Negative effects of nitrogen fertilization on herbivore fitness are exaggerated at warmer temperatures and in high-altitude populations
Ange Raharivololoniaina, Roland Busch, Franziska Deppe, Anna Hitzler, Eva Plath, Tamara Rischen, Mine Yilmazer, Klaus Fischer

TL;DR
This study shows that nitrogen fertilization harms butterfly development more at higher temperatures and in high-altitude populations, highlighting how environmental changes can interact to threaten biodiversity.
Contribution
The study reveals novel interactions between eutrophication and climate change effects on herbivore fitness across different altitude populations.
Findings
Nitrogen fertilization reduces butterfly body mass and prolongs development time.
Negative effects of fertilization are stronger at warmer temperatures and in high-altitude populations.
Populations in nutrient-poor environments may be more vulnerable to agricultural intensification.
Abstract
Biodiversity is currently under strong pressure due to anthropogenic global change. Different drivers of global change may exert direct and indirect effects on biodiversity, and may furthermore interact with one another, but our respective knowledge is still very limited. We investigated indirect and interactive effects of two important drivers of global change, eutrophication and climate change, in replicated low- and high-altitude populations of an insect herbivore, the butterfly Lycaena tityrus, in a laboratory setting. We found local adaptation in developmental traits, with low-altitude populations being adapted to warmer temperatures and longer seasons. Lycaena tityrus responded negatively to agriculturally relevant levels of fertilization of its host plant, showing reduced body mass and prolonged development time. Negative effects were particularly pronounced at warmer…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
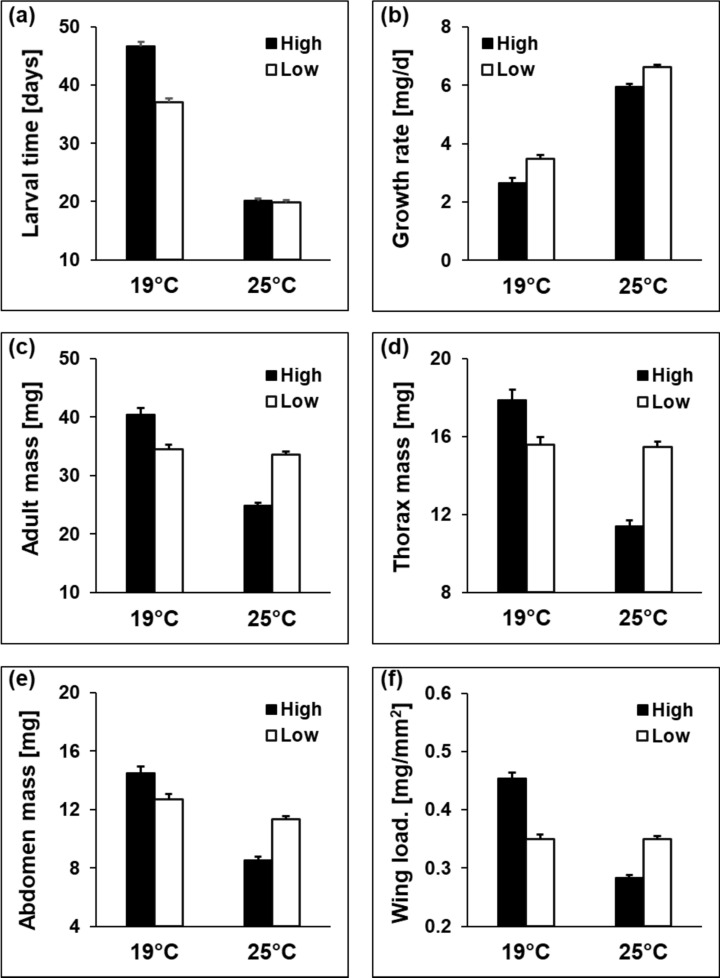 Figure 1
Figure 1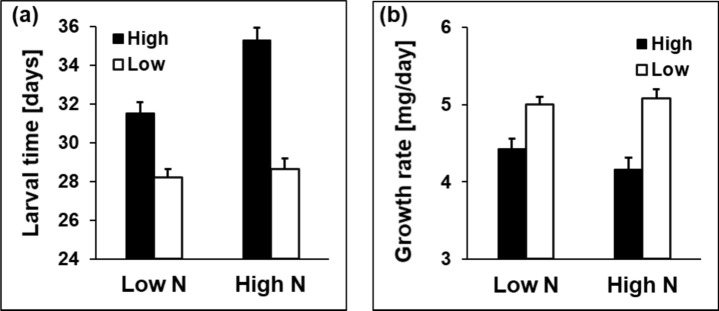 Figure 2
Figure 2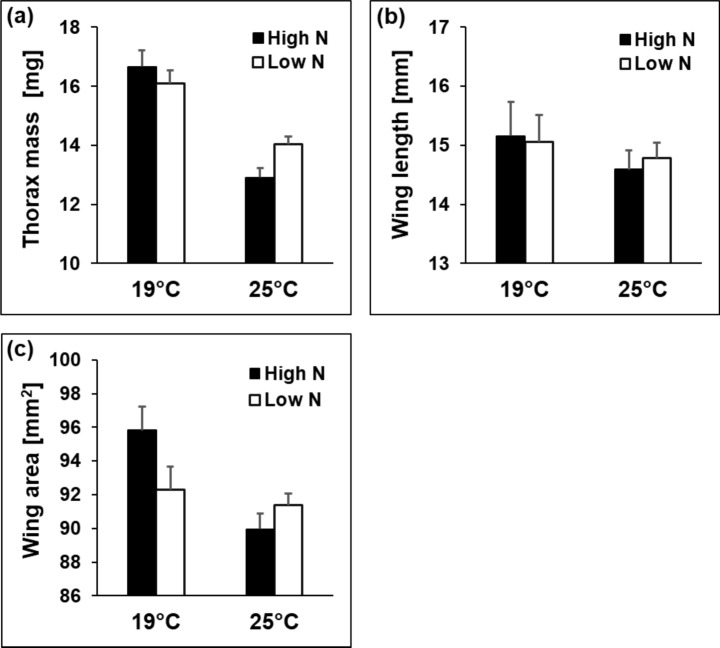 Figure 3
Figure 3- —Forschungsinitiative Land Rheinland-Pfalz
