Grewiakentingensis (Malvaceae, Grewioideae), a new species from Taiwan

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2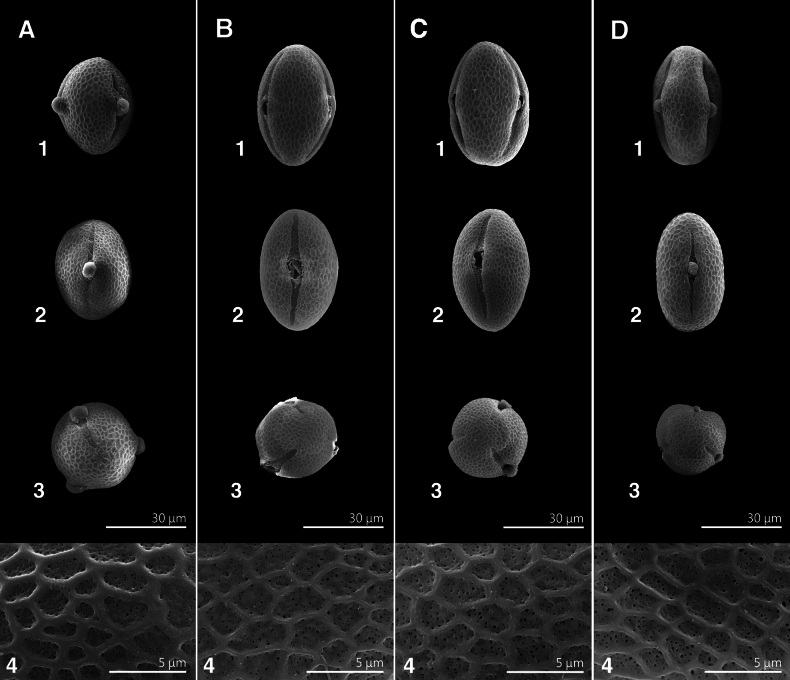 Figure 3
Figure 3 Figure 4
Figure 4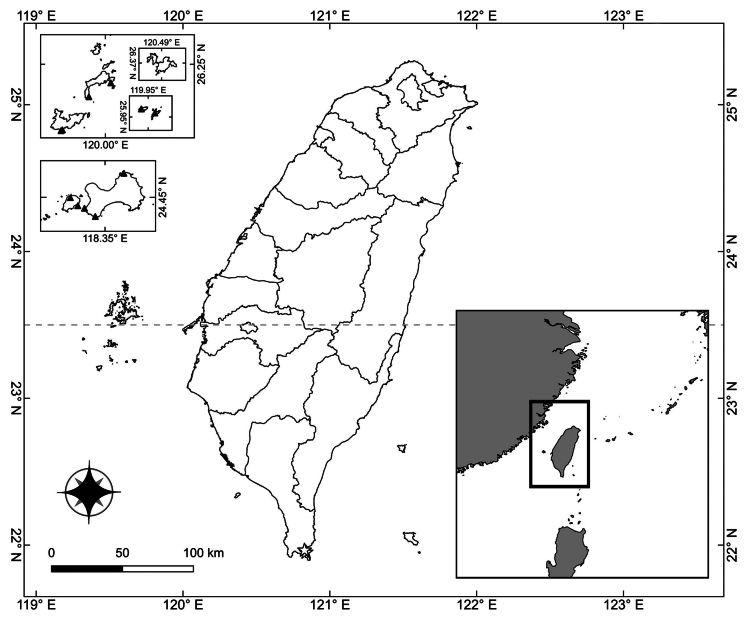 Figure 5
Figure 5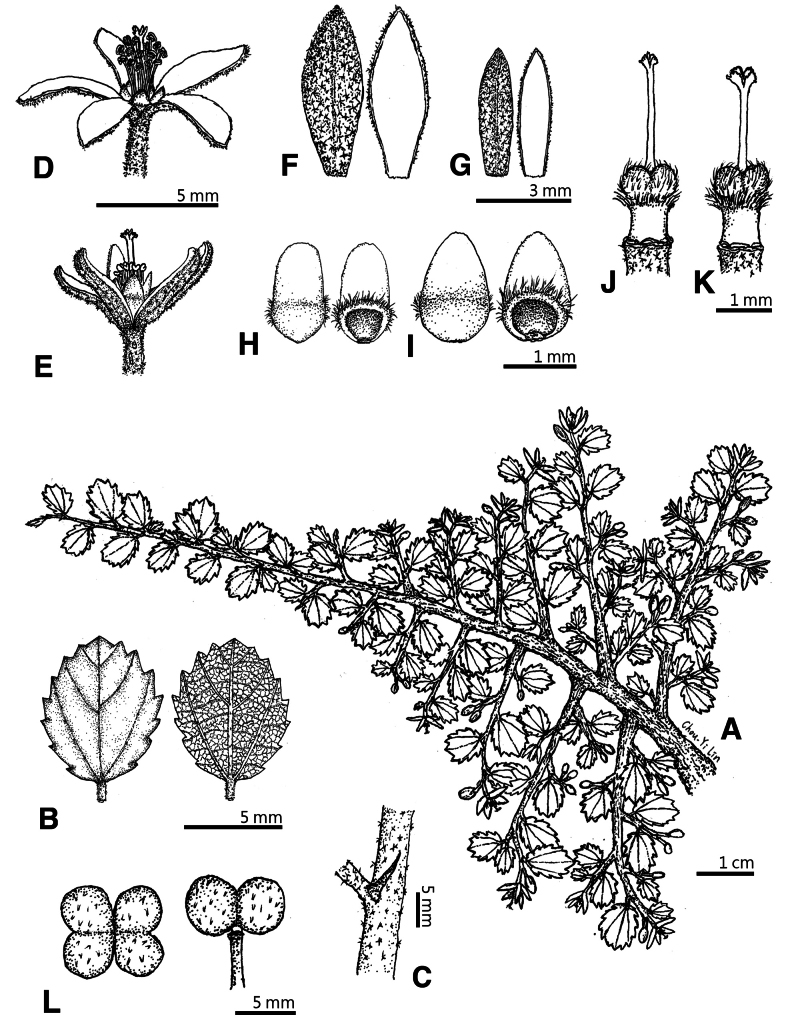 Figure 6
Figure 6 Figure 7
Figure 7 Figure 8
Figure 8 Figure 9
Figure 9| Taxon | Location | Coordinate | Altitude | Date | Voucher |
|---|---|---|---|---|---|
|
| Taiwan. Pingtung County, Hengchun Township, Fongchueisha |
| 73 m | 3 May 2024 | |
| Taiwan. Pingtung County, Hengchun Township, Sheding Formosan Sika Deer Restoration Area |
| 154 m | 26 June 2024 | ||
|
| Taiwan. Kinmen County, Jincheng Township, Zhaishan Tunnel |
| 20 m | 25 May 2024 | |
| Taiwan. Kinmen County, Jinsha Township, Mashan Observation Post |
| 4 m | 29 May 2023 | ||
| Taiwan. Lienchiang County, Nangan Township, Guanmaoshan |
| 15 m | 13 June 2023 | ||
|
| Taiwan. New Taipei City, Wanli Dist., Yehliu Geopark |
| 1 m | 22 April 2023 | |
| Taiwan. New Taipei City, Ruifang Dist., Nanyaqiyan |
| 2 m | 12 May 2024 | ||
| Taiwan. New Taipei City, Ruifang Dist., Bat cave park |
| 4 m | 27 June 2024 | ||
|
| Taiwan. Hualien County, Xiulin Township, Chongde trail |
| 24 m | 26 March 2023 | |
| Taiwan. Kaohsiung City, Mituo Dist., Ta-di Mountain Natural Park |
| 47 m | 23 July 2023 | ||
| Taiwan. Pingtung County, Chunri Township, Dahan forest road |
| 953 m | 11 August 2024 |
| Characters |
|
|
|
| |
|---|---|---|---|---|---|
| Habit | procumbent shrub, ca. 5 cm tall | erect to ascending shrub, ca. 1–2 m tall | erect to ascending shrub, ca. 1–2 m tall | small tree, ca. 3–5 m tall | |
| Leaves | |||||
| shape | broadly elliptic to elliptic | elliptic, obovate, ovate to rhomboid-ovate | rhomboid, broadly rhomboid to obtrullate | ovate, elliptic to rhomboid-ovate | |
| size (cm) | 0.76 ± 0.28c × 0.60 ± 0.25c | 3.93 ± 1.82b × 2.67 ± 1.31b | 4.12 ± 1.32b × 3.01 ± 1.23b | 6.56 ± 2.36a × 3.64 ± 1.26a | |
| margin | serrate | serrulate to biserrulate | irregularly serrulate, biserrulate to dentate | serrulate to biserrulate | |
| surfaces | Both surfaces nearly glabrous | Both surfaces nearly glabrous | Both surfaces densely covered with stellate hairs | Both surfaces nearly glabrous | |
| number of serrations | 13.83 ± 2.57c | 47.87 ± 16.00b | 49.69 ± 17.66b | 65.48 ± 24.75a | |
| stipule length (mm) | 0.78 ± 0.19b | 3.84 ± 0.73a | 3.99 ± 0.60a | 4.02 ± 0.79a | |
| Sexual system | Gynodioecy | Trioecy | Trioecy | Trioecy | |
| Inflorescences | |||||
| position | terminal | leaf-opposed, rarely axillary | leaf-opposed, rarely axillary | leaf-opposed, rarely axillary | |
| per inflorescence flower number | 2.39 ± 1.17c | 9.49 ± 2.63a | 7.41 ± 2.01b | 8.42 ± 2.19ab | |
| peduncle length (mm) | 1.49 ± 0.60b | 6.63 ± 2.61a | 6.90 ± 2.64a | 7.87 ± 2.19a | |
| pedicel length (mm) | 2.59 ± 1.21b | 6.05 ± 1.46a | 6.23 ± 1.35a | 6.57 ± 1.50a | |
| Flowers | |||||
| flower diameter (mm) | Bisexual flower | 9.49 ± 1.34c | 16.88 ± 1.71a | 15.39 ± 1.22b | 16.25 ± 1.52ab |
| Female flower | 6.75 ± 0.84c | 11.65 ± 1.29a | 11.49 ± 1.23a | 10.09 ± 0.75b | |
| sepal number | (3)4–5 | (4)5(6) | (4)5(6) | (4)5(6) | |
| sepal size (mm) | Bisexual flower | 5.34 ± 0.46c × 1.84 ± 0.27c | 8.70 ± 0.83a × 3.03 ± 0.44a | 8.15 ± 0.73b × 2.54 ± 0.43b | 7.89 ± 0.57b × 2.46 ± 0.27b |
| Female flower | 3.94 ± 0.49c × 1.29 ± 0.23c | 5.09 ± 0.34b × 1.63 ± 0.13a | 5.47 ± 0.46a × 1.51 ± 0.13b | 5.12 ± 0.46b × 1.62 ± 0.12ab | |
| petal size (mm) | Bisexual flower | 1.32 ± 0.13d × 0.75 ± 0.08c | 2.65 ± 0.28a × 1.01 ± 0.08a | 2.32 ± 0.24b × 1.00 ± 0.06a | 1.79 ± 0.19c × 0.92 ± 0.11b |
