Four new species of Cryptothecia (Arthoniaceae, Ascomycota) and Myriostigma (Arthoniaceae, Ascomycota) from China, based on morphology and molecular phylogeny

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
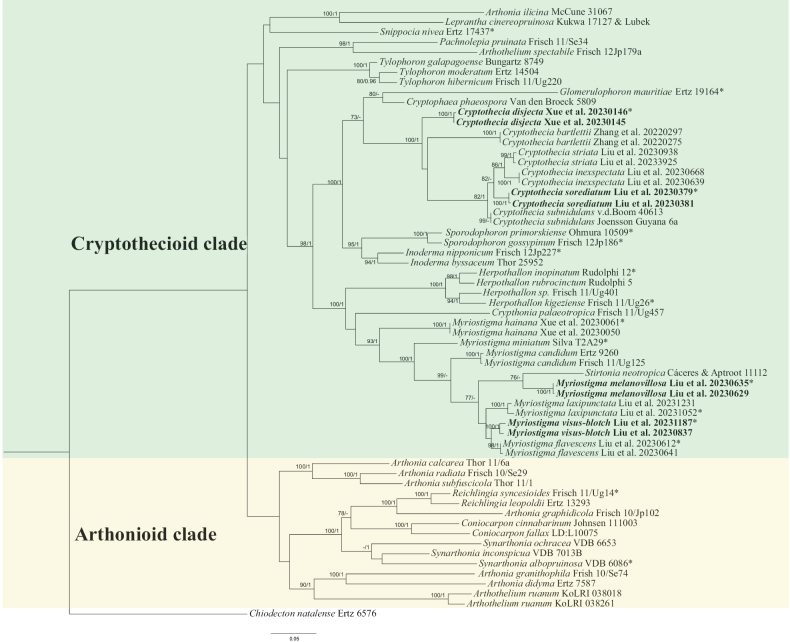 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4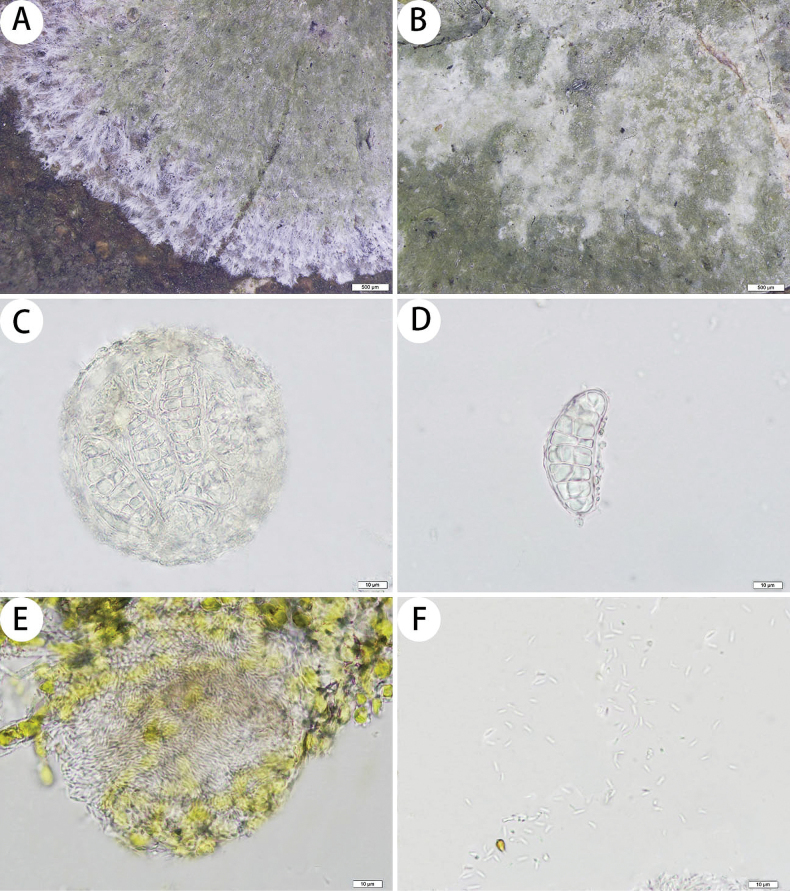 Figure 5
Figure 5| Species Name | Voucher Specimen | GenBank Accession Number | ||
|---|---|---|---|---|
| mtSSU | RPB2 | nuLSU | ||
|
| Thor 11/6a (UPS) |
|
| – |
|
| Ertz 7587 (BR) |
|
|
|
|
| Frish 10/Se74 (UPS) |
|
|
|
|
| Frisch 10/Jp102 (UPS) |
| – |
|
|
| McCune 31067 |
| – | – |
|
| Frisch 10/Se29 (UPS) |
|
| – |
|
| Thor 11/1 (UPS) |
|
| – |
|
| KoLRI 038018 |
|
| – |
|
| KoLRI 038261 |
|
| – |
|
| Frisch 12Jp179a (TNS) |
|
| – |
|
| Ertz 6576 (BR) |
|
|
|
|
| Johnsen 111003 (UPS) |
|
|
|
|
| LD: |
|
| – |
|
| Frisch 11/Ug457 (UPS) |
|
| – |
|
| Van den Broeck 5809 (BR) |
| – | – |
|
| Zhang et al. 20220297 ( |
|
| |
|
| Zhang et al. 20220275 ( |
|
| |
|
|
| – | – | |
|
|
|
| – | – |
|
| Liu et al. 20230668 ( |
|
| |
|
| Liu et al. 20230639 ( |
|
| |
|
| Liu et al. 20230938 ( |
|
| |
|
| Liu et al. 20233925 ( |
|
|
|
|
|
| – | – | |
|
|
|
| – | – |
|
| v.d.Boom 40613 (hd v.d. Boom) |
|
| – |
|
| Joensson Guyana 6a (UPS) |
|
| – |
| Ertz 19164 (BR) |
|
| – | |
| Rudolphi 12 (UPS) |
|
| – | |
| Frisch 11/Ug26 (UPS) |
|
| – | |
|
| Rudolphi 5 (UPS) |
|
| – |
| Frisch 11/Ug401 (UPS) |
|
| – | |
|
| Thor 25952 (UPS) |
|
|
|
| Frisch 12Jp227 (TNS) |
|
| – | |
|
| Kukwa 17127 & Lubek (BR) |
| – | – |
|
| Ertz 9260 (BR) |
|
|
|
|
| Frisch 11/Ug125 (UPS) |
|
| – |
| Liu et al. 20230612 ( |
|
| ||
|
| Liu et al. 20230641 ( |
| ||
| Xue et al. 20230061 ( |
|
| ||
|
| Xue et al. 20230050 ( |
|
| |
| Liu et al. 20231052 ( |
|
|
| |
|
| Liu et al. 20231231 ( |
|
|
|
|
|
|
|
| |
|
|
|
|
| – |
| Silva T2A29 (ISE—epitype) |
| – | – | |
|
|
|
| – | |
|
|
|
|
| – |
|
| Frisch 11/Se34 (UPS) |
|
| – |
|
| Ertz 13293 |
|
|
|
| Frisch 11/Ug14 (UPS) |
|
|
| |
| Ertz 17437 (BR) |
| – | – | |
|
| Cáceres & Aptroot 11112 (ISE) |
| – | – |
| Frisch 12Jp186 (TNS) |
|
| – | |
| Ohmura 10509 (TNS) |
|
| – | |
| VDB 6086 (BR<BEL>) |
| – | – | |
|
| VDB 7013B (BR<BEL>) |
| – | – |
|
| VDB 6653 (BR<BEL>) |
| – | – |
|
| Bungartz 8749 (CDS) |
| – |
|
|
| Frisch 11/Ug220 (UPS) |
|
|
|
|
| Ertz 14504 (BR) |
| – |
|
| 1 | Thallus sterile; without asci |
|
| – | Thallus fertile; with asci |
|
| 2 | Thallus with verrucose pseudisidia; medulla I+ sky-blue |
|
| – | Thallus with globose soralia; medulla I– |
|
| 3 | Asci 1–2-spored |
|
| – | Asci 8-spored |
|
| 4 | Thallus P+ yellow; with psoromic acid |
|
| – | Thallus P–; without psoromic acid |
|
| 5 | Thallus loosely attached to the substrate; ascigerous areas generally covered with globose isidia-like structures; ascospores (49–)68–100(–105) × (18–)23–36(–42) µm |
|
| – | Thallus firmly attached to the substrate; ascigerous areas whitish and usually radially elongated |
|
| 6 | Thallus generally with granula isidia-like structures; ascospores 54–80 × 21–42 µm |
|
| – | Thallus generally without isidia-like structures; ascospores 33–50 × 16–22 µm |
|
