Porphyromonas gingivalis outer membrane vesicles alter cortical neurons and Tau phosphorylation in the embryonic mouse brain
Adrienne J. Bradley, Lauren Mashburn-Warren, Lexie C. Blalock, Francesca Scarpetti, Christian L. Lauber

TL;DR
This study shows that Porphyromonas gingivalis outer membrane vesicles can affect the developing mouse brain, reducing brain weight and altering neurons and Tau proteins.
Contribution
The study reveals novel effects of Pg-OMV on embryonic brain development and Tau phosphorylation in mice.
Findings
Pg-OMV exposure significantly decreased pup brain weight without affecting dam or pup body weights.
Pg-OMV increased microglia activation (Iba-1) and reduced cortical neuron density (Cux1, SatB2).
Pg-OMV exposure increased phosphorylated Tau at Thr231 in embryonic mouse brains.
Abstract
Porphyromonas gingivalis (Pg) is an oral bacterial pathogen that has been associated with systemic inflammation and adverse pregnancy outcomes such as low birth weight and pre-term birth. Pg drives these sequelae through virulence factors decorating the outer membrane that are present on non-replicative outer membrane vesicles (OMV) that are suspected to be transmitted systemically. Given that Pg abundance can increase during pregnancy, it is not well known whether Pg-OMV can have deleterious effects on the brain of the developing fetus. We tested this possibility by treating pregnant C57/Bl6 mice with PBS (control) and OMV from ATCC 33277 by tail vein injection every other day from gestational age 3 to 17. At gestational age 18.5, we measured dam and pup weights and collected pup brains to quantify changes in inflammation, cortical neuron density, and Tau phosphorylated at Thr231. Dam…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
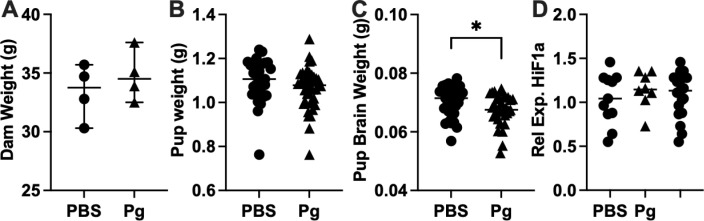 Fig 1
Fig 1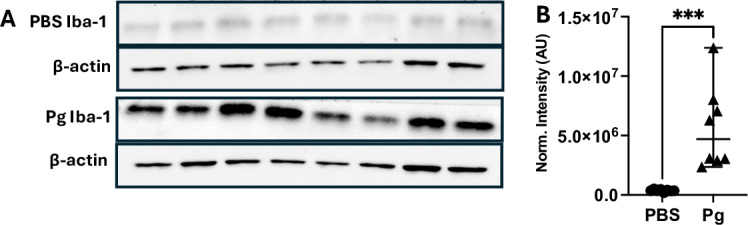 Fig 2
Fig 2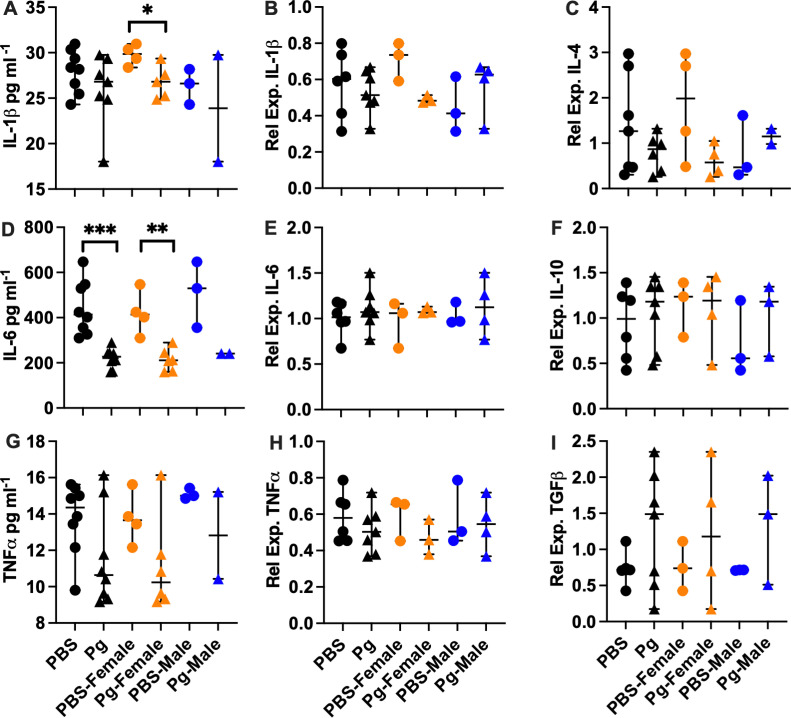 Fig 3
Fig 3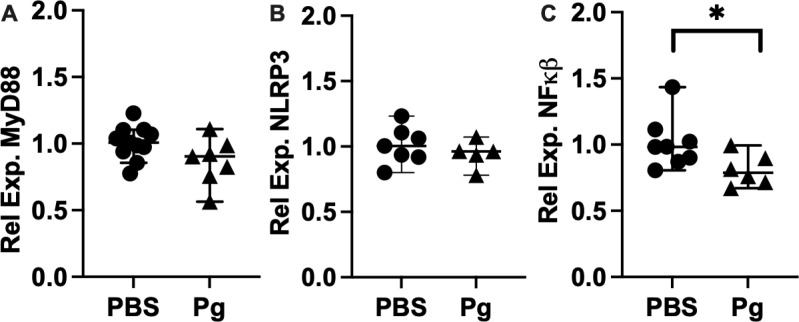 Fig 4
Fig 4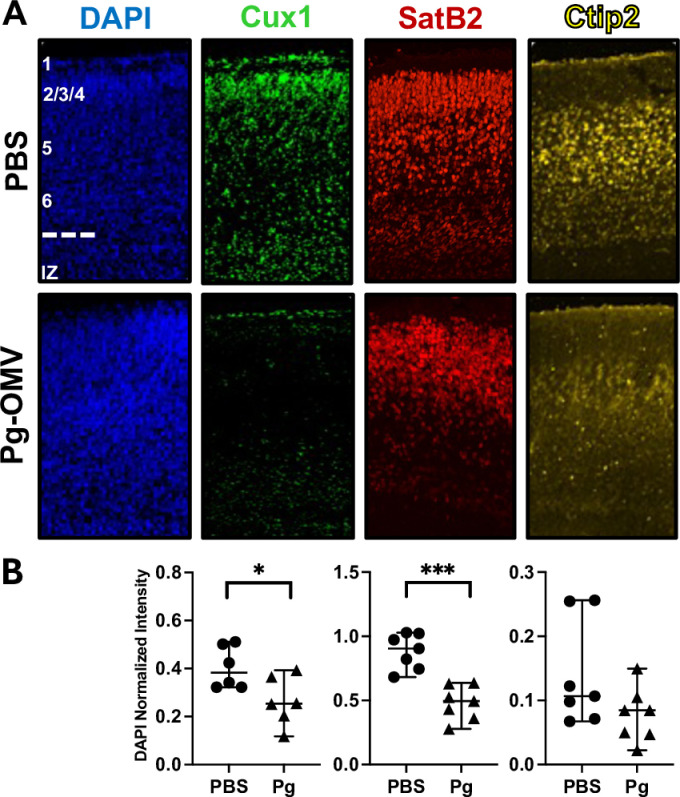 Fig 5
Fig 5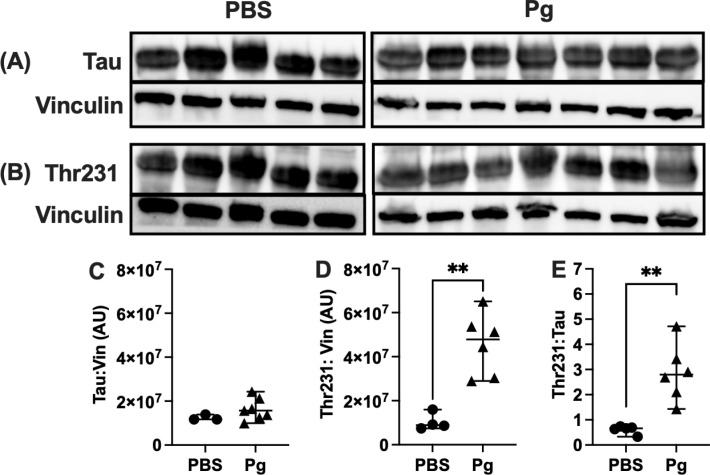 Fig 6
Fig 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial Infections and Vaccines · Oral microbiology and periodontitis research · Neonatal and Maternal Infections
Introduction