| Female flower | 1.37 ± 0.10c × 0.79 ± 0.07a | 1.93 ± 0.24a × 0.78 ± 0.13a | 1.64 ± 0.20b × 0.78 ± 0.12a | 1.87 ± 0.53a × 0.86 ± 0.20a | |
| stamens number | Bisexual flower | 29.88 ± 3.09c | 110.86 ± 4.93a | 82.89 ± 15.88b | 87.09 ± 18.74b |
| Female flower | 17.55 ± 2.58c | 79.53 ± 12.30a | 64.77 ± 8.18b | 80.67 ± 19.16a | |
| filaments length (mm) | Bisexual flower | 1.92 ± 0.56b | 4.02 ± 0.96a | 3.87 ± 1.16a | 3.61 ± 0.83a |
| Female flower | 0.76 ± 0.12c | 1.00 ± 0.27ab | 0.90 ± 0.28bc | 1.05 ± 0.37a | |
| style length (mm) | Bisexual flower | 2.44 ± 0.32c | 4.18 ± 0.58a | 3.65 ± 0.22b | 3.71 ± 0.53b |
| Female flower | 1.83 ± 0.28c | 2.25 ± 0.22ab | 2.46 ± 0.21a | 2.02 ± 0.24bc | |
| Druplet size (mm) | 4.60 ± 0.22b | 5.39 ± 0.50a | 5.43 ± 0.45a | 5.11 ± 0.36a | |
| Pollen | |||||
| polar axis (μm) | 37.61 ± 1.62b | 45.32 ± 1.34a | 45.93 ± 1.65a | 44.65 ± 1.27a | |
| equatorial axis (μm) | 28.35 ± 2.47bc | 29.07 ± 1.05ab | 29.90 ± 1.38a | 26.89 ± 1.74c | |
| P/E ratio | 1.33 ± 0.10c | 1.56 ± 0.06b | 1.54 ± 0.11b | 1.67 ± 0.11a | |
| shape | subprolate to prolate | prolate | prolate | prolate | |
| lumen size (μm) | 1.98 ± 0.56b | 2.47 ± 0.77a | 2.40 ± 0.53a | 2.24 ± 0.72ab | |
| muri width (μm) | 0.68 ± 0.06a | 0.56 ± 0.09b | 0.55 ± 0.07b | 0.54 ± 0.06b | |
| perforation size (μm) | 0.11 ± 0.05a | 0.11 ± 0.06a | 0.11 ± 0.06a | 0.12 ± 0.08a | |
| Distribution | Endemic to Taiwan; restricted to elevated coral reefs and coastal grasslands of the eastern coast of Hengchun Peninsula. | Distributed along the coast in Fujian and Hainan provinces in China. In Taiwan, found in Kinmen and Lienchiang counties. | Endemic to Taiwan; distributed along the coast and hills of northern to central Taiwan. | Distributed in central and southern China. At altitudes of ca. 10–1000 m in southern and eastern Taiwan. | |
| Characters |
|
|
|
|---|---|---|---|
| Habit | procumbent shrub, ca. 5 cm tall | shrub or tree, ca. 1–4 m tall | shrub or tree, ca. 1–4 m tall |
| Leaves | |||
| shape | broadly elliptic to elliptic | ovate | ovate |
| size (cm) | 0.4–1.6 × 0.3–1.3 | 1.0–2.5 × 0.9–1.5 | 3.0–11.5 × 2.0–7.0 |
| margin | serrate | biserrulate | biserrulate |
| surfaces | Both surfaces nearly glabrous | adaxial surface nearly glabrous, abaxial surface covered with stellate hairs | adaxial surface nearly glabrous, abaxial surface covered with stellate hairs |
| Inflorescences | solitary to cyme | umbel | umbel |
| Adaxial surface of petals | glabrous | tomentose | glabrous |
| Distribution | Endemic to Taiwan; restricted to elevated coral reefs and coastal grasslands of the eastern coast of Hengchun Peninsula. | Distributed in Sichuan and Yunnan provinces in China. | Distributed in northern to southern China and the Korean Peninsula. |
| 1a | Leaf bases cordate to rounded, usually oblique; inflorescences axillary; flowers bisexual only; sepals reflexed; fruits unlobed or 2-lobed |
|
| 2a | Abaxial surface of leaves tomentose; petals and stamens turn from yellow to orange before withering; fruit surface tomentose |
|
| 2b | Abaxial surface of leaves nearly glabrous; petals and stamens turn from yellow to red before withering; fruit surface pubescent |
|
| 1b | Leaf bases cuneate, obtuse to rounded, seldom oblique; inflorescences terminal or rarely axillary; flowers unisexual or bisexual; sepals erect or slightly reflexed; fruits usually 2- to 4-lobed |
|
| 3a | Procumbent shrubs; leaf blade length < 2 cm; gynodioecious; inflorescences terminal, 1–3(4–6) flowers per peduncle; stamens < 40 per flower |
|