| 7 | Thallus without lichen substances |
|
| – | Thallus with lichen substances |
|
| 8 | Ascospores narrow; 60–76 × 17–30 µm |
|
| – | Ascospores broad; 65–108 × 42–50 µm |
|
| 9 | Thallus P+ yellow; with psoromic acid; ascospores 50–70 × 30–37 µm |
|
| – | Thallus P–; without psoromic acid; ascospores 27–40 × 12–20 µm |
|
| 1 |
Ascigerous areas indistinct; thallus with gyrophoric acid and methyl 2’- |
|
| – | Ascigerous areas distinct |
|
| 2 |
Thallus with 2’- |
|
| – | Thallus with gyrophoric and confluentic acids |
|
| 3 | Ascigerous areas without brown dots indicate individual asci; ascospores hyaline (31–)37–74 × (14–)17–29 µm; some specimens have pycnidia |
|
| – | Ascigerous areas with brown dots indicate individual asci |
|
| 4 | Thallus with black or purple dots; ascospores hyaline to pale yellow 63–71 × 26–33 µm |
|
| – | Thallus without black or purple dots |
|
| 5 | Ascigerous areas with dense brown dots; ascospores yellow 58–76 × 19–28 µm |
|
| – | Ascigerous areas with loosely brown dots; ascospores hyaline 57–78 × 24–33 µm |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLichen and fungal ecology · Mycorrhizal Fungi and Plant Interactions · Plant Pathogens and Fungal Diseases
Introduction
Arthoniaceae (Arthoniales, Arthoniomycetes, Ascomycota) is widely distributed in tropical and subtropical regions. Arthoniaceae is characterized by its crustose, sometimes poorly developed or immersed, effuse or determinate thallus; trentepohlioid or rarely chlorococcoid photobiont; not determinate, often elongated and/or branched ascomata with rudimentary walls but rarely thick- and dark-walled; reddish or brownish, I+ blue hymenium; thick walled, ± fissitunicate, usually with a large apical dome and often I+ blue asci; septate and often multi-septate, occasionally muriform ascospores; and small, inconspicuous, coelomycetous (pycnidial) or sporodochial anamorph (Cannon et al. 2020). According to previous studies, the topology of the Arthoniaceae phylogenetic tree is mainly divided into Arthonioid and Cryptothecioid clades (Frisch et al. 2014; Thiyagaraja et al. 2020). Cryptothecia, Herpothallon and Myriostigma belong to the Cryptothecioid clade, and their morphological characteristics are similar, so distinguishing them is difficult. From 2020 to 2023, we collected a large number of lichen specimens from southern China while identifying a large number of species from these specimens. We found three new record species of Cryptothecia, eight new species and five new record species of Herpothallon, and three new species of Myriostigma (Chen et al. 2022; Liu et al. 2023a, 2023b; Xue et al. 2024; Zhang et al. 2024). Recently, we studied the remaining specimens, and we found that several of these specimens differed from the known species in their morphology and molecular phylogeny. Here, we describe two new species of Cryptothecia (C.disjecta and C.sorediatum) and two new species of Myriostigma (M.melanovillosa and M.visus-blotch).
Materials and methods
Morphology and anatomy