Pregnancy induces significant changes in mothers including shifts in the abundance of oral bacteria [1–4]. Increased blood flow and pregnancy related hormones allows for the growth of pathogens like Porphyromonas gingivalis (Pg) an organism associated with oral infections that are increasingly linked to adverse pregnancy and neurodevelopmental outcomes [5–7]. Increased Pg presence is estimated to be prevalent in 40% of pregnancies in the US and 2–11% worldwide [8,9]. This presence of bacteria sometimes resulting in periodontitis, a significant dental infection, can drive systemic inflammation [10–12]. Preterm birth, preeclampsia, and low birth weight are well-characterized effects of maternal Pg infections and Pg DNA has been detected in the placenta and amnionic fluid. Pg driven outcomes affecting children have focused on pregnancy outcomes [13–18] and only recently has Pg received attention as a modulator of fetal development and remains an understudied host-pathogen interaction [19,20].
P. gingivalis has been extensively studied to determine its role in modifying pregnancy. Rodent models of Pg infection demonstrate that exposure to Pg can result in systemic inflammation, low birth weight and pre-term delivery [21–25]. Pg mediates these and other effects thorough a number of virulence factors that modify host responses including lipopolysaccharide (LPS) and proteases known as gingipains [26–32]. Both are also present on the surface of OMV that are now suspected to be transmitted systemically in the blood stream [20,32,33]. LPS and the gingipains interact with the immune system but are unique compared to other gram-negative bacteria. For instance, Pg LPS is less immunogenic than Escherichia coli LPS but also causes inflammation and the production of pro-inflammatory cytokines [34,35]. The gingipains are unique to Pg and they modify cytokine expression that allows Pg to evade the host immune response and establish periodontal infections that drive systemic inflammation [36–39]. Pg infection also modifies the placenta to cause spiral artery remodeling that can result in a hypoxic state linked to preeclampsia and fetal growth restriction [40]. However, whether Pg-OMV may induce fetal growth restriction—a common outcome in animal models and humans with Pg infections of clinical significance—is not yet known.
