Non-lichenized Cytosporella, including C.fuligomixta sp. nov., and related plant-associated and fungicolous genera are close to foliicolous, lichenized fungi (Ascomycota, Graphidales)

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
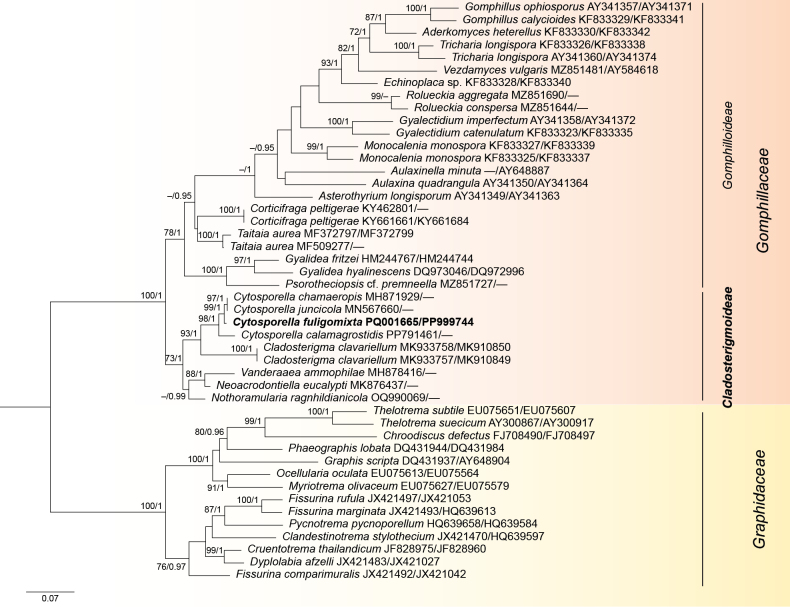 Figure 1
Figure 1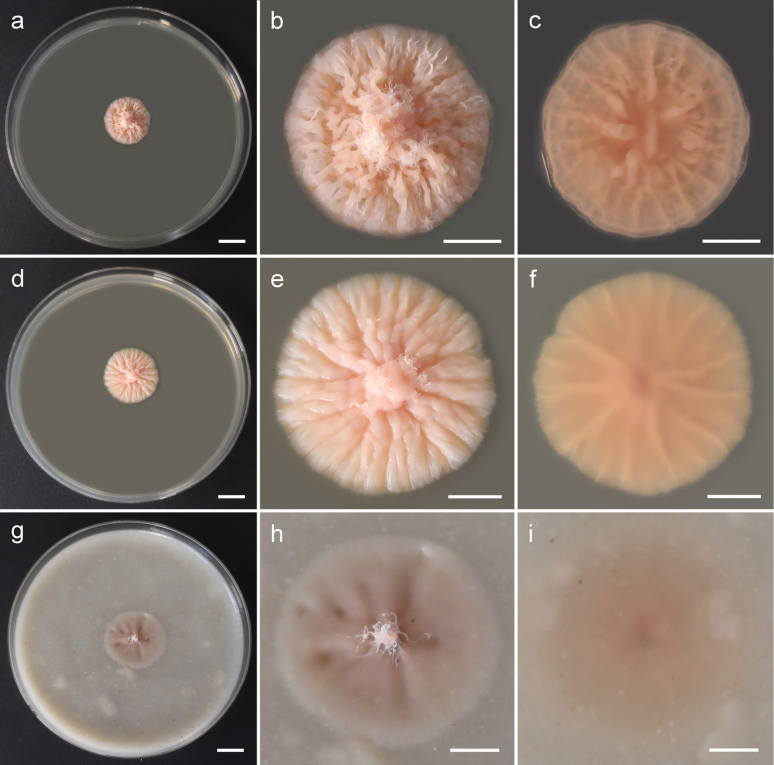 Figure 2
Figure 2 Figure 3
Figure 3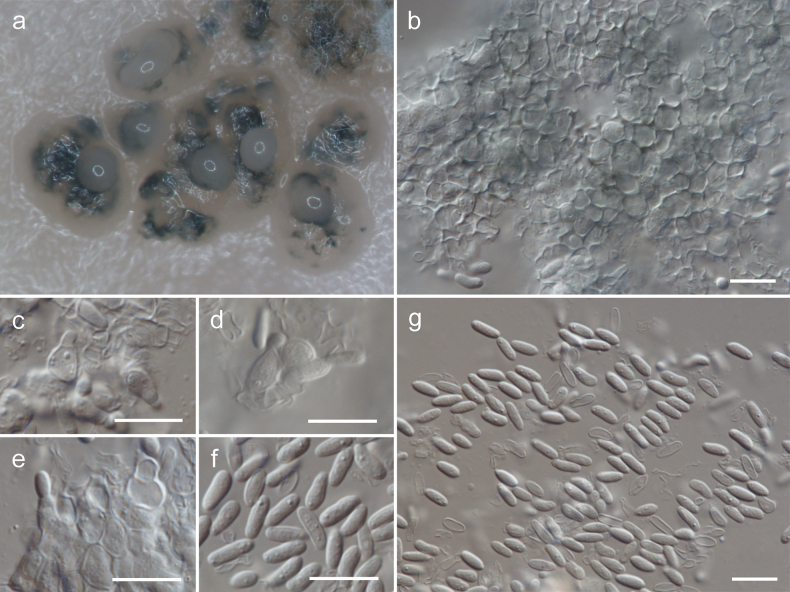 Figure 4
Figure 4| Species | Country | Host/substrate | Strain/voucher | GenBank acc. no. | References | |
|---|---|---|---|---|---|---|
|
|
| |||||
|
| Brazil | – | Cáceres & Aptroot 11953 |
|
| I. Schmitt, T. Lumbsch, E. Kraichak (unpubl.) |
|
| Costa Rica | – | Lücking s.n., F sample no. 4 |
|
|
|
|
| Costa Rica | – | Lücking s.n., F sample no. 66 |
|
|
|
|
| Costa Rica | – | E. Baloch HK2 (GZU) | – |
|
|
|
| Thailand | – | Papong 5118 |
|
| – |
|
| Brazil | UB 23227 |
|
|
| |
|
| Brazil | UB 23228 |
|
|
| |
|
| Nicaragua | – | Lücking 28636 |
|
| |
|
| Luxembourg |
| Marson 2015-05-02-1 |
| – | H.-O. Baral, G. Marson (unpubl.) |
|
| India |
| Zhurbenko 1353 (LE 260537) |
|
|
|
|
| Thailand | – | Lumbsch 19955d |
|
|
|
|
| Netherlands |
| CPC 46236 |
| – |
|
|
| Poland | sooty mould community on | G107 = |
|
| this study |
|
| USA |
| CPC 38040 |
| – |
|
|
| Italy |
|
| – |
| |
|
| USA | – | Lücking 26509a |
|
|
|
| Nicaragua | – | Lücking 28550 |
|
| I. Schmitt, T. Lumbsch, E. Kraichak (unpubl.) | |
|
| El Salvador | – | Lücking 28103 |
|
|
|
|
| Thailand | – | Lücking 24122 |
|
| |
|
| Fiji | – | Lumbsch 20521l |
|
|
|
|
| UK (Scotland) | – | Lumbsch 20100d (F) |
|
| I. Schmitt, T. Lumbsch, E. Kraichak (unpubl.) |
|
| Costa Rica | – | Will-Wolf 10006a (F) |
|
|
|
|
| Germany | – | Staiger 982 |
|
| – |
|
| Costa Rica | – | Lücking 032b |
|
| I. Schmitt, T. Lumbsch, E. Kraichak (unpubl.) |
|
| Costa Rica | – | Lücking s.n., F sample no. 2 |
|
|
|
|
| Sweden | – | Nordin 6022 (UPS) |
|
|
|
|
| Costa Rica | – | AFTOL-ID 332 |
|
|
|
|
| Costa Rica | – | Lücking 032h |
|
| I. Schmitt, T. Lumbsch, E. Kraichak (unpubl.) |
|
| Costa Rica | – | Lücking 032e |
|
| I. Schmitt, T. Lumbsch, E. Kraichak (unpubl.) |