The study was based on specimens collected during fieldwork in Hainan and Yunnan Provinces, China, and preserved in the Lichen Section of Botanical Herbarium (SDNU, Shandong Normal University). Morphological features were studied with a dissecting microscope (COIC XTL7045B2), and photos were taken under a microscope (Olympus SZX16, Japan) with a DP72 camera system. The observations included the growth type, diameter and colour of the thallus; the shape and width of the prothallus; and the shape of the ascigerous areas. Anatomical features were observed and measured by a polarizing compound microscope (Olympus CX41, Japan), and images were taken under a microscope (Olympus BX61, Japan) with a DP72 camera system. The observations included the colour of the medulla and whether it contained crystals; the shape and size of the photobiont; the width of the hyphae; and the colour, shape and size of the asci, ascospores, pycnidia and conidia.
Colour reaction and chemical analysis
Colour reactions were performed on the thallus surface and thin medulla sections with a saturated solution of aqueous sodium hypochlorite (C), a 10% aqueous solution of potassium hydroxide (K), a saturated solution of p-phenylenediamine in 95% ethyl alcohol (P), a 3% solution of Lugol’s iodine (I) and long-wavelength UV light. Polarized light microscopy (pol) was used to detect the presence of calcium oxalate crystals. The secondary metabolites of the lichens were analyzed and identified via thin layer chromatography (TLC) with solvent C (Orange et al. 2010; Elix 2014).
DNA extraction, PCR amplification and sequencing
We extracted genomic DNA from the collected specimens using the Sigma-Aldrich REDExtract-N-Amp Plant PCR Kit (St. Louis, MO, USA) following the manufacturer’s protocol, except that only 30 μL of extraction buffer and 30 μL of dilution buffer were used. For extraction, the clean growing portions of the thalli of the specimens were selected under a dissecting microscope (COIC XTL7045B2) with sterile blades and forceps.
We amplified three gene sequences: the mtSSU gene with the primer pairs mtSSU1 and mtSSU3R (Zoller et al. 1999), the RPB2 gene with RPB2-7cF and RPB2-11aR (Liu et al. 1999), and the nuLSU gene with LIC24R and LR7 (Vilgalys and Hester 1990; Miadlikowska and Lutzoni 2000). We performed PCR amplification with a 50 μL volume containing 25 μL of 2 × Taq PCR MasterMix [Taq DNA Polymerase (0.1 unit/μL); 3 mM MgCl_2_; 100 mM KCl; 0.5 mM dNTPs; and 20 mM Tris-HCl (pH 8.3)] (Tiangen, Beijing, China), 19 μL of dd H_2_O, 2 μL of forward primer, 2 μL of reverse primer and 2 μL of DNA. The mtSSU gene was amplified via the following protocol: initial denaturation at 94 °C for 10 min; followed by 34 cycles at 95 °C for 45 s, 50 °C for 45 s, and 72 °C for 90 s; and a final extension at 72 °C for 10 min. The RPB2 gene was amplified via the following protocol: initial denaturation at 94 °C for 10 min; followed by 34 cycles at 94 °C for 45 s, 52 °C for 50 s, and 72 °C for 1 min; and a final extension at 72 °C for 5 min. The nuLSU gene was amplified via the following protocol: initial denaturation at 95 °C for 15 min; followed by 45 cycles at 95 °C for 45 s, 53 °C for 45 s, and 72 °C for 1 min; and a final extension at 72 °C for 7 min. PCR products were sequenced by BioSune Biological Technology (Shanghai, China) using the same primers.