| 3b | Trees or erect to ascending shrubs; leaf blade length often > 2 cm; trioecious; inflorescences leaf-opposed, 4–15 flowers per peduncle, rarely axillary; stamens > 50 per flower |
|
| 4a | Leaves densely covered with stellate hairs above and below |
|
| 4b | Leaves surfaces nearly glabrous above and below |
|
| 5a | Small trees ca. 3–5 m tall; leaf blades ovate to elliptic, sometimes rhomboid-ovate, maximum length > 10 cm |
|
| 5b | Erect to ascending shrubs ca. 1–2 m tall; leaf blades elliptic, obovate, or ovate to rhomboid-ovate, maximum length < 10 cm |
|
| Latitude / Longitude | Specimens |
|---|---|
|
| |
|
| |
|
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Diversity and Evolution · African Botany and Ecology Studies · Plant and animal studies
Introduction
Grewia L. (Malvaceae: Grewioideae) is comprised of 280–300 species, which are distributed in the Old World tropics (Chung et al. 2003, 2005; Randrianasolo et al. 2013). The genus includes trees, shrubs, and climbers, which are characterized by leaves that are 3- to 5-nerved from the base; solitary, cymose, or umbellate inflorescences that are axillary, leaf-opposed, or terminal; and numerous free stamens, an androgynophore, and unlobed or 2- to 4-lobed drupaceous fruits (Liu and Lo 1993; Kubitzki and Bayer 2003; Tang et al. 2007). According to Liu and Lo (1993), four species have been recorded in Taiwan; G.biloba G. Don, G.eriocarpa Juss., G.piscatorum Hance, and G.rhombifolia Kaneh. & Sasaki. Chang et al. (2018) later reported an occurrence of G.tiliifolia Vahl.
On the basis of floral and fruit morphology, Burret (1926) subdivided Grewia into four sections but his classification system is probably artificial (Dorr, pers. comm.). Nonetheless, the Taiwanese taxa can be placed in two of Burret’s sections. Grewiaeriocarpa and G.tiliifolia, which are characterized by axillary inflorescences, bisexual flowers, entire stigmas lobes, and often unlobed fruits, can be assigned to sect. Axillares Burret while G.biloba, G.piscatorum, and G.rhombifolia, characterized by leaf-opposed, terminal, or rarely axillary inflorescences, unisexual or bisexual flowers, stigmas lobes with filamentary divisions, and usually 4-lobed fruits can be assigned to sect. Glomeratae Burret (see also Chang et al. 2018).
During recent field and herbarium investigations, we noticed that populations of Grewia growing on elevated coral reefs in Hengchun Peninsula, Taiwan had been identified as G.piscatorum based on sparse hairs on both leaf surfaces and white sepals. However, individuals in this locality diverged from G.piscatorum in several characters, including their procumbent habit, smaller stipules, leaves with fewer serrations, smaller flowers, shorter pedicels, and fewer stamens. After careful comparison, we concluded that the material from Hengchun represents a new species, which is described here as G.kentingensis.
Materials and methods
Morphological comparison
We compared our unknown taxon to Grewiapiscatorum. Additionally, we compared it to the Taiwanese taxa G.rhombifolia and G.bilobavar.biloba as well as to the non-Taiwanese taxa G.bilobavar.parviflora (Bunge) Hand.-Mazz. and G.bilobavar.microphylla (Maxim.) Hand.-Mazz., all of which can be assigned to sect. Glomeratae. Morphological measurements were conducted using both fresh and dried material. Quantitative characters were measured using fresh material, while dry specimens only were used for qualitative character observations and descriptions. For each taxon, at least three individuals were used for measurements and statistical tests. Morphological descriptions follow Liu and Lo (1993); Wahlert et al. (2015); and Barrett (2019).