|
| Australia | – | Lumbsch 19113f |
|
| – |
|
| Malaysia |
|
| – |
| |
|
| Germany |
| – |
| ||
|
| Australia | – | Mangold 33a |
|
| – |
|
| Bermuda | – | Berger 19598 |
|
| – |
|
| Cuba | – | Lücking et al. 41885b |
| – |
|
|
| USA | – | Lücking 26545 |
|
|
|
|
| Brazil | – | Cáceres & Aptroot 28665a |
| – |
|
|
| Brazil | – | Xavier-Leite et al. 2803 |
| – |
|
|
| Kenya |
| Kirika 5103 (EA) |
|
|
|
|
| Kenya |
| Rikkinen 16259 |
| – |
|
|
| Australia | – | Mangold 3j (F) |
|
|
|
|
| Turkey | – | Palice (ESS 21521) |
|
|
|
|
| Costa Rica | – | Lücking 033a |
|
| I. Schmitt, T. Lumbsch, E. Kraichak (unpubl.) |
|
| Costa Rica | – | Lücking s.n., F sample no. 37 |
|
|
|
|
| Netherlands | dead leaves of |
| – |
| |
|
| Brazil/Costa Rica | – | Xavier-Leite 1476/AFTOL ID 105 |
|
| |
- —Narodowe Centrum Nauki 501100004281 https://ror.org/03ha2q922 http://doi.org/10.13039/501100004281
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLichen and fungal ecology · Plant Pathogens and Fungal Diseases · Mycorrhizal Fungi and Plant Interactions
Introduction
The genus Cytosporella includes non-lichenized, plant associated fungi producing eustromatic conidiomata, phialidic conidiophores and hyaline, ellipsoid conidia (Sutton 1980; Crous et al. 2019b; Li et al. 2020). Index Fungorum (2024) includes 69 names assigned to Cytosporella but DNA sequence data are available only for three species: Cytosporellacalamagrostidis, C.chamaeropis and C.juncicola. The type species is Cytosporellasycina (Clements and Shear 1931; Sutton 1980; Li et al. 2020). It has been described from branches of Ficuscarica in France (Saccardo 1880) but not sequenced yet, making current taxonomy of the genus tentative. The species of Cytosporella are saprobic or parasitic on different hosts, mostly on branches of deciduous trees (Li et al. 2020; species data in Index Fungorum 2024). During our surveys of sooty mould communities on ornamental woody plants in southern Poland a new species of Cytosporella (described here as Cytosporellafuligomixta) was isolated from a sooty mould colony on leaves of Quercusrobur (Fagaceae).