- —Universität Koblenz (6377)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant and animal studies · Ecology and Vegetation Dynamics Studies · Species Distribution and Climate Change
Introduction
In the current era, biodiversity is strongly threatened by global environmental change, which involves a variety of human-induced drivers (Pereira et al. 2012; Ceballos et al. 2015; Johnson et al. 2017; Wagner 2020). These drivers may directly or indirectly affect biodiversity, and may additionally interact with one another (Oliver and Morecroft 2014; Jackson et al. 2016; Gunderson et al. 2017; Hamann et al. 2021). While the direct effects of individual drivers have been quite intensively studied, our knowledge on indirect effects and interactions among drivers is still limited (Oliver and Morecroft 2014; Sage 2020; Wagner 2020; Kuczyk et al. 2021a).
Important drivers of biodiversity loss are agricultural intensification and anthropogenic climate change (Pereira et al. 2012; Ceballos et al. 2015; WallisDeVries and Van Swaay 2017). With regard to agricultural intensification, adverse effects of eutrophication on species richness have been widely documented for plants (Bobbink et al. 2010; Stevens et al. 2010), but also for invertebrates (Van Dyck et al. 2009; Carvalheiro et al. 2019; Nessel et al. 2021). Negative effects of excess nitrogen on animals might be conveyed by microclimatic cooling, reduced host-plant quality, and the loss of host plants or nectar sources (Fischer and Fiedler 2000; WallisDeVries and Van Swaay 2006; Lebeau et al. 2016; Nijssen et al. 2017). However, our knowledge on the effects of nitrogen fertilization on animals is still limited (Nijssen et al. 2017). With regard to anthropogenic climate change, its effects on biodiversity have been widely demonstrated and discussed (Weiskopf et al. 2020; Pereira et al. 2024). Climate change may induce shifts in phenology or geographic ranges, and may also affect morphology, life history and behavior (Scheffers et al. 2016; Weiskopf et al. 2020). Both, eutrophication and climate change, likely play a significant role in biodiversity loss, specifically in the decline of insect diversity and abundance (Nessel et al. 2021; Raven and Wagner 2021).