Herbarium resources
Herbarium acronyms from Index Herbariorum were used in this study (Thiers 2024, continuously updated). Voucher specimens collected for the current study were deposited in the herbarium of the Department of Forestry, National Chung Hsing University, Taiwan (TCF). Physical and digital specimens in several herbaria also were examined; physical specimens: Department of Forestry and Natural Resources, National Chia-Yi University, Taiwan (CHIA), Provincial Pingtung Institute (PPI), National Taiwan University, Taiwan (TAI), Endemic Species Research Institute, Taiwan (TAIE), Taiwan Forestry Research Institute (TAIF), TCF, and National Museum of Natural Science, Taiwan (TNM); and digital specimens: SSchool of Life Sciences, Xiamen University (AU), The Natural History Museum (BM), South China Botanical Garden, Chinese Academy of Sciences (IBSC), and the Royal Botanic Gardens, Kew (K).
Pollen morphology
Pollen grains were acetolyzed following the method established by Erdtman (1960). Grains were sequentially dehydrated in ethanol and then critical point dried. The dried pollen grains were mounted on a stub and sputter-coated with gold in a Quorum SC7620 sputter coater (Quorum Technologies, Laughton, UK) for 120 s. Subsequently, the grains were observed under a Hitachi S-3400N scanning electron microscope (Hitachi, Ltd, Tokyo, Japan). At least thirty pollen grains were observed and measured for each taxon. The terminology for the morphological descriptions of pollen grains is in accordance with the terminology used by Erdtman (1952) and Halbritter et al. (2018). Information on voucher specimens is provided in Table 1.
Distribution map
A distribution map for this species was generated using QGIS ver. 3.24.2 (QGIS.org 2024) with the package developed by Lin (2018).
Data analysis
Quantitative morphological characters of taxa were measured and means and standard deviations were calculated (Table 2). Differences among taxa were analyzed using one-way analysis of variance, followed by Tukey’s Honestly Significant Difference (HSD) multiple-range test (p ≤ 0.05) (Oliveira 2013). All statistical analyses were conducted using SPSS ver. 20 (Jasrai 2020).
Results and discussion
Macromorphological differences
Grewiakentingensis can be assigned to sect. Glomeratae in the infrageneric classification system proposed by Burret (1926), as can G.piscatorum, G.rhombifolia, and G.bilobavar.biloba. The species assigned to sect. Glomeratae in Taiwan have three different habits: 1) G.bilobavar.biloba is a small tree that is ca. 3–5 m tall and usually has a distinct main trunk, 2) G.piscatorum and G.rhombifolia are erect to ascending shrubs 1–2 m tall, 3) G.kentingensis is a procumbent shrub that is clearly distinct from other taxa (Fig. 1, Table 2). Both leaf surfaces in G.rhombifolia are densely covered with stellate hairs, whereas both leaf surfaces in G.kentingensis, G.piscatorum, and G.bilobavar.biloba are nearly glabrous (Table 2). The leaf blades of Grewia taxa in Taiwan have distinct shapes: G.kentingensis has broadly elliptic to elliptic leaves, G.piscatorum displays elliptic, obovate, ovate, and rhomboid-ovate leaves, G.bilobavar.biloba features ovate, elliptic, and rhomboid-ovate leaves, and G.rhombifolia has rhomboid, broadly rhomboid, and obtrullate leaves (Table 2). Among the Grewia taxa in Taiwan, G.kentingensis has the smallest leaves (p ≤ 0.05). The leaves of G.kentingensis measure 0.4–1.6 × 0.3–1.3 cm. The leaves of G.piscatorum measure 0.7–7.8 × 0.5–5.5 cm. The leaves of G.rhombifolia measure 1.6–6.7 × 1.1–7.0 cm. The leaves of G.bilobavar.biloba measure 2.1–13.9 × 1.3–7.4 cm (Table 2). Grewiapiscatorum exhibits considerable variation in leaf size, partially overlapping with G.kentingensis (Table 2). The leaf margin in G.kentingensis is serrate, and that of G.piscatorum and G.bilobavar.biloba are serrulate to biserrulate. The leaf margin in G.rhombifolia is irregularly serrulate or biserrulate to dentate. Grewiakentingensis features significantly fewer leaf serrations (9–20) (p ≤ 0.05) than G.piscatorum (23–82), G.rhombifolia (26–88), or G.bilobavar.biloba (29–132) (Table 2).
Habit of taxa assigned to Grewiasect.Glomeratae Burret in Taiwan. AGrewiakentingensis Y.H.Tseng, Chih Y.Chang & C.Y.Lin BGrewiapiscatorum Hance CGrewiarhombifolia Kaneh. & Sasaki DGrewiabilobaG. Donvar.biloba.
Significant differences also were observed in the length of peduncles and pedicels among the Grewia taxa measured (p ≤ 0.05). Grewiakentingensis features significantly shorter peduncles and pedicels (peduncles: 0.7–2.7 mm, pedicels: 0.9–4.9 mm) (p ≤ 0.05) compared with G.piscatorum (peduncles: 3.1–17.2 mm, pedicels: 3.5–8.8 mm), G.rhombifolia (peduncles: 3.0–13.1 mm, pedicels: 3.8–8.6 mm), and G.bilobavar.biloba (peduncles: 3.7–14.3 mm, pedicels: 4.4–9.3 mm) (Table 2). Regarding the sexual or breeding system, G.kentingensis is gynodioecious, whereas the other taxas are all trioecious (Table 2). Flower diameter in G.kentingensis (7.2–11.7 mm in bisexual flowers and 5.2–7.8 mm in female flowers) is significantly smaller (p ≤ 0.05) than it is in G.piscatorum (14.8–18.5 (–20.2) mm in bisexual flowers and 9.4–14.6 mm in female flowers), G.rhombifolia (13.9–17.8 mm in bisexual flowers and 8.9–13.8 mm in female flowers), or G.bilobavar.biloba (13.9–18.7 mm in bisexual flowers and 8.8–11.3 mm in female flowers) (Table 2). Moreover, the sepals and petals in G.kentingensis are notably shorter than those in G.piscatorum, G.rhombifolia, or G.bilobavar.biloba (Table 2). Grewiakentingensis also features significantly fewer stamens (25–34 in bisexual flowers and 14–23 in female flowers) (p ≤ 0.05) compared with G.piscatorum (103–118 in bisexual flowers and 58–102 in female flowers), G.rhombifolia (64–103 in bisexual flowers and 51–80 in female flowers), and G.bilobavar.biloba (65–121 in bisexual flowers and 55–98 in female flowers (Table 2). Additionally, the filaments and styles of bisexual flowers in G.kentingensis are significantly shorter (p ≤ 0.05) than those in other taxa (Table 2).
Finally, although all the taxa assigned to sect. Glomeratae in Taiwan exhibit 4-lobed fruits with nearly glabrous surfaces that turn red after maturation, drupelet size in Grewiakentingensis (4.1–5.2 mm) is significantly smaller (p ≤ 0.05) than those in G.piscatorum (4.4–6.4 mm), G.rhombifolia (4.7–6.3 mm), or G.bilobavar.biloba (4.5–5.6 mm) (Table 2).