P. gingivalis is associated with maternal inflammation and adverse pregnancy outcomes that increase the risk of modified developmental trajectories of offspring [17,41]. Maternal infection and the resulting inflammation affect development though mechanisms not fully described at this time [42]. Live Pg is well demonstrated to increase inflammation in animal models and exposure to Pg-OMV recapitulates these observations as the vesicles harbor the same virulence factors. Ishida et al. reported increased Iba-1 indicative of microglia activation that was accompanied by increased inflammation and cytokine production and modified neurons in E 20 pup brains from dams with an established Pg infection prior to pregnancy [19]. Offspring from Pg infected dams also demonstrated declines in cognitive ability, and it was suggested the combined effects on the embryonic brain resulted in neuroinflammation that drove these changes. Similarly, Gong et al. described a neuroinflammatory phenotype in adult mice where exposure to Pg-OMV for 3 months increased IL-1β and NLRP3 expression concurrent with modification of the cortical neurons and behavior [33]. Together these studies suggest that long-term exposure to Pg and Pg-OMV can result in immune phenotypes indicative of a pro-inflammatory response that would be expected from activated microglia in the M1 activation state [43,44]. However, the maternal immune system functions differently than the non-pregnant state and changes in inflammation and the abundance of cortical neurons related to pathogen exposure in utero after pregnancy is established may also differ. This knowledge gap limits our efforts to refine predictions of the potential impact of pathogen-driven inflammation on the developmental trajectory of offspring.
Tau is an essential protein with multiple roles in maintaining neuron homeostasis [45–47]. Proper function is dependent on phosphorylation that can occur at multiple sites and can be modified by inflammation and exposure to pathogens such as Pg [48–50]. Phosphorylated Tau (p-Tau) has less affinity for tubulins and self-aggregates into β-amyloid plaques that are a hallmark of neuroinflammatory diseases such as Alzheimer’s and Parkinson’s diseases in older adults [51]. The resulting Tau tangles can trigger an immune response that is sustained through microglial NFκβ signaling and thus are considered pathogenic forms of the protein [52,53]. In a mouse model, Gong et al. showed that Pg-OMV alter the Tau profile and could result in β-amyloid plaques accompanied by increased inflammation [33]. However, relatively less is known about the role of p-Tau in the developing brain and whether exposure to pathogens during pregnancy alters this profile. A survey of fetal and adult Tau phosphorylation in humans demonstrated significant overlap in p-Tau where some of the pathogenic forms of Tau in adults with Alzheimer’s disease were also prevalent in the fetal brain suggesting p-Tau in children does not have a pathogenic effect as observed in adults [54]. Interestingly, Tau is hyperphosphorylated in the embryonic rodent brain but knock-down of Tau during gestation in mice can reduce neuronal migration to all cortical layers indicating that p-Tau is an important part of normal brain development in mice [48,50,55]. Whether or not Pg-OMV may modulate Tau phosphorylation in developing brains in utero is an open question. Addressing this knowledge gap may provide valuable insight into the mechanisms by which Pg is able to modify developmental trajectories of children.
Herein, we describe a study to determine whether maternal exposure to Pg-OMV could effect the developing mouse brain to understand this host-pathogen interaction that frequently occurs in the human population. To simulate a maternal infection with Pg, pregnant C57/Bl6 mice were exposed to a consistent dose of Pg-OMV via tail vein injection. At gestational age (GA) 18.5, we measured microglia activation, pro- and anti-inflammatory cytokines, total Tau and p-Tau Thr231, and the abundance of cortical neuron proteins in embryonic pup brains. We hypothesize that maternal exposure to Pg-OMV will garner an inflammatory response where Iba-1 expression is accompanied by increased production of pro-inflammatory cytokines that have been observed in humans and animal models attempting to define the impact of Pg infections on the adult and developing brain. Moreover, we expect that Pg-OMV will change the Tau profile and modify expression of cortical neuron proteins concordat with other observations that may be relevant in defining the impact of pathogen exposure on development during pregnancy.
Results
OMV isolation and characterization
OMV from P. gingivalis strain ATCC 33277 were collected from cleared supernatants by ultracentrifugation and stored at − 20 C in PBS. The OMV prep contained spherical particles (S1 1A Fig) that retained gingipain activity and were comprised of LPS at 0.5 EU/ml + /- 0.09 EU (S1 1B and 1C Fig). Plots of activities and concentrations can be found in the supporting information S1.
Pup body and brain weights
Previous studies in mice have demonstrated that dams with Pg infection or those consistently dosed with Pg bacteria during pregnancy can result in low birth weight of pups. We recorded weight of the dams, pups, and pup brains at GA 18.5 (Fig 1A) to determine whether exposure to Pg-OMV would have the same effect. Dam and pup weights were not significantly different in PBS and Pg-OMV groups (Fig 1A and 1B) but there was a significant decrease in the pup brain weight after 14 days of Pg-OMV treatment (Fig 1C, Mann-Whitney U p = 0.023). We included an assessment of Hif1a as Pg is known to cause hypoxia in mouse models that is related to fetal growth restriction. Relative expression of HiF1a in the embryonic mouse brain was not significantly different between the control and treated groups indicating the OMV did not induce a hypoxic state in utero that could account for the decreased pup brain weights (Fig 1D).
Dam and pup body and brain weights.(A) and (B) Weight of dams and pups were not significantly different from PBS controls. (C) Pup brain weight decreased significantly with Pg exposure (Mann-Whitney U p = 0.023). (D) Relative expression of Hif1a as indicator of hypoxia in the pup brain. Relative expression calculated by the 2^-(ΔΔCt) method with GAPDH as the reference gene. Data are shown as median with 95% CI.