The sequenced species of Cytosporella were shown to be related to three non-lichenized, plant associated or fungicolous genera: Neoacrodontiella, Nothoramularia and Vanderaaea, and altogether they were assigned to the family Acarosporaceae and order Acarosporales (Crous et al. 2019a, 2019b, 2021, 2023). This order and family contain saxicolous and terricolous lichenized fungi in the subclass Acarosporomycetidae of the Lecanoromycetes (Reeb et al. 2004; Miadlikowska et al. 2014; Westberg et al. 2015). Our initial query of sequences of Cytosporella (including new species C.fuligomixta), Neoacrodontiella, Nothoramularia and Vanderaaea in GenBank showed that most closely related sequences belong to members of the lichenized family Gomphillaceae. Also, non-lichenized, fungicolous Cladosterigmaclavariellum, which was recently included in Gomphillaceae (Guterres et al. 2020), was amongst the resultant related sequences. This family includes mostly foliicolous lichens and is included either in Graphidales or Ostropales within subclass Ostropomycetidae of the Lecanoromycetes (Baloch et al. 2010; Miadlikowska et al. 2014; Kraichak et al. 2018). Indeed, a recent phylogenetic tree, based on LSU sequences, published in Crous et al. (2024), placed Cladosterigma, Cytosporella, Neoacrodontiella and Nothoramularia inside the family Gomphillaceae. The genus Vanderaaea was not included in these analyses. Similarly, multi-locus (using mtSSU, SSU, LSU, ITS, rpb2 and tef1 sequences) and two-locus (using LSU and mtSSU sequences) trees, published by Guterres et al. (2020), placed Cladosterigma inside the family Gomphillaceae with Vezdamycesvulgaris (syn. Gyalideopsisvulgaris) being the most basal species. However, in these cases (based on GenBank accession numbers of sequences included by Guterres et al. 2020 in their tables), sequences of “Gyalideopsisvulgaris” used in the multi-locus tree belonged to Gyalideapraetermissa that is a member of Gyalectaceae (Ertz et al. 2021), while LSU sequence used in two-locus tree belonged to other species (closest hits in GenBank are species of Eurotiomycetes) that could have been responsible for such a result.
In this study, we describe and illustrate a new species of Cytosporella isolated from a sooty mould colony on leaves of Quercusrobur. Additionally, based on publicly available sequences, we reassess phylogenetic placement of species of Cladosterigma, Cytosporella, Neoacrodontiella, Nothoramularia and Vanderaaea using two-locus (LSU, mtSSU) sequence analyses.
Materials and methods
Strain and morphological analyses
The strain was obtained from sooty mould communities during the study of sooty moulds on ornamental woody plants cultivated in municipal greenery in southern Poland (Piątek et al. 2023). Macroscopic features of cultures were documented using 2-week-old colonies grown on malt extract agar (MEA – Blakeslee’s formula), potato dextrose agar (PDA) and oatmeal agar (OA) at 6 °C, 15 °C and 25 °C. Growth at different temperatures was assessed by measuring the colony diameter after 2 weeks and 4 weeks. Microscopic features were studied using colonies grown on MEA and OA at 15 °C after 7 weeks and 6 weeks, respectively. The characteristics of hyphae were observed on MEA and characteristics of conidiomata, conidiophores and conidia were studied on OA. Hyphae taken from the edge of the colony and mature conidiomata were mounted in lactic acid (80%) on microscope slides and analysed under Nikon Eclipse 80i light microscope. Microscopic structures were measured and photographed using NIS‐Elements BR 3.0 imaging software. Holotype is a dried specimen obtained from culture and is stored in the fungal collection of the W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków (KRAM F). Culture is deposited in the culture collection of the Westerdijk Fungal Biodiversity Institute (CBS) and in the W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków.