Other taxa that are morphologically similar to Grewiakentingensis and assigned to sect. Glomeratae but not found in Taiwan include G.bilobavar.parviflora, which is distributed from northern to southern China and the Korean Peninsula (Chang 1989; Tang et al. 2007; Chang et al. 2014) and G.bilobavar.microphylla, which is distributed in Sichuan and Yunnan provinces in China (Chang 1989; Wu et al. 1995; Tang et al. 2007). They all exhibit common characteristics such as white sepals, unisexual or bisexual flowers, and 4-lobed drupes that turn red upon maturation (Chang 1989; Tang et al. 2007). However, these two varieties are shrubs or trees 1–4-m tall (Chang 1989; Tang et al. 2007), whereas G.kentingensis is a procumbent shrub (Table 3). In addition, G.kentingensis can be readily distinguished from G.bilobavar.parviflora by its broadly elliptic to elliptic (vs. ovate) leaf blade shape, smaller leaf blade size (0.4–1.6 × 0.3–1.3 cm vs. 3.0–11.5 × 2.0–7.0 cm), nearly glabrous (vs. abaxial surface covered with stellate hairs) leaf surfaces, and solitary to cymose (vs. umbellate) inflorescences (Bunge 1835; Chang 1989; Tang et al. 2007) (Table 3). The leaf blade size in G.bilobavar.microphylla is smaller (1.0–2.5 × 0.9–1.5 cm), and its range overlaps with that of G.kentingensis (0.4–1.6 × 0.3–1.3 cm) (Table 3). Grewiabilobavar.microphylla has ovate leaf blades with an acute to obtuse apex, stellate hairs covering the abaxial surface of leaves, and tomentum on the adaxial surface of its petals (Maximowicz 1889; Chang 1989; Tang et al. 2007). Grewiakentingensis features broadly elliptic to elliptic leaf blades with a rounded to obtuse apex, nearly glabrous leaf surfaces, and glabrous adaxial surface on its petals (Table 3).
Pollen morphology
Pollen grains of taxa assigned to sect. Glomeratae in Taiwan are medium-sized and tricolporate, subprolate, or prolate (Fig. 3, Table 2). These observations are consistent with those provided by Huang (1972) and Hsieh and Huang (1983). Grewiakentingensis has a significantly shorter polar axis (p ≤ 0.05) (35.8–41.4 μm) than G.piscatorum (42.8–47.8 μm), G.rhombifolia (42.8–48.2 μm), or G.bilobavar.biloba (42.5–46.9 μm) (Fig. 3, Table 2). The P/E ratio in G.kentingensis is the smallest (1.1–1.5) and is significantly lower (p ≤ 0.05) than that in the other species (Fig. 3, Table 2). In addition, pollen grains of G.kentingensis are subprolate to prolate in shape, whereas those of the other taxa are prolate (Fig. 3, Table 2). The exine in all these taxa is reticulate, with perforations in the lumen (Fig. 3). Perforation size is not significantly different among the four species (Fig. 3, Table 2). Lumen size in G.kentingensis (1.0–2.9 μm) is significantly smaller (p ≤ 0.05) than that in G.piscatorum (1.1–4.0 μm) or G.rhombifolia (1.4–3.5 μm). However, no significant difference was observed in the lumen size between G.kentingensis and G.bilobavar.biloba (1.1–3.6 (–4.0) μm) (Fig. 3, Table 2). Muri width in G.kentingensis (0.6–0.8 μm) is significantly larger (p ≤ 0.05) than that in G.piscatorum (0.4–0.7 μm), G.rhombifolia (0.4–0.7 μm), or G.bilobavar.biloba (0.4–0.7 μm) (Fig. 3, Table 2). In conclusion, G.kentingensis pollen can be distinguished from that of other taxa by its shape, smaller polar axis, smaller P/E ratio, smaller lumen size, and wider muri.
Comparison of morphological characters. AGrewiakentingensis Y.H.Tseng, Chih Y.Chang & C.Y.Lin BGrewiapiscatorum Hance 1 leaves 2 bisexual inflorescence 2' female inflorescence 3 bisexual flower 3' female flower 4 sepal (bisexual) 4' sepal (female) 5 petal (bisexual) 5' petal (female) 6 bisexual flower (sepals and petals removed) 6' female flower (sepals and petals removed) 7 fruit (left: overhead view; right: side view).
Comparison of pollen morphology of four Grewia taxa. AGrewiakentingensis Y.H.Tseng, Chih Y.Chang & C.Y.Lin BGrewiapiscatorum Hance CGrewiarhombifolia Kaneh. & Sasaki DGrewiabilobaG. Donvar.biloba1 equatorial view 2 colporate view 3 polar view 4 surface sculpture.
Re-evaluation of Grewiapiscatorum in Kinmen and Lienchiang counties, Taiwan
Grewiapiscatorum was described by Hance (1866), who based the name on a collection by Swinhoe made on Lamyet Island (Nanjeih Island) in Fujian Province, China (Swinhoe in Herb. Hance 6527). During our field trip to Kinmen and Lienchiang counties in Taiwan and based on examination of specimens, we found that the Grewia in Kinmen and Lienchiang counties closely resembled the type specimen of G.piscatorum. Additionally, the Grewia populations found in Kinmen and Lienchiang counties are geographically close to the type locality of G.piscatorum; both counties located on islands along the coast of Fujian Province.
In previous studies, the Grewia in Kinmen and Lienchiang counties was identified as G.rhombifolia (Kuo 2004; Lu 2011). However, we found that this Grewia has nearly glabrous leaf surfaces on both sides, with sparse stellate hairs on the veins. Thus, it differs from G.rhombifolia, which is characterized by a dense covering of stellate hairs on both leaf surfaces. Hance (1866) stated that G.piscatorum has a small leaf blade size. After measuring the type specimen, we determined the leaf blade size to be 0.7–2.5 cm. Nevertheless, based on field observations, we found that leaf blade size varies greatly within an individual plant, differing by leaf position on the branchlet (Fig. 4). On the same branchlet, leaves near the distal end of a branchlet are usually larger, reaching 3–7 cm, whereas leaves near the proximal end of a branchlet are smaller, with a size of 0.5–2 cm. This latter leaf blade size matches that of the type specimen of G.piscatorum (Fig. 4). This variation in leaf size has also been found in Bombacoideae (Malvaceae) (Carvalho-Sobrinho et al. 2024). Additionally, such variation in leaf morphology has been observed in G.rhombifolia. Smaller leaf blade forms of G.rhombifolia in Taiwan have been identified as G.piscatorum (Sasaki 1928; Kanehira 1936; Li 1963; Liu and Lo 1993). In previous studies, leaf blade size was often used for identification (Li 1963; Liu et al. 1988; Liu and Lo 1993; Liu et al. 1998). However, variation in leaf blade size can contribute to overlap in size measurements among different taxa thereby rendering the identification of closely related taxa by this criterion alone challenging. In addition to leaf blade size, other characters such as habit, leaf vestiture, and floral morphology should be considered in comparisons for identification (Table 2).
