of these plastomes are summarized in Table 1. Plastome map for the O.luteola is shown in Fig. 4. Consistent with previous studies in legumes, the plastomes were highly conserved, with no structural variations or content rearrangements (Jin et al., 2019). The plastome sizes of the Ohwia species ranged from 150,217 bp for O.luteola to 150,250 bp for O.caudata. All the two species presented a classical quadripartite structure, a LSC, an SSC, and two IRs. The length of the LSC region ranged from 83,227 bp to 83,242 bp. The SSC region varied from 18,442 bp to 18,480 bp in length, and that of the IR regions ranged from 24,264 bp to 24,274 bp (Table 1). A total of 128 genes were identified, including 83 protein-coding genes, 37 transfer RNA (tRNA) genes, and 8 ribosomal RNA (rRNA) genes. The GC content of the two species was identical in the whole chloroplast genome (35.1%), with the GC content in the IR regions (42.0%) noticeably higher than that in the SSC (28.3%) and LSC (32.6%) regions in each chloroplast genome. Our study identified a total of 384 SSRs in the two Ohwia species (Fig. 5). The number of SSRs in Ohwiacaudata is 95, while the number of simple repeats in O.luteola is 97. Among them, the A/T mononucleotide SSRs are the most abundant.
Plastid genome map of Ohwialuteola.
Specific forms of SSRs in 4 genomes from Ohwia.
Taxonomy
Ohwia
luteola
Taxon classificationPlantaeFabalesFabaceae
(H. Ohashi & T. Nemoto) H. Ohashi
282DB6A8-11E1-5AFA-8C93-8F4A73F8472F
Diagnosis.
Ohwialuteola resembles O.caudata but differs from the latter by having terminal leaflets length/width ratio rang from 2.9 to 3.6 (vs. terminal leaflets length/width ratio rang from 4.2 to 6.7), leaflets apex acute (vs. acuminate) terminal inflorescences (vs. terminal and axillary), wings with distinctly auriculate at base, inner side indistinctly rugose (vs. wings with slightly auriculate at base, inner side distinctly rugose), hilum center, not over 3/5 length of seed (vs. hilum off-center, over 1/2 length of seed).
OhwialuteolaA habitat B, C habit D fruits E inflorescence F flower G calyx H standard I wings J keel-petal K ovary and stamens L seed.
New record.
Populations of Ohwialuteola are known from Xixiping Street, Yaping village, and Bamaoxi village of Zhangjiajie. It is growing on limestone along the Lishui River. The companion species mainly including Adina rubella Hance, Distyliumbuxifolium (Hance) Merr., and Cornusquinquenervis Franch.
Specimens examined.
China • Hunan: Zhangjiajie City, Yongding District, Sanjiaguan Township, Yaping village, under Zhanghua Lishui Large Bridge, on limestone areas along Lishui River, alt. 218 m, 29.111375°N, 110.258679°E, 31 Aug. 2023, M. H. Zhang et al. 0029 (JIU); • Yunnan: Jinping County, Laomeng River, alt. 750 m, 20 May 1974, Lüchun Exped. 944 (KUN 0608532); • Yiliang County, Niujie, alt. 450 m, 23 Sep. 1972, Northeast Yunnan Exped. 928 (KUN 0608538).
Revised description.
Shrubs, erect, 1–2 m tall, main stem ca. 1 cm in diam at base, much branched. Leaves 3-foliolate, thickly papery to subleathery, both surfaces pilose and more densely hairy on raised veins, margin entire. Petiole 2–3 cm long, with narrowly winged on both, 0.2–0.3 mm wide. Terminal leaflet oblong-elliptic, widest near the middle part, 4–7.1 × 1.5–2.4 cm, principal veins 10–14 pairs, reaching the leaf margin, apex acute, base cuneate, small petiole 0.8–1.2 cm long, pubescent. Lateral leaflets smaller, 3.7–6.4 × 1.1–1.8 cm, small stipe 0.2–0.3 cm long, widest near the middle part, principal veins 6–12 pairs, reaching the leaf margin, apex acute, base cuneate, small petioles 0.2–0.3 cm long, densely pubescent. Stipules 3–7 mm long, ca. 1.0 mm wide at the base, densely pubescent, persistent. Inflorescences terminal, 7–19 cm long, rachis densely pubescent intermixed with minute uncinate and appressed or spreading longer hairs, 2–4-flowered at each node; bracts subulate, ca. 0.3 cm long. Pedicels 0.4–0.6 cm long, densely pubescent. Calyx campanulate, 0.8–1.2 cm long, outside densely appressed pubescent, 4-lobed, lobes united for ca. 1/2 length, lobes ca. 0.5 cm long, longest one linear-lanceolate. Corolla greenish-white or yellowish-white, ca. 1.5 cm long, distinctly veined; standard elliptic, 0.8–1.7 × 0.5–1.0 cm, claw ca. 2.5 mm, slightly auriculate at base, apex slightly retuse; wings shorter than keel, 1.3–1.6 cm long, apex obtuse, lamina narrowly elliptic, distinctly auriculate at base, claw ca. 3 mm, keel 0.8–1.8 cm long, apex rounded, slightly auriculate at base, claw ca. 3 mm. Vexillary stamen slightly connate at base from other 9, ca. 1.6 cm long, puberulent at upper part; remaining 9 stamens connate for 4/5 or more of length, puberulent at upper part. Style curved upward, ovary densely ap-pressed pilose on both sutures. Disk present at base of pistil. Legume linear, flat, 3.5–7 cm long, stipe ca. 5 mm long, 3–6-jointed; articles nearly rectangle, 1–1.3 × 0.5–0.7 cm, with dense, transparent to brown, uncinate hairs. Seeds compressed, reniform, ca. 12 × 5 mm; hilum center, not over 3/5 length of seed. Flowering from July to early September; fruiting from September to November.