DNA isolation, amplification and sequencing
Genomic DNA was extracted from four-week-old MEA culture using DNeasy® Plant Mini Kit (Qiagen, Germany), according to the manufacturer’s protocol. A total of five loci were amplified: ITS1‐5.8S‐ITS2 rRNA (= ITS), partial large subunit rRNA (28S D1–D2 = LSU), small subunit mtRNA (=mtSSU), partial DNA-directed RNA polymerase II second largest subunit (rpb2) and translation elongation factor 1-alpha (tef1). To amplify the regions of ITS, LSU, mtSSU, rpb2 and tef1, five different primer pairs were used, namely ITS1–ITS4 (White et al. 1990), LSU1Fd–LR5 (Vilgalys and Hester 1990; Crous et al. 2009), mrSSU1–mrSSU3R (Zoller et al. 1999), fRPB2-5F–fRPB2-7cR (Liu et al. 1999) as well as EF1-983F and EF1-2218R (Rehner and Buckley 2005), respectively. Polymerase chain reaction mixtures were performed in a total volume of 25 μL as explained in Piątek et al. (2023). Amplification conditions for ITS and LSU were described in Czachura et al. (2021), while conditions for subsequent loci were set as follows: an initial denaturation at 94 °C for 3 min, followed by 35 cycles (mtSSU, rpb2) or 40 cycles (tef1) of denaturation at 94 °C for 60 sec (mtSSU, rpb2) or 30 sec (tef1); annealing at 52 °C (mtSSU) or 54 °C (rpb2) or 55 °C (tef1) for 60 sec (mtSSU) or 90 sec (rpb2) or 50 sec (tef1); and extension at 72 °C for 1 min (mtSSU, tef1) or 2 min (rpb2). The process was finished with the final extension at 72 °C for 7 min (mtSSU) or 10 min (rpb2, tef1). PCR amplifications were confirmed on 1% agarose electrophoresis gels stained with SimplySafe (EURx, Poland). Amplicons were enzymatically cleaned using Exo-BAP Mix (EURx, Poland). The same primers as given above were used in sequencing reactions which were carried out by Macrogen Europe B.V. (Amsterdam, The Netherlands). Obtained sequences were assembled and trimmed in Geneious Prime 2020.0.4. Consensus sequences were deposited in the NCBI’s GenBank nucleotide database (https://www.ncbi.nlm.nih.gov/genbank/).
Phylogenetic analyses
The affinities of obtained Cytosporella sequences and sequences of related genera and species were determined in the NCBIs GenBank nucleotide database using the megablast search tool (Zhang et al. 2000). For the phylogenetic analyses only LSU and mtSSU sequences were used due to limited sampling of reference sequences available for members of Gomphillaceae. The assembled concatenated LSU-mtSSU alignment contained available sequences of all sequenced species of Cladosterigma, Cytosporella, Neoacrodontiella, Nothoramularia and Vanderaaea, and sequences of selected members of Gomphillaceae and members of its most closely related family Graphidaceae used as an outgroup (Table 1). In the case of problematic sequences of Vezdamycesvulgaris (syn. Gyalideopsisvulgaris) used by Guterres et al. (2020), the correct sequences obtained from two different specimens were taken following Xavier-Leite et al. (2022).
Sequence reads which we obtained were checked for quality and assembled. The complete LSU and mtSSU sequences were separately aligned for each single-gene dataset using MAFFT algorithm (Katoh et al. 2005) in Geneious 11.1.5. Phylogenetic trees were constructed by using the Maximum likelihood (ML) and Bayesian inference (BI) analysis. For both ML and BI analyses, the model of DNA evolution that best fitted the dataset was determined using the ModelTest-NG v. 0.2.0 under the Bayesian Information Criterion (BIC) (Darriba et al. 2020). Maximum likelihood (ML) analyses were performed using the RAxML-NG v. 1.1.1 (Kozlov et al. 2019). Branch support was inferred with 1000 bootstrap replicates. Bayesian analyses were conducted using a MrBayes v. 3.2.6 (Ronquist et al. 2012). One million generations were run, sampling every 100 generations. Four parallel chains, one cold and three heated, were used. A consensus tree was generated after discarding the first 25% of trees as burn-in. Average standard deviations of split frequencies dropped below 0.01 at the end of the runs. The final phylogenetic trees were graphically visualised using FigTree v.1.4.3.