Variation in leaf size of Grewiapiscatorum Hance. A variation in leaf size at different positions on the same branchlet B leaves near the distal end of a branchlet C leaves near the proximal end of a branchlet.
The leaf blade size in Grewiakentingensis is much more consistent, with a leaf blade size of 0.5–1.5 cm. Leaves do not exceed 2 cm, which is significantly different from leaves of G.piscatorum. In summary, based on the similarity to the type specimen and description of G.piscatorum by Hance (1866), we consider the Grewia previously identified as G.rhombifolia in Kinmen and Lienchiang counties to be G.piscatorum.
Comparison of the distributions of Grewiakentingensis and G.piscatorum
Grewiakentingensis is only found along the eastern coastline of Hengchun Peninsula. This species grows on elevated coral reefs and open coastal grasslands with intense sunlight. By contrast, G.piscatorum is found in coastal hills or islands in Fujian and Hainan provinces in China (Chun and Chang 1965; Chang 1987; Chang 1989; Tang et al. 2007). In Taiwan, G.piscatorum is only found in Kinmen and Lienchiang counties (Fig. 5).
Distribution map of Grewiakentingensis Y.H.Tseng, Chih Y.Chang & C.Y.Lin (white star) and G.piscatorum Hance (black triangle) in Taiwan.
Key to Grewia in Taiwan modified from Liu and Lo (1993) and Chang et al. (2018)
**: **
Taxonomic treatment
Grewia
kentingensis
Taxon classificationPlantaeMalvalesMalvaceae
Y.H.Tseng, Chih Y.Chang & C.Y.Lin sp. nov.
87979378-1FEA-58F4-AFC1-CF1A4D0954B9
urn:lsid:ipni.org:names:77357855-1
Diagnosis.
Grewiakentingensis is similar to G.piscatorum but can be distinguished by its procumbent (vs. erect to ascending) habit, smaller leaf size, serrate (vs. biserrulate) leaf margin, terminal (vs. often leaf-opposed) inflorescences, gynodioecious (vs. trioecious) breeding system, smaller flowers, fewer stamens, smaller fruits, and smaller pollen grains.
Grewiakentingensis Y.H.Tseng, Chih Y.Chang & C.Y.Lin. A habit B leaves C stipule D bisexual flower E female flower F sepals (bisexual) G sepals (female) H petals (bisexual) I petals (female) J gynoecium (bisexual) K gynoecium (female) L fruit (left: overhead view; right: lateral view).
Type.
Taiwan. Pingtung County • Hengchun Township, Sheding Formosan Sika Deer Restoration Area, 154 m alt., 21°57'53.0"N, 120°49'39.2"E, 26 June 2024, C. Y. Lin et al. 93 (holotype: TCF; isotype: TNM).
Grewiakentingensis Y.H.Tseng, Chih Y.Chang & C.Y.Lin. A habitat B habit C stipule D leaf blade variation E inflorescence (bisexual) F inflorescence (female) G bisexual flowers H bisexual flower (sepals and petals removed), H' gynoecium (bisexual) I sepals (bisexual) J petals (bisexual) K female flowers L female flower (sepals and petals removed), L' gynoecium (female) M sepals (female) N petals (female) O androgynophore (bisexual) O' androgynophore (female) P ovary (bisexual) P' ovary (female) Q fruit (left: overhead view; right: lateral view) R pyrenes.
Description.
Procumbent shrubs to ca. 5 cm tall, stems rooting at the nodes; young branchlets puberulent, older branchlets nearly glabrous. Leaf blades broadly elliptic to elliptic, 0.4–1.6 cm long, 0.3–1.3 cm wide, apex rounded to obtuse, base cuneate to rounded, 3-nerved, margin serrate, nearly glabrous, stellate hairs sparsely distributed along the veins on both surfaces; petioles 0.4–1.3(–1.7) mm long, stellate hairs sparse; stipules linear, 0.4–1.2 mm long. Inflorescences terminal, cymose or flowers solitary; peduncles 0.7–2.7 mm long, 1 to 3(4 to 6) flowers, pedicels 0.9–4.9 mm long; bracts linear, 0.7–1.4(–1.8) mm long. Flowers bisexual or functionally unisexual (female), bisexual flowers 7.2–11.7 mm diam., female flowers 5.2–7.8 mm diam. Sepals (3)4 to 5, narrowly oblong to oblong, bisexual flowers 4.5–6.0 mm long, 1.4–2.3 mm wide; female flowers 3.2–4.8 mm long, 0.9–1.9 mm wide, apex acute, abaxial surface yellowish green, stellate–pubescent, adaxial surface white, glabrous. Petals (3)4 to 5, oblong to ovate, apex rounded, bisexual flowers 1.1–1.5 mm long, 0.6–0.9 mm wide, female flowers 1.2–1.6 mm long, 0.7–0.9 mm wide; nectaries present at the base of adaxial surface, 0.6–0.9 mm diam., surrounded by ciliate hairs. Androgynophore cylindrical, bisexual flowers 0.7–0.9 mm long; female flowers 0.8–0.9 mm long, glabrous, ciliate hairs only at the apex. Ovary globose to oblate, pubescent, bisexual flowers 0.8–1.0 mm diam.; female flowers 0.8–1.0 mm diam. Bisexual flowers with stamens 25 to 33, filaments white, glabrous, 1.2–3.2 mm long, anthers dehiscing longitudinally; style (1.8–)2.3–2.7 mm long, glabrous, stigma 4-lobed, each lobe dentate at apex. Female flowers with 15–20 stamens, filaments white, glabrous, 0.6–1.1 mm long, anthers white, always indehiscent; style 1.3–2.2 mm long, glabrous, stigma 4-lobed, each lobe dentate at apex. Fruits drupaceous, usually 4-lobed, fruit lobe 4.1–5.2 mm diam., globose, puberulent to nearly glabrous, red when mature.
Holotype of Grewiakentingensis Y.H.Tseng, Chih Y.Chang & C.Y.Lin, C. Y. Lin et al. 93 (TCF).
Phenology.
Flowering from May to August and fruiting from June to September.
Distribution and habitat.
The species is endemic to Taiwan. Grewiakentingensis is sparsely distributed only on the elevated coral reefs and the coastal open grasslands at 50–200 m alt. along the eastern coastline of the Hengchun Peninsula. Commonly associated species include Maytenusdiversifolia (Maxim.) Ding Hou (Celastraceae), Pandanusodorifer (Forssk.) Kuntze. (Pandanaceae), PhoenixloureiroiKunthvar.loueiroi (Palmae), Rostellulariahayatae (Yamam.) S.S.Ying (Acanthaceae), Galactiatashiroi Maxim. (Fabaceae), and Cirsiumalbescens Kitam. (Compositae).