Herbivores comprise particularly interesting organisms to study the consequences of global change, as they are sensitive to direct as well as indirect effects, the latter being mediated by alterations in host-plant quality or availability (Harvey et al. 2020; Kuczyk et al. 2021a). For instance, increased temperatures during plant growth may diminish host-plant quality for insect herbivores (Bauerfeind and Fischer 2013; Kuczyk et al. 2021a, b), which may result from increased C:N ratios, concentrations of secondary metabolites, and leaf toughness (Meihls et al. 2012; Block et al. 2019). Similarly, nitrogen fertilization, as abundantly applied in modern agriculture, affects host-plant chemistry (Kuczyk et al. 2021a, b). This may in turn diminish herbivore performance, e.g. by reducing survival rates, growth rates and body mass (Fischer and Fiedler 2000; Kurze et al. 2018; Raharivololoniaina et al. 2023).
In species with a wide geographical distribution, populations may respond differently to environmental stressors (Kuczyk et al. 2021b). Such within-species variation in the vulnerability to global change has been largely neglected thus far, although geographical variation in trait values is common (Kawecki and Ebert 2004; Blanquart et al. 2013). For instance, in the widespread butterfly Pieris napi, fast-growing Italian populations are more sensitive to poor food quality than slower growing German ones (Kuczyk et al. 2021b). Such studies on local adaptation provide important insights into the power of natural selection relative to gene flow and other evolutionary forces (Kawecki and Ebert 2004), but add an additional level of complexity.
Here, we study indirect and interactive effects of two important global change drivers, namely eutrophication and a vital component of climate change, viz. increasing temperature. Specifically, we investigate effects of host-plant nitrogen fertilization and herbivore developmental temperature, on high- and low-altitude populations of the butterfly Lycaena tityrus (Poda 1761). This species has been repeatedly used as a model to study phenotypic plasticity, local and thermal adaptation as well as dispersal (e.g., Karl et al. 2008, 2009a, b; Fischer and Karl 2010; Fischer et al. 2011; Reim et al. 2018a, b). These studies revealed, amongst others, that high- compared to low-altitude populations showed longer development times, increased cold- but decreased heat-stress resistance, reduced plasticity in heat resistance, temperature-induced plasticity in developmental traits, and a pronounced sexual dimorphism. In terms of host-plant quality, effects of nitrogen fertilization of host plants (Fischer and Fiedler 2000; Kurze et al. 2018), and nitrogen fertilization in combination with drought stress (Raharivololoniaina et al. 2023) or high plant growing temperature have been investigated (Raharivololoniaina et al. 2021). Results showed detrimental effects of high levels of host-plant fertilization and of drought stress on the species’ performance, but positive effects of limited nitrogen fertilization and of higher plant rearing temperatures. However, earlier studies have not investigated effects of host-plant nitrogen fertilization (1) across altitudes or (2) at different insect developmental temperatures, which are the aims of the current study.
We predict that (1) cold-adapted high-altitude populations of L. tityrus, which are not (yet) exposed to high levels of eutrophication, are more sensitive to host plant fertilization than low-altitude populations, and that (2) detrimental effects of fertilization are more pronounced at higher ambient temperatures, indicating an interactive effect between two important drivers of biodiversity loss, eutrophication and climate change.
Materials and methods
Study organism
Lycaena tityrus (Poda 1761) is a widespread, temperate-zone butterfly ranging from Spain throughout large parts of central and southern Europe to western Asia (Ebert and Rennwald 1991; Bräu et al. 2013). In most parts of its range, it has 2–3 generations per year (Ebert and Rennwald 1991; Tolman and Lewington 2008). Two subspecies are recognized in central Europe, the widespread nominate form L. tityrus tityrus (Poda 1761) and the alpine subspecies L. tityrus subalpinus (Speyer 1851). The latter is confined to the higher altitudes of the European Alps and some other mountain ranges, where it has only one generation per year (Tolman and Lewington 2008). The altitudinal distribution of L. tityrus subalpinus ranges from 1200 to 2500 m a.s.l. (Tolman and Lewington 2008).