Inflammation
Maternal inflammation can affect the developing embryo in mouse models, and we tested whether exposure to Pg-OMV during pregnancy would result in an inflammatory response concordant with increased presence of pathogens experienced during pregnancy. In a similar approach used by Ishida et al [19], we assessed the concentration of Iba-1 as a marker of microglia activation indicative of maternal Pg infection that can lead to increased cytokine production, inflammation, and damage to neurons and may be activated by maternal factors [42,56]. Western blotting of whole brain homogenates showed that Iba-1 was significantly increased in Pg-OMV pup brains compared to PBS controls (Fig 2A and 2B, Mann-Whitney U p = 0.0003). Next, we tested whether increased Iba-1 was accompanied by increased concentration and expression of pro-inflammatory cytokines IL-1β, IL-6 and TNFα (Fig 3) indicative of M1 activation that produces a pro-inflammatory response. ELISA quantification showed that PBS pups typically had higher levels of cytokines but only IL-6 was significantly greater in the PBS brains compared to pups from the Pg-OMV group (Fig 3D, Student’s T-test p = 0.002). The levels of IL-1β and TNFα showed similar trends but were not statistically different between the treatment groups (Fig 3A and 3G, p > 0.05 in both cases). We extended this analysis to include effects that may be apparent at the level of expression in the embryonic brain tissue. RT-qPCR analysis of these cytokines showed no significant difference between the PBS and Pg-OMV pups (Fig 3B, 3E and 3H). However, microglia are pleiotropic and can exist in multiple states when active. As Iba-1 is a general marker of microglia activation, it can not be used to differentiate microglia in the M1 state from those in the so-called M2 state where the production of anti-inflammatory cytokines may be observed [56,57]. Thus, we tested whether there was an anti-inflammatory immune response given that microglia can be in two broad functional categories when activated [43,56,58]. Analysis of IL-4, IL-10 and TGFβ typical of M2 activated microglia were not significantly different between the treatment groups (Fig 3C, 3F, 3I, p > 0.05 in all cases). Lastly, given these results we tested whether the expression of NLRP3, MyD88 and NFκβ, that can be modified by exposure to Pg or it’s OMV, were also modified in our pups. RT-qPCR analysis of MyD88 and NLRP3 transcripts (Fig 4) had similar downward trends with Pg-OMV treatment but were not significantly different compared to controls (Fig 4A, and 4B, p > 0.05 in both cases). Interestingly, NFkβ transcripts that are typically reported as being upregulated by Pg were significantly decreased in the Pg-OMV group (Fig 4C, Student’s T-test p = 0.004). Full Western blots for Iba-1 can be found in the supporting information S2.
Increased Iba-1 in mouse pups exposed to Pg-OMV during pregnancy.(A) Western blot of Iba-1 and β-actin in GA 18.5 pups from PBS and Pg-OMV exposed dams. Full blot can be found in the Supporting information. (B) Comparison of β-actin normalized signal intensity of Iba-1 showed significant increase in Iba-1 in the Pg-OMV group compared to controls (Mann-Whitney U p = 0.0003). Data are shown as median with 95% CI.
ELISA and RT-qPCR evaluation of cytokines in the embryonic mouse brain.(A, D, G): ELISA of IL-1β, IL-6, and TNFα in pg ml-1. Cytokines were extracted from frozen whole brains and quantified. IL-1β was significantly greater in the PBS female pups compared to the female pups from the Pg-OMV group (Fig 3A, Mann-Whitney U test p = 0.04). IL-6 was significantly greater in the PBS group and in the female pups compared to the OMV treatment group (Fig 3D, Student’s T-test p = 0.002 and 0.0095, respectively.) (B, E, H): RT-qPCR of proinflammatory cytokines IL-1β, IL-6, and TNFα. (C, F, I): RT-qPCR of anti-inflammatory cytokines IL-4, IL-10, and TGFβ. Total RNA was extracted from frozen whole brains and reverse transcribed to cDNA. Relative expression calculated by the 2^-(ΔΔCt) method with GAPDH as the reference gene. Group differences for the ELISAs on the males was not determined due to low sample size (n = 2). Only significant differences are shown. Data are presented with the median and 95% CI. Symbols: Circles = PBS, triangles = Pg; orange = females; blue = males.
RT-qPCR evaluation of MYD88, NLRP3, and NFκβ in GA 18.5 pup brains.(A) MyD88 and (B) NLRP3 were not significantly different in pup brains from the control or treatment group. (C) NFkβ was significantly reduced in the pup brains (Student’s T-test p = 0.004). Total RNA was extracted from frozen whole brains and reverse transcribed to cDNA. Relative expression calculated by the 2^-(ΔΔCt) method with GAPDH as the reference gene. Means across fore, mid, and hind brain sections are shown for each sample. Data are presented with the median value and 95% CI.
We were interested in identifying potential modifications in cytokine concentration and expression that may differ between female and male offspring as maternal inflammation may have sex specific effects. When parsed by sex, we observed that IL-6 and IL-1β concentrations were significantly lower in female pups from the Pg-OMV group compared to the female controls (Fig 3A, Mann-Whitney U test p = 0.04 and 3D, Student’s T-test p = 0.0095) and though TNFα was lower in Pg-female pups, it was not statistically different in our analysis (Fig 3A, 3D. 3G). The effect of Pg-OMV on transcription was highly variable for IL-1β, TNFα, IL-4, IL-10 and TGFβ but generally trended upwards (Fig 3B–3I) for the female pups. Trends in the males were similar but were not significant and likely result from low sample sizes (Fig 3), though we could not assess difference in cytokine concentrations as the Pg male group had only 2 observations (Fig 3A, 3D, 3G).