Sequence alignment and phylogenetic analysis
We compared the newly generated sequences with the available sequences in the GenBank database (http://www.ncbi.nlm.nih.gov/BLAST/) to ensure the reliability of all the new sequences. Then, we selected new sequences with high similarity to the clothing species of the Cryptothecia and Myriostigma for further analysis. We assembled the new sequences via SeqMan v.7.0 (DNAstar packages). The sequences of other genera of Arthoniaceae used in this study were downloaded from GenBank (Table 1). We aligned the sequences via the online version of MAFFT v.7.0.26. The algorithm of MAFFT is chosen automatically (FFT-NS-1, FFT-NS-2, FFT-NS-i or L-INS-i; depending on the data size). The sequences were edited via MEGA v.7.0. To construct the phylogenetic tree, the species Chiodectonnatalense Nyl. was selected as the outgroup taxon (Woo et al. 2017).
The multigene phylogenetic trees were inferred via maximum likelihood (ML) and Bayesian inference (BI). The three gene sequences were combined via the Concatenate Sequence function in PhyloSuite v1.2.3 (Zhang et al. 2020). We used the CIPRES Science Gateway (http://www.phylo.org/portal2/) (Miller et al. 2010) and performed ML analyses via RaxML-HPC v. 8.2.12 (Stamatakis 2014) under the default parameters implemented in CIPRES. Support values were based on 1000 nonparametric bootstrap pseudoreplicates. Bootstrap support values for ML equal to or greater than 70 were given above or below the nodes in the phylogenetic tree (Fig. 1). We used PhyloSuite to infer BI phylogenies via MrBayes 3.2.6 (Ronquist et al. 2012) under a partition model, for which the initial 25% of the sampled data were discarded as burn-in. Four Markov chains were run for 2,000,000 generations for the dataset. Trees were sampled every 1000^th^ generations. The stationarity of the analysis was determined by examining the standard deviation of the split frequencies (<0.01). Bayesian posterior probabilities equal to or greater than 0.95 were given above or below the nodes in the phylogenetic tree (Fig. 1). The phylogenetic trees generated were visualized via FigTree v1.4.2 (Rambaut 2012) and edited via Adobe Illustrator (AI). The new sequences were submitted to GenBank (Table 1).
Phylogenetic tree constructed via maximum likelihood (ML) analysis of Arthoniaceae species on the basis of the concatenated mtSSU-RPB2-nuLSU dataset. Bootstrap support values ≥ 70 for ML and posterior probabilities ≥ 0.95 (second value) for Bayesian methods are indicated above or below the branches. Newly obtained sequences are marked in bold, * represents type material.
Results
Phylogenetic analyses
A total of 8 mtSSU sequences, 4 RPB2 sequences and 1 nuLSU sequence were newly generated from 8 specimens. We constructed ML and BI topologies on the basis of these mtSSU, RPB2 and nuLSU sequences and 108 additional sequences downloaded from NCBI (https://www.ncbi.nlm.nih.gov/) (Table 1). The phylogenetic trees obtained from the ML and BI analyses presented similar topologies; therefore, we present only the ML tree, with BS ≥ 70 for the ML analysis and PP ≥ 0.95 for the Bayesian analysis (Fig. 1).
The phylogenetic tree structure we obtained was similar to that described in previous studies (Frisch et al. 2014; Thiyagaraja et al. 2020). All the species positions’ strongly supported the results of the ML and Bayesian analyses. Our phylogenetic tree revealed that C.disjecta and C.sorediatum clustered with C.subnidulans (the type species of Cryptothecia). However, on the basis of differences in phylogeny and morphology compared with those of C.subnidulans, which are described in detail below, they are classified as two new species. M.melanovillosa and M.visus-blotch clustered with Myriostigma and Stirtonianeotropica Aptroot. According to Aptroot (2009), S.neotropica has transversely septate ascospores; thus, on the basis of its morphological characteristics and phylogenetic analysis, we propose two new species in Myriostigma. Therefore, on the basis of morphological characteristics and phylogenetic analysis of the combined mtSSU, RPB2 and nuLSU sequence datasets, there is sufficient evidence to verify four previously unknown new species: Cryptotheciadisjecta J.X. Xue & Lu L. Zhang, sp. nov.; C.sorediatum J.X. Xue & Lu L. Zhang, sp. nov.; Myriostigmamelanovillosa J.X. Xue & Lu L. Zhang, sp. nov.; and M.visus-blotch J.X. Xue & Lu L. Zhang, sp. nov.
Taxonomy
Cryptothecia
disjecta
Taxon classificationFungiArthonialesArthoniaceae
J.X. Xue & Lu L. Zhang sp. nov.
97D8EE49-011D-5432-ACDE-6A13A38A6173
855064
Diagnosis.
The new species is distinguished from other Cryptothecia species by its verrucose pseudisidia, which are loosely scattered on the thallus. The upper parts of the pseudisidia are white or have darker dots.
Type.