Lycaena tityrus inhabits humid to moderately dry locations such as low-intensity grassland, alpine meadows, sandy heathland, bogs, and forest edges (Ebert and Rennwald 1991; Bräu et al. 2013). It uses different species of sorrel (Rumex spec., Polygonaceae) as larval host plant (Ebert and Rennwald 1991; Tolman and Lewington 2008). The primary hostplant is Rumex acetosa L., which is a perennial herb containing a variety of primarily non-nitrogenous secondary compounds, especially oxalic acids (Korpelainen and Pietiläinen 2020). Adults are rather opportunistic nectar feeders, using a wide array of plants (Ebert and Rennwald 1991). Lycaena tityrus has suffered substantial population losses due to grassland intensification, meadow conversion, and afforestation in recent decades (Bräu et al. 2013). It is projected to shift its distributional range northward and upward due to anthropogenic climate change (Settele et al. 2008).
Population sampling
For this study, we collected in 2022 fecund females from four replicated low-altitude populations in northeastern and western Germany (L. tityrus tityrus; Greifswald, coordinates: 54.05°N, 13.44°E; Altwarp: 53.74°N, 14.26°E; Pottum: 50.59°N, 8.00°E; Westerburg: 50.54°N, 7.97°E; altitudinal range 0–420 m a.s.l.), and from four replicated high-altitude populations in Austria (L. tityrus subalpinus; Kühtai: 47.23°N, 10.97°E; Obernberg: 46.98°N, 11.42°E; Obergurgl: 46.86°N, 11.02°E; Venn: 47.01°N, 11.54°E; altitudinal range 1560–2030 m a.s.l.). All high-altitude populations were sampled on unfertilized alpine meadows in the central Alps, i.e., above crystalline primary rock (gneiss, slate) resulting in acidic, nutrient-poor soils. The low-altitude populations, in contrast, originate from slightly fertilized meadows from areas that are also naturally richer in nutrients (basalt mountains or marshland). Note that sampling locations not only differed in altitude but also in latitude. However, the impact of variation in altitude is expected to surpass the potential impact of latitude by far (Karl et al. 2008, 2009a). Per population, we sampled between 6 and 20 females, in total 34 from low- and 48 from high-altitude populations. Females were afterwards transferred to and randomly distributed among two climate cabinets (26 °C, 70% humidity, L19:D5 photoperiod; Panasonic MIR-554) at Koblenz University for egg laying. Therefore, females were kept individually in translucent plastic pots (1 L) covered with gauze. They were provided sucrose solution and fresh flowers for adult feeding, and leaves of Rumex acetosa as oviposition substrate. Deposited eggs were collected daily and transferred, separated by female, to small glass vials kept in one climate cabinet at 22 °C (70% humidity, L19:D5 throughout) until hatching (Panasonic MIR-554). Hatchlings were fed on field-collected R. acetosa leaves under the same conditions for 4 days, after which they were allocated to treatments.