Results
Phylogenetic analyses
The concatenated LSU-mtSSU alignment contained sequences belonging to 41 species. The alignment comprised a total of 1779 characters (LSU: 969, mtSSU: 810), including alignment gaps. The best matching substitution models selected for single locus alignments in the ML analysis were as follows: TIM2+I+G4 for LSU and TPM3uf+I+G4 for mtSSU. The BI analysis was performed with the following substitution model: GTR+I+G4 for LSU and mtSSU. ML and BI analyses resulted in similar tree topologies. The best scoring maximum likelihood phylogenetic tree is shown on Fig. 1. Maximum likelihood bootstrap (MLB) support values above 70% and Bayesian posterior probabilities (BPP) above 0.95 are shown at the nodes.
Phylogenetic tree of selected representatives of Gomphillaceae, including all sequenced species of Cladosterigma, Cytosporella, Neoacrodontiella, Nothoramularia and Vanderaaea, obtained from a maximum likelihood analysis of the combined two-locus alignment (LSU, mtSSU). Representatives of Graphidaceae are used as an outgroup. The positions of Cytosporellafuligomixta sp. nov. and Cladosterigmoideae subfam. nov. are indicated in bold. GenBank accession numbers (LSU/mtSSU) are given after species name. Numbers above branches indicate maximum likelihood bootstrap (MLB) support values > 70% and Bayesian posterior probabilities (BPP) > 0.95, respectively (MLB/BPP). The scale bar represents the expected number of changes per site.
Representatives of the family Gomphillaceae formed strongly supported monophyletic lineage (MLB/BPP = 100/1). The strain of the new Cytosporella species clustered with members of the genus Cytosporella as sister to C.chamaeropis and C.juncicola (MLB/BPP = 98/1). Representatives of the genera Cladosterigma, Cytosporella, Neoacrodontiella, Nothoramularia and Vanderaaea formed a well-supported clade (MLB/BPP = 73/1) as a sister group to the well-supported clade (MLB/BPP = 78/1) of the remaining Gomphillaceae that contained lichenized species and two non-lichenized lichenicolous genera (Corticifraga, Taitaia).
Taxonomy
Gomphillaceae Walt. Watson ex Hafellner, Beih. Nova Hedwigia 79: 280. 1984
Gomphilloideae
Taxon classificationFungiGraphidales
Rivas Plata, Lücking & Lumbsch, Fungal Diversity 52(1): 108. 2012
1607D10D-10EF-5A0E-8565-8C5216B8C2F6
Notes.
Nominative subfamily includes current members of the family Gomphillaceae, excluding Cladosterigma, Cytosporella, Neoacrodontiella, Nothoramularia and Vanderaaea. The development of hyphophores with their diahyphae is a unique feature of this subfamily. Hyphophores with diahyphae are present in many, though not all, members of this lineage (Ferraro 2004; Lücking et al. 2004; Xavier-Leite et al. 2022, 2023).
Cladosterigmoideae
Taxon classificationFungiGraphidales
Piątek, Stryjak-Bogacka & Czachura subfam. nov.
5697C85D-A201-509C-AB0B-29324B8E92D1
857024
Etymology.
Named after the genus Cladosterigma.
Description.
Non-lichenized, plant associated or fungicolous fungi. Conidiomata sporodochial, synnematal, eustromatic or conidiophores arising directly from hyphae. Conidiophores hyaline, smooth, subcylindrical, conical, ampulliform or subglobose, branched or not, with terminal and/or intercalary conidiogenous cells, sometimes reduced to conidiogenous cells. Conidia solitary or rarely in chains, hyaline, smooth, 0–1-septate, ellipsoid, fusoid or subcylindrical. Sexual morph undetermined [based on generic descriptions in Sutton 1980; Guterres et al. 2020; Crous et al. 2019a, 2021, 2023].
Type genus.
Cladosterigma Pat.
Notes.
This subfamily includes genera Cladosterigma, Cytosporella, Neoacrodontiella, Nothoramularia and Vanderaaea. Members of this subfamily are different morphologically (absence of hyphophores with diahyphae), phylogenetically (distinct, sister lineage in molecular analyses) and ecologically (non-lichenized and non-lichenicolous species) from representatives of the nominative subfamily.
Cytosporella
fuligomixta
Taxon classificationFungiGraphidales
Piątek, Stryjak-Bogacka & Czachura sp. nov.
97681508-4D2D-5567-A418-9B6D1752CE5E
857025
Etymology.
Name refers to the isolation of this fungus from sooty mould communities.