Cortical neurons
Without a strong inflammatory phenotype, we tested the possibility that Pg-OMV treatment could nonetheless result in differences in the cortical neurons of the developing mouse brain. We examined the distribution of neuronal proteins Cux1, SatB2, and Ctip2 that are differentially expressed across the layers of the cortex in sagittal sections of embryonic mouse brains. The distribution and density of these proteins were modified by exposure to Pg-OMV compared to pups from the PBS group (Fig 5A and 5B). We saw significant decreases in Cux1 (Student’s T-test p = 0.03) and SatB2 (Students T-test p = 0.0001) signal intensity but not Ctip2 (Student’s T-test p = 0.064) although it also followed a similar downward trend in the Pg-OMV pup brains (Fig 5B). These findings suggests that cortical layering was drastically altered in Pg exposed animals.
Modified intensity and location of cortical layer marker proteins following Pg-OMV exposure in utero.(A) Representative immunofluorescent images from sagittal sections of PFA fixed brains at GA 18.5 pup brains showing location of cortical layer markers Cux1, SatB2, and Ctip2. Approximate position of cortical layers 1–6 with the intraventricular zone (IZ) are shown in the PBS-DAPI image. (B) Quantification and comparison of cortical layer proteins stained with the marker antibodies. Intensities were averaged across three serial section per sample for statistical analysis. Cux1 (Student’s T-test p = 0.03) and SatB2 (students T-test p = 0.0001) were significantly reduced in the Pg-OMV group. Data are presented with the median and 95% CI.
Total Tau and p-TauThr231
We evaluated expression of total Tau and p-Tau Thr231 as potential markers of shifts in the embryonic Tau pool in response to maternal exposure to Pg-OMV. Western blot and probing with the anti-tau Tau5 antibody indicated a slight but not significant decrease in the mean total Tau concentration in brains from the Pg-OMV group compared to the PBS controls (Fig 6C, Student’s t-test, p = 0.1761).
Differences in total Tau and p-Tau Thr231 following maternal Pg-OMV exposure.(A-B) Representative images of WB showing total Tau and p-Tau Thr231 Tau from E18.5 pup brains. (C-D) Quantification of western blots showing decrease in total Tau and increase in p-Tau Thr231 (Student’s T-Test p = 0.0013). (E) Ratio of p-Tau to Total Tau was significantly increased in the Pg pups (Mann-Whitney U Test p = 0.0025). Data show mean + /- SEM.
Probing with the anti-pTau Thr231 antibody demonstrated Tau was in a phosphorylated state concordant with other observations of Tau in the developing mouse brain (Fig 6A). Normalizing the p-Tau Thr231 against vinculin and total Tau showed that p-Tau Thr231 was significantly increased in the brains of embryos whose mothers were dosed with OMVs compared to the embryos of unexposed mothers (Fig 6D, Student’s T-Test p = 0.0013, 6E Mann-Whitney U Test p = 0.0025).
Discussion
Animal models of Pg infection demonstrate modified pregnancy outcomes and can alter birth weight of offspring. Our experimental design did not recapitulate these observations as pup weights in the Pg-OMV group were not statistically different from PBS controls. Moreover, dam weights were also unchanged indicating that food and water intake were likely not different between the control and treatment groups. The mechanism behind reduced brain weight is not clear from our observations, although pup weight followed a similar downward trend suggesting that the effect of Pg-OMV was systemic for the developing embryos. It could be possible that exposure to pathogens after initiation of pregnancy is different from a state where the pathogen is already present and interacting with host defense mechanisms. Likewise, the dosage of Pg-OMV we used may have not been sufficient to recapitulate the full effect of Pg on pregnancy outcomes, where more robust responses of the host and fetus may be required to lower birth weights. This would coincide with unchanged expression of Hif1a. Nevertheless, treatment with a low dosage of Pg-OMV during pregnancy impacted the development of the embryonic brain that could have implications after birth.
P. gingivalis and Pg-OMV iinduced neuroinflammation is suspected to be involved in cognitive decline in aging cohorts and have a significant impact on development in utero. Our hypothesis that maternal exposure to Pg-OMV would produce a pro-inflammatory response in the embryonic mouse brain was not supported. Cytokine concentrations and levels of mRNA transcripts in the pup brains tended to be lower in the Pg-OMV group. The lack of concordance with previous work was somewhat unexpected as most reports demonstrate an inflammatory response upon exposure to Pg or its OMV. Significant increases of Iba-1 the embryonic brain of Pg-OMV exposed dams could indicate sensing of bacterial-derived components, such as LPS, that can stimulate immune response, though this was not accompanied by a significant increase in cytokine production in our mice. Moreover, Pg has atypical LPS compared to organisms like E. coli LPS, that can bind to TLR-4, but can be recognized by alternative sensing pathways. We addressed this possibility by quantifying the expression of MyD88 and NLRP3 inflammasome mRNAs that are known to increase upon exposure to Pg or Pg-OMV. Our analysis found that concentrations of both were unaffected by OMV treatment in GA 18.5 pup brains. These relationships suggest direct interaction of Pg, OMV, or circulating virulence factors could be responsible for the observed phenotypes. However, microglia could be activated indirectly by Pg or Pg-OMV through maternal signals not quantified in this study. In either case there needs to be more work to clarify direct and indirect fetal-pathogen interactions that modify development. We suspect several factors may be contributing to our observations. First, our results could originate from the delivery of Pg-OMV via tail vein injection which bypass sensing mechanisms such as dendritic cells of the gastrointestinal tract that would detect Pg and LPS being translocated from the oral cavity in saliva. Second, it is equally possible that Pg-OMV induced inflammation reported elsewhere may be an outcome in adult mice with long-term exposure to Pg or Pg-OMV but may not necessarily be an outcome for exposure after pregnancy has been established given the maternal immune system functions differently compared to the pre-pregnancy state. Thus, our observations may not be comparable to reports that use Pg infection or treatment with Pg or Pg-OMV at high doses over several weeks prior to inducing pregnancy. Importantly, the dosing scheme described here suggests there is a potential use of Pg-OMV as a proxy for bacteria to study effects on fetal tissues independent of systemic inflammation driven by common oral pathogens that become more prevalent during pregnancy. More work is needed to elucidate how the timing and dosage of pathogen exposure relative to the start of pregnancy modulates immune responses during gestation that could be associated with long-term consequences for offspring in human populations.