China • Hainan Province, Baoting Li and Miao Autonomous County, Qixianling Hot Spring National Forest Park, 18°42'14.43"N, 109°41'47.92"E, alt. 325 m, on the bark of trees, 8 March 2023, J.X. Xue et al. 20230146 (SDNU, holotype).
Description.
Thallus corticolous, up to 15 cm in diameter, ecorticate, cottony, dull, pale green, loosely attached to the substrate. Pseudisidia verrucose, loosely scattered on the thallus, upper parts white or with darker dots, and most pseudisidia have few projecting hyphae, 0.13–0.26 × 0.13–0.22 mm. Prothallus is usually distinct, thin, whitish byssoid, mainly composed of interwoven and radiating hyphae, 1.1–1.5 mm wide. Medulla white, with calcium oxalate crystals. Photobionts trentepohlioid, cells elliptical to oblong, single or aggregate into bundles, 10–23 × 7–18 µm. Hyphae 1–2.5 µm wide.
Cryptotheciadisjecta (SDNU 20230144, type) A thallus and prothallus B pseudisidia. Scale bars: 500 µm (A); 200 µm (B).
Asci and pycnidia were not observed.
Chemistry.
thallus C+ red, K+ pale yellow, P–, UV+ pale grey-white; medulla and paraphysoids I+ sky-blue. TLC: gyrophoric acid and lecanoric acid.
Etymology.
The epithet refers to its pseudisidia, which are verrucose and loosely distributed across the thallus.
Ecology and distribution.
This species is found only in China on the bark of trees in a humid tropical forest in Hainan Province.
Notes.
Morphologically, Cryptotheciadisjecta is similar to both Herpothallon and Cryptothecia. However, we describe C.disjecta on the basis of its morphological and chemical characteristics and assign it to Cryptothecia on the basis of sequencing (Aptroot et al. 2024). Cryptotheciadisjecta is similar to C.eungellae G. Thor, as they both have whitish byssoid prothallus, I+ sky-blue medulla and C+ red thallus, but C.eungellae has 1-spored asci, muriform ascospores, and gyrophoric acid and norstictic acid as secondary metabolites (Thor 1997; Jagadeesh Ram and Sinha 2016).
Phylogenetically, Cryptotheciadisjecta is clustered with C.bartlettii G. Thor. They both have cottony thallus, whitish byssoid prothallus and C+ red thallus, but C.bartlettii has delimited ascigerous areas (developing in the thallus centre and covered with globose isidia-like structures), 1-spored asci and muriform ascospores (Thor 1997).
Additional specimens examined.
China • Hainan Province, Baoting Li and Miao Autonomous County, Qixianling Hot Spring National Forest Park, 18°42'14.43"N, 109°41'47.92"E, alt. 325 m, on the bark of trees, 8 March 2023, J.X. Xue et al. 20230144 (SDNU); • ibid., 20230145 (SDNU).
Cryptothecia
sorediatum
Taxon classificationFungiArthonialesArthoniaceae
J.X. Xue & Lu L. Zhang sp. nov.
73062017-1A15-5128-B1FB-098533134A32
855065
Diagnosis.
The new species differs from other species of Cryptothecia in its soralia and I– medulla.
Type.
China • Yunnan Province, Xishuangbanna Dai Nationality Autonomous Prefecture, Jinghong City, Jinuo Mountain, Jinuo Ethnic Township, 21°54'52.26"N, 101°11'33.04"E, alt. 630 m, on the bark of trees, 3 March 2023, L.L. Liu et al. 20230379 (SDNU, holotype).
Description.
Thallus corticolous, up to 4 cm in diameter, ecorticate, cottony, dull, pale green, firmly attached to the substrate. Soralia globose, with many calcium oxalate crystals, 0.07–0.2 × 0.07–0.2 µm. Prothallus is usually distinct, thick, whitish byssoid, mainly composed of interwoven and radiating hyphae, 0.9–1.7 mm wide. Medulla white, with calcium oxalate crystals. Photobionts trentepohlioid, cells rounded to elliptical, single or aggregate into bundles, 7–12 × 6–12 µm. Hyphae 1–2.5 µm wide.
Cryptotheciasorediatum (SDNU 20230377, type) A thallus and prothallus B soralia. Scale bars: 500 µm (A, B).
Asci and pycnidia were not observed.
Chemistry.
thallus C+ red, K–, P–, UV+ pale grey-white; medulla and paraphysoids I–. TLC: gyrophoric acid and lecanoric acid.
Etymology.
The epithet refers to the presence of soralia on its thallus.
Ecology and distribution.
This species is found only in China on the bark of trees in a humid tropical forest in Yunnan Province.
Notes.
Morphologically, Cryptotheciasorediatum is similar to C.bartlettii G. Thor, as they both have C+ red thallus and globose structures (C.sorediatum has globose soralia, and C.bartlettii has globose isidia-like structures) on their thallus, but C.bartlettii has I+ sky-blue medulla, 1-spored asci and muriform ascospores (Thor 1997).