Conservation status.
During our field investigations in 2022 and 2024, many populations of O.luteola were found in Zhangjiajie. The number of individuals of each population ranges from tens to hundreds. In addition, it is distributed along the river. We believe that it should have a much wider distribution than is currently known. Due to its wide distribution range and large population size, O.luteola is here recommended as Least Concern (LC) (IUCN 2022).
Supplementary Material
XML Treatment for Ohwia luteola
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Azani N Babineau M Bailey CD Banks H Barbosa AR Pinto RB Boatwright JS Borges LM Brown GK Bruneau A Candido E Cardoso D Chung K-F Clark RP Conceição AS Crisp M Cubas P Delgado-Salinas A Dexter KG Doyle JJ Duminil J Egan A Nde la Estrella M Falcão MJ Filatov DA Fortuna-Perez AP Fortunato RH Gagnon E Gasson P Rando J Gde Azevedo Tozzi AMG Gunn B Harris D Haston E Hawkins JA Herendeen PS Hughes CE Iganci JRV Javadi F Kanu SA Kazempour-Osaloo S Kite GC Klitgaard BB Kochanovski FJ Koenen EJM Kovar L Lavin Mle Roux M Lewis G Pde Lima HCLópez-Roberts MC Mackinder B Maia VH Malécot V Mansano VF Marazzi B Mattap · doi ↗
- 2Greiner S Lehwark P Bock R (2019) Organellar Genome DRAW (OGDRAW) version 1.3.1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Research 47(W 1): W 59–W 64. 10.1093/nar/gkz 238PMC 660250230949694 · doi ↗ · pubmed ↗
- 3Huang PH Ohashi H (2010) Ohwia H. Ohashi. In: Wu ZY, Raven PH, Hong DY (Eds) Flora of China, vol. 10. Pp. 267–269. Science Press, Beijing & Missouri Botanical Garden Press, St. Louis.
- 4IUCN (2022) Guidelines for Using the IUCN Red List Categories and Criteria. Version 15. Prepared by the Standards and Petitions Committee. http://www.iucnredlist.org/documents/Red List Guidelines.pdf
- 5Jin DP Choi IS Choi BH (2019) Plastid genome evolution in tribe Desmodieae (Fabaceae: Papilionoideae). P Lo S ONE 14(6): e 0218743. 10.1371/journal.pone.0218743 PMC 659082531233545 · doi ↗ · pubmed ↗
- 6Jin JJ Yu WB Yang JB Song Yde Pamphilis CW Yi TS Li DZ (2020) Get Organelle: A simple and fast pipeline for de novo assembly of a complete circular chloroplast genome using genome skimming data. Genome Biology 21: 241. 10.1186/s 13059-020-02154-5PMC 748811632912315 · doi ↗ · pubmed ↗
- 7Katoh K Standley DM (2013) MAFFT multiple sequence alignment software version 7: Improvements in performance and usability.Molecular Biology and Evolution 30(4): 772–780. 10.1093/molbev/mst 01023329690 PMC 3603318 · doi ↗ · pubmed ↗
- 8Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper A Markowitz S Duran C Thierer T Ashton B Meintjes P Drummond A (2012) Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data.Bioinformatics 28(12): 1647–1649. 10.1093/bioinformatics/bts 19922543367 PMC 3371832 · doi ↗ · pubmed ↗