Several reports have quantified the impact of Pg and Pg-OMV on neurons as that cell type is affected by inflammation and is important in brain development. Our data indicate that Pg-OMV have a significant impact on neurons as their prevalence was modified in the cortical layers of the embryonic brain. Cux1, SatB2, and Ctip2 signals were lower overall and showed modified distribution in the Pg-OMV exposed brains compared to controls. The down regulation of SatB2 and Cux1 are particularly interesting as abnormal expression of both are associated with altered behavior in mice and are suspected to be a contributor to autism spectrum disorder-like symptoms in humans. Likewise, deficiencies in Cux1 expression have been linked to neurodevelopmental delays, while down regulation of Ctip2 is known to affect axonal growth and is involved in spiny neuron differentiation. The changes observed herein are aligned with previous studies, yet novel because these cortical layer aberrations were not accompanied by any indication of inflammation. Collectively, these data suggest that Pg, or perhaps Pg-OMV specially, affect the developing brain through a different mechanism. Parsing out the effects of Pg-OMV on cell proliferation and migration appears to be critical in deciphering how Pg mediates effects on the developing brain and should be addressed in future studies to better understand this important host-pathogen interaction.
The importance of Tau in the developing brain has not been fully described but our observations indicate maternal exposure to Pg-OMV can alter the amount of p-Tau Thr231 in mouse embryos. Tauopathies have multiple effects on the central nervous system including sustained inflammation through NFκβ signaling microglia that detects the pathogenic form of Tau in adult animal models. Our data would appear to support the notion that p-Tau Thr231 in the embryonic mouse brain is not pathogenic as an innate immune response, concordant with an inflammation-based hypothesis of neurodegenerative phenotypes, was not observed. Accordingly, we did not observe an increase in NFκβ that can encourage transcription of cytokines. We did not perform a comprehensive assessment of Tau phosphorylation sites, nor did we specifically address potential interactions between p-Tau and microglia. Nonetheless, the alteration of Thr231 abundance aids in explaining how changes in neuron migration and axon growth may result from exposure to Pg in utero; our observed changes in cortical layer markers could be the result of altered microtubule binding driven by altered p-Tau. Thus, much work remains to be done in elucidating the role of Pg and Pg-OMV on human neurodevelopment and cognitive trajectories from the early to elderly phases of life.
Methods
OMV isolation and characterization
Porphyromonas gingivalis strain ATCC 33277 was grown in anaerobic conditions (85% N_2_, 10% CO_2_, 5% H_2_) using Tryptic Soy Broth (Difco cat# 211768) medium supplemented with 5.0 mg ml-1 hemin (ThermoFisher, cat# AAA1116503), 1.0 mg ml^-1^ menadione (MP Biomedicals, cat # 102259), and 0.5 g L^-1^ L-cysteine-HC (MP Biomedicals, cat # 101446). A cleared supernatant was produced by pelleting cells at 8000 rpm for 15 minutes followed by filtration of the supernatant through a 0.45 um membrane filter under low vacuum pressure. Using Sorvall 11.5 mL ultracentrifuge tubes and ultracrimp tube plugs, filtered supernatant was ultacentrifuged at 65,000 rpm for 1 hour at 4°C using a Sorvall WX Ultra 80 centrifuge. Immediately after centrifugation, supernatant was discarded and the pellet containing the OMV was resuspended in 500 uL of PBS [59]. The gingipain activity was quantified using the assay as described by Potempa and Nguyen using L-BAPNA (Millipore-Sigma cat# B3279) as the substrate and live Pg as positive control [60]. LPS was quantified by using the Pierce Chromogenic Endotoxin Quant Kit (ThermoFisher cat # A39552) as directed by the manufacturer. Assays were conducted in clear 96-well flat bottom plates and the absorbance read on a Synergy H1 multimode plate reader (BioTek). Transmission electron microscopy of OMV was performed by placing 10 ul of Pg OMV prep on a formvar coated copper grid for 1 minute. The liquid was then wicked off and UranyLess electron microscopy stain (Electron Microscopy Sciences, Hatfield, PA) was added directly to the grid for 15 seconds and then wicked off. Grids were dried for 10 minutes and imaged with a Hitachi H-7650 Transmission Electron Microscope transmission electron microscope. The total protein of the OMV were quantified using the Pierce BCA (ThermoFisher cat # 23227) assay with BSA as a standard. OMV preps were stored at -20°C for future use.