Experimental design
On day 5 after hatching, larvae were individually transferred to transparent plastic boxes (250 ml) lined with moist tissue and containing a fresh cutting of R. acetosa. Larvae were randomly allocated to one out of four treatments, using a split-brood design. We employed a full-factorial design with two rearing temperatures and two host plant treatments. Rearing temperatures had a mean of 19.2 °C (light phase 20 °C, dark phase 16 °C; L19:D5 photoperiod) or 25.2 °C (light phase 26 °C, dark phase 22 °C), and we used one climate cabinet per temperature (Panasonic MIR-554). To rule out effects of individual cabinets, temperature regimes and according larvae were shifted among both cabinets every four days. Host plant treatments involved low versus high levels of nitrogen fertilization, the latter mimicking levels typically used in modern agriculture (Raharivololoniaina et al. 2021, 2023). Host plants, 200 per treatment, were grown individually in standard pots (9 × 9 × 10 cm, 0.5 L) in a greenhouse from commercially available seeds. Plants were provided with a common nitrogenous fertilizer (NH_4_NO_3_), and received a total amount of 1.225 ml (low N) or 3.675 ml (high N) of NH_4_NO_3_. The total amount of NH_4_NO_3_ was equally divided into five applications of 0.245 ml (low N) or 0.735 ml (high N) in 100 ml water each, corresponding to one application per week during the larval feeding phase. These nitrogen treatments have been shown to be highly effective in changing plant chemistry; specifically, higher nitrogen levels resulted in higher nitrogen and water contents but a reduced C:N ratio (Raharivololoniaina et al. 2021, 2023). All boxes were cleaned daily and supplied with a fresh leaf cutting and moist tissue. Leaf cuttings were taken from at least 20 plants per day and randomly allocated to the larvae within treatments. Boxes were shifted around daily to minimize potential temperature and light differences within the climate cabinets. All offspring (n = 579) were reared at the treatment they were allocated to until adult eclosion. Afterwards, the sex of each individual was determined and the animals frozen at -18 °C for later analyses.
Data collection and laboratory analyses
For each individual, we scored different developmental and morphological traits as well as mortality. Larval and pupal development time were measured in days. Pupal mass was weighed on the day following pupation to the nearest 0.1 mg (KERN ABJ-NM/ABS-N). Larval growth rate was calculated as mean weight gain per day (LN (pupal mass)/larval time). Frozen butterflies were first weighed to obtain total adult mass. Then, wings, head and legs were removed, and the abdomen was separated from the thorax. Abdomen and thorax were weighed separately, and thorax-abdomen ratio was calculated as a measure of the relative investment into flight versus reproduction. The length and area of the right forewings were measured using digital images and the software ImageJ. Wing loading was calculated as adult body mass divided by forewing area, and wing aspect ratio as 4 × forewing length^2^ divided by forewing area (Berwaerts et al. 2002). Finally, abdomen fat content was quantified, but due to the high work load only in a subset of individuals, following Fischer et al. (2003). In short, abdomens were dried at 60 °C for 24 h and then weighed. Thereafter, fat was extracted from the dried abdomens for 48 h on a laboratory shaker using 1.5 mL of dichloromethane (CH_2_CL_2_) for each abdomen. The extraction was repeated once, after which the abdomens were once again dried at 60 °C for 24 h and afterwards weighed. Abdomen fat content was then calculated by subtracting the fat-free abdomen dry mass from the initial abdomen dry mass and is given in percent.
Statistical analyses
We used general linear mixed models (GLMMs) to analyze all traits obtained (except survival, see below), with origin (low versus high altitude), sex, developmental temperature, and host-plant nitrogen treatment as fixed factors, and replicate population (nested within origin) and family (the offspring of a given female; nested within origin and population) as random factors. Minimum adequate models were constructed by sequentially removing non-significant interaction terms, in case no higher order interactions were significant. Larval time, pupal time, and larval growth rate were LN-transformed prior to analyses to meet GLMM requirements. Survival data were analyzed using a binary logistic regression with the same factors and model structure as above. All tests were computed using Statistica 12.0 (Statsoft, Tulsa, USA) or R 4.4.2 (R Core Team 2024) using the ‘lme4’ package (Bates et al. 2015). Throughout the text, we present means ± standard error.