Morphology of cultures of Cytosporellafuligomixta (strain G107 = CBS 152343) after 4 weeks of growth at 15 °C: a–c general view, upper side and reverse side of colony on MEA d–f general view, upper side and reverse side of colony on PDAg–i general view, upper side and reverse side of colony on OA. Scale bars: 10 mm (a, d, g); 5 mm (b, c, e, f, h, i).
DNA barcodes.
ITS (PQ001666), LSU (PQ001665), mtSSU (PP999744), rpb2 (PP997507) and tef1 (PP997508).
Morphology of Cytosporellafuligomixta on MEA (strain G107 = CBS 152343): a, b hyphae, white arrows show swellings and black arrow shows anastomosing hyphae c intertwined hyphae d hyphal fascicles. Scale bars: 10 µm.
Typus.
Poland • Małopolska Province, Kraków County: Kraków-Czyżyny (Park Lotników), municipal greenery (city park), isolated from sooty mould community on Quercusrobur leaves, 10 Oct. 2018, leg. M. Piątek, W. Bartoszek & P. Czachura (holotype KRAM F-59995; culture ex-type G107 = CBS 152343).
Morphology of Cytosporellafuligomixta on OA (strain G107 = CBS 152343): a conidiomata b wall of textura angularis c–e conidiophores f, g conidia. Scale bars: 10 µm.
Description.
Mycelium composed of sparsely branched, septate, hyaline, straight to curved, thin-walled hyphae, 1.0–1.5 µm wide; sometimes with swellings, 2.0–3.0 µm wide. Hyphae sometimes anastomose, intertwine or form fascicles [description on MEA]. Conidiomata flat, erumpent, separate, eustromatic, brown, disintegrating at the top during maturation, up to 500 µm diam, exuding a creamy conidial mass, partly enclosed by a wall of greenish-olive textura angularis. Conidiophores reduced to conidiogenous cells lining the inner cavity, hyaline, smooth, ampulliform or subcylindrical, phialidic, 3.5–8 × 3–5 µm. Conidia solitary, aseptate, hyaline, smooth, cylindrical, rarely slightly allantoid, apex obtuse, base bluntly rounded, (4–)5–8.5 × 2–2.5(–3) µm [description on OA].
Culture characteristics.
Colonies on MEA erumpent, spreading, convex, rosaceous, reaching 4 mm diam after 2 weeks at 6 °C, 9 mm diam after 2 weeks at 15 °C and 6 mm diam after 2 weeks at 25 °C, reaching 6 mm diam after 4 weeks at 6 °C, 18 mm diam after 4 weeks at 15 °C and 8 mm diam after 4 weeks at 25 °C, surface cerebriform, with sparse aerial mycelium, margin finely crenate. Reverse rosaceous. Colonies on PDA erumpent, spreading, umbonate, slimy rosaceous, reaching 4 mm diam after 2 weeks at 6 °C, 11 mm diam after 2 weeks at 15 °C and 4 mm diam after 2 weeks at 25 °C, reaching 7 mm diam after 4 weeks at 6 °C, 20 mm diam after 4 weeks at 15 °C and 7 mm diam after 4 weeks at 25 °C, surface with radial furrows starting from centre towards margin, with sparse aerial mycelium, margin finely crenate. Reverse rosaceous. Colonies on OA spreading, flat, rosaceous, reaching 4 mm diam after 2 weeks at 6 °C, 10 mm diam after 2 weeks at 15 °C and 5 mm diam after 2 weeks at 25 °C, reaching 8 mm diam after 4 weeks at 6 °C, 22 mm diam after 4 weeks at 15 °C and 5 mm diam after 4 weeks at 25 °C, surface with indistinct radial furrows starting from the centre towards the margin, without aerial mycelium, margin entire. Reverse rosaceous.
Notes.
Cytosporellafuligomixta is well delimited morphologically and ecologically from four other Cytosporella species described on Quercus hosts. These are Cytosporellamendax, C.pisiformis, C.quercus and C.sphaerosperma. All of them were described from branches or wood of Quercus sp. or Quercusrobur and differ from C.fuligomixta in shape and sizes of conidia. The conidia are globose-ellipsoid, hyaline, 4–5 × 3.5–4 µm in C.mendax (Saccardo 1884; Saccardo and Roumeguère 1884), globose, yellowish, 3–4 µm in C.pisiformis (Saccardo 1884), perfectly globose, hyaline, 9–12 µm in C.quercus (Saccardo and Sydow 1902), and globose and hyaline in C.sphaerosperma (Saccardo 1884).