Chinese name.
kěn-dīng-bǔ-yú-mù (墾丁捕魚木)
Etymology.
The species epithet kentingensis refers to the type locality of Kenting in Hengchun Peninsula, Pingtung County, Taiwan.
Palynology.
Pollens grains are tricolporate, subprolate to prolate, 35.8–41.4 × 24.1–33.4 μm. Exine reticulate, with muri 0.6–0.8 μm wide and lumen 1.0–2.9 μm. Perforations present in the lumen, 0.04–0.24 μm.
Conservation status.
Grewiakentingensis has an extremely limited distribution on the Hengchun Peninsula and is known from 12 collections representing three populations. It has a geographic range in the form of an estimated EOO of 8 km^2^ (adjusted upward from 1.423 km^2^ following IUCN 2024) and an AOO of 8 km^2^. Even though the distribution of the species is located within the areas of Kenting National Park, its habitat is threatened by grazing, wildfires, and invasive plant species (Cuscutacampestris Yunck., Stachytarphetajamaicensis (L.) Vahl, and Bidenspilosavar.radiata (Sch. Bip.) J.A. Schmidt). Given these ongoing threats, we infer a continuing decline in the area, extent and quality of habitat. With respect to the most serious plausible threat of wildfire, the three occurrences represent one location, which falls within the limits for “Critically Endangered” status. Grewiakentingensis is therefore preliminarily assessed as “Critically Endangered” [CR B1ab(iii)+2ab(iii)] in accordance with the IUCN Red List Categories and Criteria (IUCN 2024). The above analysis is based on the specimens cited and georeferenced as shown in Table 4.
Specimens examined.
Taiwan. Pingtung County • Fongchueisha, 28 Jun 1988, T. C. Huang & S. F. Huang 13542 (TAI) • Ibid., 23 Jul 2013, K. C. Chang 4893, 4894 (CHIA) • Ibid., 29 Mar 2015, C. Y. Chang & C. H. Liu 397 (TNM) • Ibid., 3 May 2024, C. Y. Lin et al. 69 (TCF) • Longzaipu, 6 Sep 2008, P. F. Lu 16921 (TAIF) • Ibid., 17 Sep 2008, T. C. Hsu 1743 (TAIF) • Ibid., 23 Jul 2013, C. M. Wang et al. 15470 (TNM) • Ibid., 14 Aug 2013, C. F. Chen 4697 (TAIF) • Ibid., 12 Apr 2014, C. Y. Chang 30 (PPI) • Sheding, 12 Aug 1995, Y. H. Yu 7968 (TAIF) • Sheding Formosan Sika Deer Restoration Area, 15 Jan 2021, C. Y. Chang et al. 3236 (TNM).
Grewia
piscatorum
Taxon classificationPlantaeMalvalesMalvaceae
Hance in Ann. Sci. Nat., Bot., sér. 5, 5. 208. 1866.
BE673D1D-687D-5801-B36D-31B82A816AB9
Type.
China • Fokien province: Lamyet island, 1860, Swinhoe s.n. in Herb. Hance 6527 (lectotype, designated here: BM [barcode BM000795018 as image!]; isolectotype, K [barcode K000686751 as image!]).
Notes.
In the protologue, Hance (1866) cited a Swinhoe collection that he assigned one of his own collection numbers (“Herb. propr., no. 6527”). Hance (1866) did not unequivocally state that he based the name on a single specimen and since there are duplicates of this Hance number in BM (BM000795018) (Fig. 9) and K (K000686751), we designate a lectotype for Grewiapiscatorum. The sheet selected is in Hance’s herbarium (BM), and it is the duplicate with relatively more leaves and flowers.
Lectotype of Grewiapiscatorum Hance, Swinhoe s.n.in Herb. Hance 6527 (BM000795018).
Description.
Erect to ascending shrubs, ca. 1–2 m tall; young branchlets scabrous, older branchlets nearly glabrous. Leaf blades elliptic, obovate, ovate to rhomboid-ovate, 0.7–7.8 cm long, 0.5–5.5 cm wide, apex acute, obtuse to rounded, base cuneate to obtuse, 3-nerved, margin serrulate to biserrulate, nearly glabrous, stellate hairs sparsely distributed along the veins on both surfaces; petioles 1.3–13.4 mm, sparsely stellate hairy; stipules linear, 2.4–5.1 mm long. Inflorescences leaf-opposed or rarely axillary, umbellate; peduncles 3.1–17.2 mm long, 6 to 15 flowers, pedicels 3.5–8.8 mm long; bracts linear, 1.4–2.6(–3.0) mm long. Flowers bisexual, functionally unisexual male or functionally unisexual female; bisexual flowers 14.8–18.5(–20.2) mm diam.; male flowers 14.2–18.8 mm diam.; female flowers 9.4–14.6 mm diam. Sepals (4)5(6), narrowly oblong to oblong, bisexual flowers 7.5–10.8 mm long, 2.4–4.1 mm wide, male flowers 7.3–8.4 mm long, 2.1–3.0 mm wide, female flowers 4.5–5.7 mm long, 1.4–2.0 mm wide, apex acute, abaxial surface yellowish green, stellate–pubescent, adaxial surface white, glabrous. Petals (4)5(6), oblong to ovate, apex rounded, bisexual flowers 2.2–3.3 mm long, 0.9–1.2 mm wide, male flowers 2.1–2.9 mm long, 0.9–1.2 mm wide, female flowers 1.5–2.5 mm long, 0.6–1.0 mm wide; nectaries present at the base of adaxial surface, 0.8–1.3 mm diam., surrounded by ciliate hairs. Androgynophore cylindrical, bisexual flowers 1.1–1.6 mm long; male flowers 1.1–1.5 mm long; female flowers 0.9–1.2 mm long, glabrous, ciliate hairs only at the apex. Ovary globose to oblate, pubescent, bisexual flowers 1.2–1.4 mm diam.; male flowers 1.2–1.5 mm diam.; female flowers 0.9–1.3 mm diam. Bisexual flowers with 103–118 stamens, filaments white, glabrous, 2.4–6.8 mm long, anthers dehiscing longitudinally; style 3.6–5.4 mm long, glabrous, stigma 4-lobed, each lobe dentate at apex. Male flowers with 70–88 stamens, filaments white, glabrous, 2.1–4.7 mm long, anthers dehiscing longitudinally; style 3.1–4.1 mm long, glabrous, stigma 4-lobed, each lobe dentate at apex. Female flowers with 58–102 stamens, filaments white, glabrous, 0.5–1.5 mm long, anthers white, always indehiscent; style 1.8–2.6 mm long, glabrous, stigma 4-lobed, each lobe dentate at apex. Fruits drupaceous, usually 4-lobed, fruit lobe 4.4–6.4 mm diam., globose, puberulent to nearly glabrous, red when mature.