Phylogenetically, Cryptotheciasorediatum is clustered with C.subnidulans Stirton, C.inexspectata G. Thor and C.striata G. Thor. They all have a cottony thallus and whitish byssoid prothallus, but C.subnidulans has a C– thallus, muriform ascospores (69–90 × 29–44 µm) and psoromic acid as a secondary metabolite (Thor 1997). C.inexspectata has whitish ascigerous areas, I+ sky-blue medulla and muriform ascospores (33–50 × 16–22 µm) (Thor 1997). C.striata has granular isidia-like structures on the thallus, I+ sky-blue medulla and muriform ascospores [(46–)55–70(–80) × (19–)23–29(–37) µm] (Thor 1991).
Additional specimens examined.
China • Yunnan Province, Xishuangbanna Dai Nationality Autonomous Prefecture, Jinghong City, Jinuo Mountain, Jinuo Ethnic Township, 21°54'52.26"N, 101°11'33.04"E, alt. 630 m, on the bark of trees, 3 March 2023, L.L. Liu et al. 20230377 (SDNU); • ibid., 20230381 (SDNU).
Myriostigma
melanovillosa
Taxon classificationFungiArthonialesArthoniaceae
J.X. Xue & Lu L. Zhang sp. nov.
91DCB7B4-BE04-5AE2-B50D-74C823DB5907
855066
Diagnosis.
The new species differs from other species of Myriostigma in the presence of black or purple dots on the thalli and hyaline to pale yellow ascospores (63–71 × 26–33 µm).
Type.
China • Yunnan Province, Xishuangbanna Dai Nationality Autonomous Prefecture, Jinghong City, Primitive Forest Park, 22°1'55.75"N, 100°52'37.47"E, alt. 689 m, on the bark of trees, 7 March 2023, L.L. Liu et al. 20230635 (SDNU, holotype).
Description.
Thallus corticolous, up to 6 cm in diameter, ecorticate, cottony, dull, greenish grey to whitish grey, with black or purple dots, firmly attached to the substrate. Isidia not observed. Prothallus is usually distinct, thin, whitish byssoid, mainly composed of interwoven and radiating hyphae, 0.7–1.3 mm wide, forming a dark brown to black line while bordering different species. Medulla white, with calcium oxalate crystals. Photobionts trentepohlioid, cells rounded to elliptical, single or a few cells aggregated, 5–11 × 5–8 µm. Hyphae 1–2 µm wide.
Myriostigmamelanovillosa (SDNU 20230629, type for (A, B); SDNU 20230635, holotype for (C, D, E, F)) A prothallus B black or purple dots C thallus D ascigerous areas E asci F ascospores. Scale bars: 200 µm (A, B, D); 500 µm (C); 10 µm (E, F).
Ascigerous areas are distinct, generally delimited, erumpent, slightly raised above the thallus level, plaque, irregular in outline, white with dense brown dots indicating individual asci. Asci frequent, hyaline, pale yellow when mature, globose to subglobose, often covered by hyaline hyphae, 8-spored, 103–115 × 97–115 µm. Ascospores hyaline, pale yellow when mature, oblong, muriform, curved, often broader in the centre, 63–71 × 26–33 µm.
Pycnidia were not observed.
Chemistry.
thallus C+ red, K–, P–, UV+ pale grey-white; medulla and paraphysoids I+ sky-blue. TLC: gyrophoric acid, lecanoric acid and confluentic acid.
Etymology.
The epithet refers to the presence of black or purple dots on the thallus.
Ecology and distribution.
This species is found only in China on the bark of trees in a humid tropical forest in Yunnan Province.
Notes.
Morphologically, Myriostigmamelanovillosa is similar to M.irregularis Lücking, Aptroot, Kalb & Elix, as they both have irregular erumpent and with brown dots whitish ascigerous areas, but M.irregularis has narrower asci (40–70 µm wide) and psoromic acid, subpsoromic acid, 2′-O-demethylpsoromic acid and trace confluentic acid as secondary metabolites (Lücking et al. 2006).
Phylogenetically, Myriostigmamelanovillosa is clustered with Stirtonianeotropica Aptroot. They both have C+ red thalli and 8-spored asci, but S.neotropica has linear shape ascigerous areas, ovoid asci and transversely septate ascospores (35–38 × 10–12 µm) (Aptroot 2009).
Additional specimens examined.
China • Yunnan Province, Xishuangbanna Dai Nationality Autonomous Prefecture, Jinghong City, Primitive Forest Park, 22°1'55.75"N, 100°52'37.47"E, alt. 689 m, on the bark of trees, 7 March 2023, L.L. Liu et al. 20230629 (SDNU); • ibid., 20234628 (SDNU).
Myriostigma
visus-blotch
Taxon classificationFungiArthonialesArthoniaceae
J.X. Xue & Lu L. Zhang sp. nov.
300734BC-667D-5854-9AD5-77EDF677CE68
855067
Diagnosis.
The new species differs from other species of Myriostigma in ascigerous areas, which have many irregular small patches that are scattered or clustered together radially elongated; hyaline ascospores (31–)37–74 × (14–)17–29 µm.
Type.