Other than being phylogenetically distinct, Cytosporellafuligomixta differs also morphologically from three sequenced species of this genus: C.calamagrostidis, C.chamaeropis and C.juncicola (Crous et al. 2019b, 2024). Cytosporellacalamagrostidis described from old leaves of Calamagrostisarenaria has slightly shorter conidia, (5–)6–7 µm long (Crous et al. 2024), C.chamaeropis described from rotten Chamaeropshumilis has globose conidia (Passerini 1888) and C.juncicola described from culms of Juncuseffusus has slightly shorter and narrower conidia, (4–)5–6(–7) × 2 µm (Crous et al. 2019b).
Discussion
Cytosporella is an understudied genus without modern revision and only with few available DNA sequence data (Sutton 1980; van der Aa et al. 2001; Crous et al. 2019b, 2024; Li et al. 2020). The type species Cytosporellasycina has not been sequenced yet, making current taxonomy of the genus tentative. However, all sequenced species of Cytosporella form monophyletic lineage. All species assigned to this genus in Index Fungorum (2024) are described from different host plants or differ morphologically if described from the same host plant, which suggests that Cytosporella species might be host specific. Currently, sparse DNA sequence data does not exclude host specialization of members of this genus. Cytosporellafuligomixta described here from the sooty mould community on Quercusrobur leaves is well delimited morphologically and ecologically from four other species described on Quercus hosts.
The phylogenetic analyses including sequences of two loci (LSU, mtSSU) showed that sequenced species of Cytosporella, together with four other non-lichenized, plant associated or fungicolous genera, namely Cladosterigma, Neoacrodontiella, Nothoramularia and Vanderaaea, form sister group to lichenized and lichenicolous fungi in the family Gomphillaceae. Thus, these five genera belong to the family Gomphillaceae and order Graphidales and not to the family Acarosporaceae and order Acarosporales where they were previously included in most studies (Crous et al. 2019a, 2019b, 2021, 2023). The placement of Cladosterigma inside the Gomphillaceae was previously reported by Guterres et al. (2020). Recently, Crous et al. (2024) in a phylogenetic tree based on LSU sequences showed placement of these genera (except Vanderaaea that was not included in their analyses) inside the Gomphillaceae that was assigned to Ostropales. Most lichenized clades within Ostropales s.l. are also recognised as distinct orders (Graphidales, Gyalectales, Odontotrematales, Ostropales s.str. and Thelenellales) (Kraichak et al. 2018; Lücking 2019). Therefore, if that concept is accepted, Cladosterigma, Cytosporella, Neoacrodontiella, Nothoramularia and Vanderaaea belong to the family Gomphillaceae and order Graphidales. Moreover, the current analyses showed for the first time that they form distinct, sister lineage to remaining, mostly lichenized genera and species.
Biologically and ecologically, the above mentioned lineage of Gomphillaceae represents a coherent group of non-lichenized species occurring on plants or other fungi. The genus Cladosterigma contains only one species Cladosterigmaclavariellum that is fungicolous hyphomycete (hyperarasite) occurring on Phyllachora species infecting Eugenia species (Myrtaceae) in Paraguay and Brazil (Seifert and Bandoni 2001; Guterres et al. 2020). Cytosporella, as stated above, includes species forming eustromatic conidiomata and is associated with diverse host plants. The genus Neoacrodontiella is typified with Neoacrodontiellaeucalypti that produces conidiophores aggregated in sporodochia and occurs on leaves of Eucalyptusurophylla (Myrtaceae) in Malaysia (Crous et al. 2019a). The only known species in the genus Nothoramularia, namely Nothoramulariaragnhildianicola, is a fungicolous hyphomycete (hyperparasite) occurring on cercosporoid Ragnhildianaferruginea, which in turn is parasitic on Artemisiavulgaris (Asteraceae) in Germany (Crous et al. 2023). Vanderaaea is typified with Vanderaaeaammophilae that forms sporodochial conidiomata with curved, 0-1-septate conidia and occurs on dead leaves of Ammophilaarenaria (Poaceae) in the Netherlands (Crous et al. 2021).
So far, the family Gomphillaceae included mostly foliicolous lichenized fungi, which are predominantly known from the tropics (Xavier-Leite et al. 2022, 2023, 2024). Lichenized fungi from this family are unique in that they form a special type of asexual conidiomata called hyphophores, which produce diahyphae (Ferraro 2004; Lücking et al. 2004; Xavier-Leite et al. 2022, 2023). The family also contained three lichenicolous genera Corticifraga, Paragyalideopsis and Taitaia (Pino-Bodas et al. 2017; Suija et al. 2018; Lücking and Kalb 2002; Xavier-Leite et al. 2022, 2024; Diederich et al. 2024). Current confirmation or inclusion of non-lichenized, plant associated or fungicolous genera Cladosterigma, Cytosporella, Neoacrodontiella, Nothoramularia and Vanderaaea enlarge the concept of this predominantly lichenized family Gomphillaceae. Due to phylogenetic, morphological (notably: absence of hyphophores with diahyphae) and ecological characteristics a new subfamily Cladosterigmoideae is described for these five non-lichenized genera. It is worthy to note that within Graphidales a very similar situation is found in the lichenized family Graphidaceae, where two non-lichenized genera (Furcaspora, Rubikia) are now included therein as subfamily Rubikioideae (Cáceres et al. 2020), Additionally, non-lichenized genus Papilionovela is a member of the core Graphidaceae (Cáceres et al. 2020). Thus, Gomphillaceae and Graphidaceae, along with still other families of the subclass Ostropomycetidae (e.g. Stictidaceae), constitute good models to study transitions between lichenized and non-lichenized lifestyles.