Animal breeding and care, dosing and tissue collection
2–3 month old C57/Bl6 mice (male and female) were purchased from Jackson Labs (Maine, USA). All animals were housed in the animal research core under 12 h light/dark cycle. Mice were bred and dams were checked for plugs 2 x /day. Once a plug was visible, the dam was removed and was marked as gestational day 0 (GA 0). Starting at E3, 50 µg Pg OMV were administered via tail vein injection every other day until GA 18.5. An equal volume of sterile PBS was used for the control group. Pregnant dams were anesthetized using isoflurane and pups were removed via C-sections (care and euthanization details in the following section). Pups were decapitated and whole brain and placenta were collected from each pup. Half of the pup brains were randomly chosen and fixed using 4% paraformaldehyde followed by 30% sucrose prior to being mounted in O.C.T. and frozen for Immunohistochemical and Immunofluorescent analysis. The remaining brains were snap frozen in a liquid N_2_ at time of collection and used for protein, RNA, and DNA extraction. Fixed tissues were stored at 4°C and frozen tissues were stored at -80°C.
Humane endpoints were used to ensure the ethical treatment of animals and minimize suffering. Briefly, pregnant mice were dosed with OMVs every other day from GA 3 to GA17. Lab staff were trained in euthanasia techniques and tail vein injections but staff members from the Animal Resource Center at the Research Institute at Nationwide Children’s Hospital administered the OMV via tail vein injection for this experiment. A total of 20 mice, 4 males and 16 females, were used for this with tissues coming from 8 dams. Pregnant mice were anesthetized at GA 18.5 using isoflurane for the duration of the C-section. Once all pups had been removed, euthanasia proceeded by transcardiac perfusion of a saline solution followed by ice cold 4% paraformaldehyde fixative with subsequent decapitation. Indications that mice were in distress were monitored daily and included: decreased activity levels, lethargy, pallor, hunched posture, decreased grooming and rough fur, decreased fecal output, and a positive skin tent test for dehydration. Signs of dehydration would be treated with subcutaneous fluid replacement that if not resolved within 48 hours of the first observation would constitute meeting an endpoint for euthanization. Body weights for each mouse recorded every 48 hours and a decrease of 20% or greater mass over this time met endpoint criteria for euthanization. Any dams that met endpoint criteria prior to GA 18, or males that met criteria at any time, were euthanized by CO_2_ asphyxiation and cervical dislocation on the day the animal met endpoint criteria. Fortunately, no animals met endpoint criteria prior to GA 18.5 and no deaths were recorded for the 18.5 day period of the experiment. Females that did not become pregnant and all males were euthanized by CO_2_ asphyxiation and cervical dislocation at the conclusion of the experiment. All experimental procedures were approved by the IACUC under protocol AR21-00010.
Protein extraction, western blotting and ELISA
Protein from snap-frozen whole brains were extracted using RIPA Buffer (Cell Signaling Technology cat# 9806) with protease inhibitor cocktail (ThermoFisher cat# 87785). Concentration was determined prior to loading using the Pierce BCA Assay Kit with subsequent colorimetric detection using BioTek Synergy H1 microplate reader. Samples were denatured in Laemmli buffer at 95°C for 5 min. 50 µg total protein from each sample was loaded onto 10% Mini-PROTEAN® TGX™ Precast Protein Gels (Bio-Rad) and separated for 1 h at 150 V. Proteins were transferred onto nitrocellulose or PVDF membranes using Turbo transfer (Bio-Rad) for 7 min at 2.5 A. Membranes were incubated in Everyblot blocking buffer (Bio-Rad) for 1 h at RT. For Iba-1 probe, membranes were cut at 37 kDa marker and lower half was incubated with Iba1, top half was incubated with b-actin. For Tau and p-Tau Thr231, membranes were cut between 75 and 100 kDa marker. Top half was incubated with vinculin, bottom half was incubated Tau5 (ThermoFisher cat# AHB0042) or phospho-Tau Thr231 (Cell Signaling Technology cat # 71429). A control Tau protein (Acro BioSystems cat # TAU-H5147) with similar molecular mass as mouse Tau was run. All primary antibody incubations were overnight at 4°C. Membranes were washed in 1 X Tris-buffered saline with 0.1% Tween-20 (TBST) prior to a secondary incubation with either goat anti-mouse HRP or goat anti-rabbit HRP at a dilution of 1:3000 for 1 h at RT. Membranes were washed a second time with 1 X TBST prior to a 5 min incubation in ECL reagent (ThermoFisher, SuperSignal West Pico Plus ECL substrate kit, cat# 34577). Blots were imaged on a Bio-Rad Chemidoc. Densitometric analysis was performed in ImageLab Software (Bio-Rad). Target signals were normalized to a control protein on each blot of either Vinculin or β-actin. Aliquots of the RIPA extracted proteins (from above) were used to quantify the cytokines by ELISA for IL-1β (ThermoFisher IL-1 beta Mouse ELISA kit, cat # BMS6002), IL-6 (ThermoFisher IL-6 Mouse ELISA kit, cat # BMS603-2) and TNFα (ThermoFisher TNF alpha Mouse ELISA kit, cat # BMS607-3), from whole brains according to manufacturer’s protocol. All samples were run in duplicate. The average from both wells was used for quantification. A full list of reagents and western blots can be found in the supporting information S4.