China • Yunnan Province, Xishuangbanna Dai Nationality Autonomous Prefecture, Jinghong City, Primitive Forest Park, 22°2'9.71"N, 100°53'5.81"E, alt. 716 m, on the bark of trees, 7 March 2023, L.L. Liu et al. 20230681 (SDNU, holotype); • ibid., Mengla County, Menglun Town, Xishuangbanna Tropical Botanical Garden, 21°55'12.06"N, 101°16'5.55"E, alt. 496 m, on the bark of trees, 5 March 2023, L.L. Liu et al. 20231187 (SDNU, paratype).
Myriostigmavisus-blotch (SDNU 20230950, type for (A, B); SDNU 20230681, holotype for (C, D); SDNU 20230847, type for (E, F)) A thallus and prothallus B ascigerous areas C asci D ascospores E pycnidia F conidia. Scale bars: 500 µm (A, B); 10 µm (C, D, E, F).
Description.
Thallus corticolous or bambusicolous, up to 6.5 cm in diameter, ecorticate, cottony, dull, pale green to greenish grey, firmly attached to the substrate. Isidia not observed. Prothallus is usually distinct, thin, whitish byssoid, mainly composed of interwoven and radiating hyphae, 0.8–2.5 mm wide, forming a dark brown to black line while bordering different species. Medulla white, with calcium oxalate crystals. Photobionts trentepohlioid, cells elliptical to oblong, single or aggregate into bundles, 6–14 × 6–9 µm. Hyphae 1–2 µm wide.
Ascigerous areas are distinct, generally delimited, slightly raised above the thallus level, developing in the thallus centre, colour lighter than the thallus, many irregular small patches that are scattered or clustered together radially elongated. Asci hyaline, globose to subglobose, often covered by hyaline hyphae, 8-spored, 82–91 × 81–90 µm. Ascospores hyaline, oblong, muriform, curved, often broader in the centre, (31–)37–74 × (14–)17–29 µm.
Pycnidia hyaline, globose to subglobose, scattered and immersed across the thallus, 55–85 × 54–80 μm. Conidia hyaline, short bacilliform, 4–5 × 0.9–1 μm.
Chemistry.
thallus C+ red, K+ pale yellow, P–, UV+ pale grey-white; medulla and paraphysoids I+ sky-blue. TLC: gyrophoric acid, lecanoric acid and confluentic acid.
Etymology.
The epithet refers to its ascigerous areas having many irregular pale green small patches.
Ecology and distribution.
This species is found only in China on bamboo and trees in humid tropical forests in Hainan and Yunnan Provinces.
Notes.
Myriostigmavisus-blotch is widely distributed in the Xishuangbanna Dai Nationality Autonomous Prefecture. The ascigerous areas of M.visus-blotch begin as a small bulge slightly higher than the thallus and gradually become multiple small patches or radially elongated in the centre of the thallus, and the colour of the ascigerous areas is pale green. In addition, we found that species with asci generally do not have pycnidia, and species with pycnidia generally do not have asci.
Phylogenetically, Myriostigmavisus-blotch is clustered with M.flavescens J.X. Xue & Lu L. Zhang and M.laxipunctata J.X. Xue & Lu L. Zhang. They all have C+ red thalli, globose asci and muriform ascospores, but M.flavescens has a black linear shape prothallus, white ascigerous areas and yellow ascospores (58–76 × 19–28 µm). M.laxipunctata has complete ascigerous areas (not small patches or radially elongated), larger asci (95–124 × 93–119 µm), and loose brown dots indicating asci (Xue et al. 2024).
Additional specimens examined.
China • Hainan Province, Baoting Li and Miao Autonomous County, Qixianling Hot Spring National Forest Park, 18°42'14.43"N, 109°41'47.92"E, alt. 325 m, on the bark of trees, 8 March 2023, J.X. Xue et al. 20230142 (SDNU); • ibid., 20230138 (SDNU); • ibid., 20230134 (SDNU); • Yunnan Province, Xishuangbanna Dai Nationality Autonomous Prefecture, Jinghong City, Mengla County, Menglun Town, Xishuangbanna Tropical Botanical Garden, 21°55'45.40"N, 101°15'29.17"E, alt. 518 m, on the bark of trees, 4 March 2023, L.L. Liu et al. 20230950 (SDNU); • ibid., 20230969 (SDNU); • ibid., 21°55'30.14"N, 101°15'42.72"E, alt. 527 m, on the bark of trees, 5 March 2023, L.L. Liu et al. 20233946 (SDNU); • ibid., 21°55'50.99"N, 101°15'33.29"E, alt. 512 m, on the bark of trees, 4 March 2023, L.L. Liu et al. 20230973 (SDNU); • ibid., 21°55'55.96"N, 101°14'35.00"E, alt. 560 m, on the bark of trees, 4 March 2023, L.L. Liu et al. 20230847 (SDNU); • ibid., on bamboo, 4 March 2023, L.L. Liu et al. 20230837 (SDNU); • ibid., Jinuo Mountain, Jinuo Ethnic Township, 21°54'52.26"N, 101°11'33.04"E, alt. 640 m, on the bark of trees, 3 March 2023, L.L. Liu et al. 20230502 (SDNU); • ibid., Wild Elephant Valley, 22°10'37.70"N, 100°51'24.54"E, alt. 749 m, on the bark of trees, 6 March 2023, L.L. Liu et al. 20230602 (SDNU); • ibid., Primitive Forest Park, 22°2'9.71"N, 100°53'5.81"E, alt. 746 m, on the bark of trees, 7 March 2023, L.L. Liu et al. 20230692 (SDNU).