Phenology.
Flowering from May to September and fruiting from June to October.
Distribution and habitat.
Distributed in Fujian Province and Hainan Province in China and in Taiwan on the coastal areas of Kinmen and Lienchiang Counties.
Palynology.
Pollens grains are tricolporate, prolate, 42.8–47.8 × 27.4–31.0 μm. The exine is reticulate, with muri 0.4–0.7 μm wide and lumen size 1.1–4.0 μm. Perforations present in the lumen, 0.04–0.26 μm.
Specimens examined.
China. Fujian Province • Fuzhou City, Tangyu, 24 May 1964, T. H Chen 493 (AU) • Fuzhou City, Yemayu, 31 May 1964, T. H Chen 515 (AU); Fuzhou City, Zhuyu, 17 May 1963, T. H Chen 1561 (AU) • Xiamen City, Nanputuo, 20 Jul 1954, J. C. Lin 3857 (AU). Hainan Province • Dongfang City, seacoast of Dingjiao village, 20 Aug 1936, S. K. Lau 27727, 27730 (IBSC); Sanya City, Xigu island, 3 Aug 2018, H. L. Hou 92038 (AU). Taiwan. Kinmen County • Brave Fortress, 24 Jun 2017, S. W. Chung 12988 (TAIF) • Caicuo Trail, 2 Jun 2019, C. T. Lu et al. 2515 (TNM) • Doumen to Taiwu Mountain, 22 Jul 2007, C. M. Wang 10497 (TNM) • Fengshang, 20 May 2009, J. S. Shiu & Y. L. Hsueh H315 (CHIA) • General’s Spring, 28 May 2023, C. Y. Lin 22 (TCF) • Jiugong Pier, 25 May 2002, C. M. Wang 5611 (TNM) • Ibid., 7 Sep 2021, C. Y. Chang et al. 3539 (TNM)Ibid., 28 May 2023, C. Y. Lin 17 (TCF) • Jiugong Tunnel to shore-line road, 28 May 2023, C. Y. Lin 20 (TCF) • Liaoluo, 22 May 2017, T. C. Hsu 9131 (TAIF) • L36 Fortress, 25 May 2024, C. Y. Lin et al. 85 (TCF) • Mashan Observation Post, 26 Aug 2015, C. Y. Chang et al. 724 (TCF, TNM) • Ibid., 29 May 2023, C. Y. Lin 24 (TCF) • Paichushan to Huotou, 10 Jun 1999, S. T. Chiu 5386 (TNM) • Qilinshan, 25 May 2024, C. Y. Lin et al. 86 (TCF) • Qingyuan Lake, 9 Aug 2002, I. H. Chiang 120 (TAI) • Shuitou Pier, 28 May 2023, C. Y. Lin 21 (TCF) • Taiwu Mountain, 27 Jul 2005, K. C. Chen s.n. (TAIF) • Ibid., 15 Jun 2006, C. H. Chen 7283 (TNM) • Ibid., 4 Jun 2016, S. W. Chung 12534 (TAIF) • Taiwu nursery, 22 Jul 2007, C. M. Wang 10519 (TNM) • Tashan, 14–17 May 2010, F. Y. Lu et al. H2794 (CHIA) • Tzuhu, 25 Jul 2005, K. C. Chen s.n. (TAIF) • Tzuti, 25 Jul 2005, K. C. Chen s.n. (TAIF) • Wuhu Mountain, 13–16 Jul 2009, F. Y. Lu et al. 1015 (CHIA) • Xiyuan beach to Chengkung Fortress, 29 May 2023, C. Y. Lin 23 (TCF) • Zhaishan Tunnel, 25 May 2024, C. Y. Lin 80 (TCF). Lienchiang County • Beihai Tunnel, 19 Jul 2008, C. M. Wang 12180 (TNM) • Chengkungshan, 27 Aug 2007, T. W. Hsu 13020 (TAIE) • Chiukueishan, 27 Aug 2001, Y. L. Huang et al. 816 (TAI) • Daping to lighthouse, 30 May 1994, T. Y. Liu & H. L. Ho 653 (TNM) • Dapu Inscription, 9 Sep 2007, C. M. Wang & C. P. Lu 10690 (TNM) • Fuzheng, 30 Jul 1999, S. H. Su 624 (TAI) • Jinsha, 1 Jul 1999, S. H. Su 352 (TAI) • Dawo Mountain, 16 Sept 2014, S. W. Chung 11782 (TAIF) • Kunqiu Trail, 17 Oct 2017, C. Y. Chang 1473 (TCF, TNM) • Leishan, 29 Aug 2007, T. W. Hsu 13079 (TAIE).
Supplementary Material
XML Treatment for Grewia kentingensis
XML Treatment for Grewia piscatorum
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Barrett RL (2019) Three new species of Corchorus L. and Grewia L. (Sparmanniaceae / Malvaceae subfamily Grewioideae) from northern Australia, an earlier name in Grewia, and recircumscription of Triumfettakenneallyi Halford.Austrobaileya 10(3): 458–472. 10.5962/p.299917 · doi ↗
- 2Bunge AA (1835) Enumeratio plantarum, quas in China boreali collegit. Mémoires présentés à l’Académie Impériale des Sciences de St.-Pétersbourg par divers Savans et lus dans ses Assemblées 2: 75–148. 10.5962/bhl.title.41483 · doi ↗
- 3Burret M (1926) Beiträge zur kenntnis der Tiliaceen.Notizblatt des Botanischen Gartens und Museums zu Berlin-Dahlem 9(88): 592–880. 10.2307/3994327 · doi ↗
- 4Carvalho-Sobrinho J Mota AC Dorr LJ (2024) Eriothecapaganuccii (Bombacoideae, Malvaceae), A new endangered species from montane forests in the Atlantic Forest of Bahia, northeastern Brazil.Phyto Keys 243: 215–230. 10.3897/phytokeys.243.12570838974889 PMC 11224664 · doi ↗ · pubmed ↗
- 5Chang YT (1987) Tiliaceae. In: Fujianica writing group (Eds) Flora Fujianica, Vol.3. Fujian Science Technology Publishing House, Fujian, China, 393–403.
- 6Chang HT (1989) Flora Reipublicae Popularis Sinicae, Vol. 49(1).Science Press, Beijing, 132 pp.
- 7Chang CS Kim H Chang KS (2014) Provisional Checklist of Vascular Plants for the Korea Peninsula Flora (KPF) (Version 1.0).Designpost, Pajo, 660 pp.
- 8Chang CY Chen CH Wang CM (2018) Grewiatiliifolia Vahl (Malvaceae): A newly recorded species of Taiwan.Collection and Research 31: 71–83. 10.6693/CAR.201812_(31).0006 · doi ↗