Nucleic acid extraction, PCR, and RT-qPCR
DNA and RNA was extracted for sexing embryonic mice and RT-qPCR analysis of cytokine mRNAs present in the pup brains. Nucleic acids were extracted from frozen whole brains using the Omega Bio-tek E.Z.N.A. kit according to manufacturer’s protocol (cat# R6731-01). RNA and DNA concentrations were determined using Qubit Fluorometer (Applied Biosystems) prior to subsequent analysis. For sexing of pups we added 1 µ L DNA to a reaction with the Q5 Master Mix (New England Biolabs, cat # M0492S) and primer sets for amplification of the IL3 (For 5’- GGG ACT CCA AGC TTC AAT CA-3’, Rev 5’- TGG AGG AAG AAA AGC AA-3’) and Sry (For 5’-TGG GAC TGG TGA CAA TTG TC, Rev 5’-GAG TAC AGG TGT GCA GCT CT-3’) genes [61]. PCR conditions were 95°C for 4.5 min followed by 33 cycles of 95°C for 35 s, 50°C for 1 min and 72°C for 1 min. PCR products were terminated with a final extension at 72°C for 5 min. Bands positive for both are labeled as male while samples only expressing IL3 are labeled as female.
cDNAs for RT-qPCR were made by adding 1000 ng RNA to a reaction using the High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, cat# 4368814). 1 µ L of cDNA was added per reaction with gene specific primer pair sets for GAPDH, IL-1β, IL-6, TNFα, IL-4, IL-10, TGFβ, MyD88, NFκΒ, NLRP3 for RT-qPCR (Integrated DNA Technologies, PrimeTime qPCR Primer Assay). All samples were run in triplicate on a QuantStudio6 Pro (Applied Biosystems) using the PowerUp SYBR Green Master Mix (ThermoFisher, cat # A25741). Cycling conditions are as follows: 95°C for 15s followed by 60 °C for 1 min for 40 cycles. Gene expression was measured using the QuantStudio6 Pro detection system and normalized to GAPDH. Ct values were averaged from each triplicate and used to calculate the relative expression by the 2^^-(ΔΔCt)^ method with GAPDH as the reference gene.
Immunofluorescent staining
Fixed brains were embedded in OCT medium and 25 µm sagittal sections were collected by a cryostat. Sections were washed with PBS, blocked in PBS containing 0.1% tritonX-100, and 10% donkey serum and probed with and primary antibodies to Cux1 (Protein Tech 11733-1-AP; diluted 1:300), SatB2 (Abcam ab92446; diluted 1:100), and Ctip2 (Abcam ab-18465; diluted 1:500). Sections were then washed with PBS and incubated with Alexa Fluor® secondary antibodies (ThermoFisher Scientific) and DAPI followed by mounting with Prolong Diamond Antifade (ThermoFisher Scientific). Images were acquired with an ImageXpress Micro Confocal High Content Imaging System (IXMC; Molecular Devices) and analyzed using MetaXpress software (Molecular Devices).
Data analysis
We used the analysis routines in GraphPad PRISM v. 10.0 to perform statistical analysis and plotting of data points. Outlier data points were identified by the ROUT method with Q = 1% and removed from analysis. Data were tested for normality with the Kolmogorov-Smirnov test. Group differences for data with normal distributions were tested with the two-tailed, unpaired Student’s-test, otherwise statistical differences were determined with the Mann Whitney-U test. Data are presented with the median and 95% Confidence Interval (CI).
Supporting Information
S1 FileOMV TEM, gingipain activity and LPS quantification plots.(PDF)
S2 FileIBA-1, Tau and p-TauThr231 western blots.(PDF)
S3 DataData for figures.(XLSX)
S4 FileReagent list.(XLSX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Carrillo-de-Albornoz A, Figuero E, Herrera D, Bascones-Martínez A. Gingival changes during pregnancy: II. influence of hormonal variations on the subgingival biofilm. J Clin Periodontol. 2010;37(3):230–40. doi: 10.1111/j.1600-051X.2009.01514.x 20088983 · doi ↗ · pubmed ↗
- 2Fujiwara N, Tsuruda K, Iwamoto Y, Kato F, Odaki T, Yamane N, et al. Significant increase of oral bacteria in the early pregnancy period in Japanese women. J Investig Clin Dent. 2017;8(1):e 2189. doi: 10.1111/jicd.12189 26345599 · doi ↗ · pubmed ↗
- 3Lin W, Jiang W, Hu X, Gao L, Ai D, Pan H, et al. Ecological shifts of supragingival microbiota in association with pregnancy. Front Cell Infect Microbiol. 2018;8:24. doi: 10.3389/fcimb.2018.00024 29497601 PMC 5819318 · doi ↗ · pubmed ↗
- 4Paropkari AD, Leblebicioglu B, Christian LM, Kumar PS. Smoking, pregnancy and the subgingival microbiome. Scientific Reports. 2016;6(1):30388.27461975 10.1038/srep 30388 PMC 4961950 · doi ↗ · pubmed ↗
- 5Jang H, Patoine A, Wu T, Castillo D, Xiao J. Oral microflora and pregnancy: a systematic review and meta-analysis. Scientific Reports. 2021;11(1):16870.34413437 10.1038/s 41598-021-96495-1PMC 8377136 · doi ↗ · pubmed ↗
- 6Powell AM, Khan FZA, Ravel J, Elovitz MA. Untangling associations of microbiomes of pregnancy and preterm birth. Clin Perinatol. 2024;51(2):425–39. doi: 10.1016/j.clp.2024.02.009 38705650 PMC 11070640 · doi ↗ · pubmed ↗
- 7Wen P, Li H, Xu X, Zhang F, Zhao D, Yu R, et al. A prospective study on maternal periodontal diseases and neonatal adverse outcomes. Acta Odontol Scand. 2024;83:348–55. doi: 10.2340/aos.v 83.40836 38860278 PMC 11302652 · doi ↗ · pubmed ↗
- 8Kassebaum NJ, BernabéE, Dahiya M, Bhandari B, Murray CJL, Marcenes W. Global burden of severe periodontitis in 1990-2010: a systematic review and meta-regression. J Dent Res. 2014;93(11):1045–53. doi: 10.1177/0022034514552491 25261053 PMC 4293771 · doi ↗ · pubmed ↗