Discussion
Cryptothecia was established in 1877 (Stirton 1877), and Myriostigma was established in 1874 (Krempelhuber 1874). These genera are prevalent in the humid tropical and subtropical forests of southern China, boasting a remarkable species diversity. Nevertheless, their morphological distinctions are scarce, making molecular analyses indispensable for classification and species identification. This study focused on Cryptothecioid lichens in Yunnan and Hainan Provinces of China, employing phylogenetic analyses based on DNA sequence data, and identified four novel species: C.disjecta, C.sorediatum, M.melanovillosa, and M.visus-blotch.
Prior to this study, China had reported eight species of Cryptothecia (C.aleurella, C.aleurocarpa, C.bartlettii, C.inexspectata, C.polymorpha, C.striata, C.subnidulans, and C.subtecta) and four species of Myriostigma (M.candidum, M.flavescens, M.hainana, and M.laxipunctata), as documented by Aptroot and Sipman (2001), Aptroot and Sparrius (2003), Aptroot and Rodrigues (2005), Xue et al. (2024), and Zhang et al. (2024). The two new Cryptothecia species, C.disjecta and C.sorediatum, are readily distinguishable from all other Chinese Cryptothecia species by their sterile thalli featuring verrucose pseudisidia (C.disjecta) or globose soralia (C.sorediatum), along with gyrophoric acid and lecanoric acid as secondary metabolites. Among the other Chinese Cryptothecia species containing gyrophoric acid, namely C.bartlettii, C.inexspectata, and C.striata, all are fertile (Thor 1997; Xue et al. 2024; Zhang et al. 2024). Myriostigmamelanovillosa is further distinguished from all other Chinese Myriostigma species by its thallus with black or purple dots, a whitish byssoid prothallus, and the presence of gyrophoric acid and confluentic acid. It also exhibits white plaque ascigerous areas with dense brown dots indicating individual asci, and hyaline to pale yellow ascospores. Although M.flavescens and M.laxipunctata also contain gyrophoric acid and confluentic acid, M.flavescens can be differentiated by its black line prothallus, smaller asci (95–100 × 83–95 µm), and yellow ascospores. Meanwhile, M.laxipunctata is distinguished by its pale greenish complete ascigerous area with loose brown dots (Xue et al. 2024). M.visus-blotch further distinguishes itself from all other Chinese Myriostigma species by its whitish byssoid prothallus, the presence of gyrophoric acid and confluentic acid, and pale greenish patchy (scattered or clustered together radially elongated) ascigerous areas. Myriostigmahainana also possesses a whitish byssoid prothallus and gyrophoric acid, but it can be differentiated by its indistinct ascigerous area, larger asci (120–138 × 120–135 µm), and the additional presence of methyl 2’-O-methylmicrophyllinate alongside gyrophoric acid (Xue et al. 2024).
In summary, the molecular phylogenetic and morphological results support the identification of the four new species in this study.
Key to the Cryptothecia species occurring in China
**: **
Key to the Myriostigma species occurring in China
**: **
Supplementary Material
XML Treatment for Cryptothecia disjecta
XML Treatment for Cryptothecia sorediatum
XML Treatment for Myriostigma melanovillosa
XML Treatment for Myriostigma visus-blotch
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aptroot A (2009) A revision of the lichen genus Stirtonia.Lichenologist 41(5–6): 1–11. 10.1017/S 0024282909990107 · doi ↗
- 2Aptroot A Rodrigues AF (2005) New lichen records for the Azores, with the report of some tropical species new to Europe. Cryptogamie.Mycologie 26(3): 273–280.
- 3Aptroot A Sipman HJM (2001) New Hong Kong Lichens, Ascomycetes and lichenicolous fungi.The Journal of the Hattori Botanical Laboratory 91: 317–343.
- 4Aptroot A Sparrius LB (2003) New microlichens from Taiwan.Fungal Diversity 14(1): 1–50.
- 5Aptroot ACáceres ME Sdos Santos LA (2024) The taxonomy of sterile Arthoniaceae from Brazil: White crusts on overhanging tropical trees can be named.Lichenologist 56(1): 1–13. 10.1017/S 0024282924000021 · doi ↗
- 6Cannon PF Aptroot A Coppins BJ Orange A Sanderson NA Simkin JA Yahr R (2020) Revisions of British and Irish Lichens.British Lichen Society, London, 48 pp.
- 7Chen PF Liu LL Xie CM Zhang L (2022) Four new species of Herpothallon (Arthoniaceae, Arthoniales, Arthoniomycetes, Ascomycota) from China.Phytotaxa 536(1): 83–91. 10.11646/phytotaxa.536.1.5 · doi ↗
- 8Elix JA (2014) A Catalogue of Standardized Thin-Layer Chromatographic Data and Biosynthetic Relationships for Lichen Substances, 3rd edn. Australian National University, Canberra.