Supplementary Material
XML Treatment for Gomphilloideae
XML Treatment for Cladosterigmoideae
XML Treatment for Cytosporella fuligomixta
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Baloch ELücking R Lumbsch HT Wedin M (2010) Major clades and phylogenetic relationships between lichenized and non-lichenized lineages in Ostropales (Ascomycota: Lecanoromycetes).Taxon 59(5): 1483–1494. 10.1002/tax.595013 · doi ↗
- 2Cáceres MESLücking R Schumm F Aptroot A (2020) A lichenized family yields another renegade lineage: Papilionovelaalbothallina is the first non-lichenized, saprobic member of Graphidaceae subfam. Graphidoideae.The Bryologist 123(2): 144–154. 10.1639/0007-2745-123.2.144 · doi ↗
- 3Clements RE Shear CL (1931) The Genera of Fungi, 2nd edn. H.W. Wilson, New York.
- 4Crous PW Schoch CL Hyde KD Wood AR Gueidan Cde Hoog GS Groenewald JZ (2009) Phylogenetic lineages in the Capnodiales. Studies in Mycology 64: 17–47S 7. 10.3114/sim.2009.64.02PMC 281696520169022 · doi ↗ · pubmed ↗
- 5Crous PW Carnegie AJ Wingfield MJ Sharma R Mughini G Noordeloos ME Santini A Shouche YS Bezerra JDP Dima B Guarnaccia V Imrefi I JurjevićŽKnapp DG Kovács GM MagistàD Perrone GRämäT Rebriev YA Shivas RG Singh SM Souza-Motta CM Thangavel R Adhapure NN Alexandrova AV Alfenas AC Alfenas RF Alvarado P Alves AL Andrade DA Andrade JP Barbosa RN Barili A Barnes CW Baseia IG Bellanger JM Berlanas C Bessette AE Bessette AR Biketova AY Bomfim FS Brandrud TE Bransgrove K Brito ACQ Cano-Lira JF Cantillo T Cavalcanti AD Cheewangkoon R Chikowski RS Conforto C Cordeiro TRL Craine JD Cruz R Damm Ude Oliveira RJ Vd · doi ↗ · pubmed ↗
- 6Crous PW Wingfield MJ Lombard L Roets F Swart WJ Alvarado P Carnegie AJ Moreno G Luangsaard J Thangavel R Alexandrova AV Baseia IG Bellanger JM Bessette AE Bessette AR De la Peña-Lastra S García D GenéJ Pham THG Heykoop M Malysheva E Malysheva V Martín MP Morozova OV Noisripoom W Overton BE Rea AE Sewall BJ Smith ME Smyth CW Tasanathai K Visagie CM Adamčík S Alves A Andrade JP Aninat MJ Araújo RVB Bordallo JJ Boufleur T Baroncelli R Barreto RW Bolin J Cabero J CaboňM CafàG Caffot MLH Cai L Carlavilla JR Chávez Rde Castro RRL Delgat L Deschuyteneer D Dios MM Domínguez LS Evans HC Eyssartier G Ferreira B · doi ↗ · pubmed ↗
- 7Crous PW Braun U Mc Donald BA Lennox CL Edwards J Mann RC Zaveri A Linde CC Dyer PS Groenewald JZ (2021) Redefining genera of cereal pathogens: Oculimacula, Rhynchosporium and Spermospora. Fungal Systematics and Evolution 7(1): 67–98. 10.3114/fuse.2021.07.04PMC 816596834124618 · doi ↗ · pubmed ↗
- 8Crous PW Akulov A Balashov S Boers J Braun U Castillo J Delgado MA Denman S Erhard A Gusella G JurjevićŽKruse J Malloch DW Osieck ER Polizzi G Schumacher RK Slootweg E Starink-Willemse Mvan Iperen AL Verkley GJM Groenewald JZ (2023) New and interesting fungi. 6.Fungal Systematics and Evolution 11(1): 109–156. 10.3114/fuse.2023.11.0938545457 PMC 10966675 · doi ↗ · pubmed ↗