Results
Low- and high-altitude populations differed significantly in larval time, pupal time, larval growth rate, wing length, wing area, wing loading, wing aspect ratio, and abdomen fat content (Table 1). High- compared with low-altitude populations showed longer larval times (33.4 ± 0.4 vs. 28.4 ± 0.4 d), shorter pupal times (10.5 ± 0.1 vs. 11.5 ± 0.1 d), lower larval growth rates (4.30 ± 0.10 vs. 5.04 ± 0.08 mg/d), shorter forewings (14.44 ± 0.06 vs. 15.26 ± 0.05 mm), smaller forewing areas (88.0 ± 0.8 vs. 96.5 ± 0.6 mm^2^), higher wing loadings (0.369 ± 0.006 vs. 0.350 ± 0.005 mg/mm^2^), and lower wing aspect ratios (9.59 ± 0.06 vs. 9.72 ± 0.04) and abdomen fat contents (9.6 ± 0.9 vs. 17.3 ± 0.7%). In addition, thorax masses tended to be lower in high- versus low-altitude populations (14.7 ± 0.3 vs. 15.5 ± 0.2 mg).Table 1. Results of general linear mixed models for the effects of origin (low vs. high altitude), sex, rearing temperature, host-plant nitrogen fertilization (low vs. high; all fixed), replicate population (random, nested within origin), and family (random, nested within population and origin) on various traits of the butterfly Lycaena tityrusSourceEffectDFnDFdMSF**pLarval time OriginFixed180.595.630.0470 SexFixed15341.2830.72**< 0.0001** TemperatureFixed153450.031205.15**< 0.0001** NitrogenFixed15340.307.150.0077 OriginTempFixed15341.0425.00**< 0.0001** OriginNitroFixed15340.348.180.0044 Population (Origin)Random6780.132.650.0217 Family (OriginPop)Random315340.051.280.1472 Error5340.04Pupal time OriginFixed170.3924.680.0015 SexFixed15250.3945.01**< 0.0001** TemperatureFixed152520.632402.60**< 0.0001** NitrogenFixed1525< 0.010.060.8133 Population (Origin)Random6640.021.330.2563 Family (OriginPop)Random315250.021.820.0049 Error5250.01Pupal mass OriginFixed181175.781.880.2066 SexFixed15297075.6424.21**< 0.0001** TemperatureFixed1529694.772.380.1237 NitrogenFixed1529367.531.260.2626 OriginSexFixed152921.960.080.7841 OriginTempFixed15291051.883.600.0584 SexTempFixed15291.29< 0.010.9470 OriginNitroFixed1529337.251.150.2832 SexNitroFixed1529510.611.750.1868 TempNitroFixed1529819.632.800.0946 OriginTempNitroFixed15291297.524.440.0356 Population (Origin)Random654732.401.400.2303 Family (OriginPop)Random31529698.632.390.0001 Error529292.28Larval growth rate OriginFixed181.1911.600.0094 SexFixed15310.244.870.0277 TemperatureFixed153149.761015.84**< 0.0001** NitrogenFixed15310.183.670.0560 OriginTempFixed15310.5912.140.0005 OriginNitroFixed15310.295.850.0159 Population (Origin)Random6810.122.220.0494 Family (OriginPop)Random315310.061.200.2111 Error5310.05Adult mass OriginFixed11519.930.430.5237 SexFixed153210,338.96187.37**< 0.0001** TemperatureFixed15326496.44117.73**< 0.0001** NitrogenFixed1532297.575.390.0206 OriginTempFixed15325645.05102.30**< 0.0001** Population (Origin)Random65039.720.340.9114 Family (OriginPop)Random31532165.613.00**< 0.0001** Error53255.18Thorax mass OriginFixed11548.444.540.0504 SexFixed153383.116.270.0125 TemperatureFixed15331079.8381.530.0000 NitrogenFixed153318.841.420.2336 OriginSexFixed153387.406.600.0105 OriginTempFixed1533960.2772.50**< 0.0001** TempNitroFixed153354.664.130.0427 Population (Origin)Random6549.200.400.8776 Family (OriginPop)Random3153330.742.320.0001 Error53313.25Abdomen mass OriginFixed192.440.130.7318 SexFixed15356287.01601.96**< 0.0001** TemperatureFixed15351336.71127.99**< 0.0001** NitrogenFixed153547.684.560.0331 OriginTempFixed1535560.0053.62**< 0.0001** SexTempFixed1535110.8210.610.0012 Population (Origin)Random64922.121.000.4343 Family (OriginPop)Random3153530.972.97**< 0.0001** Error53510.44Thorax-abdomen ratio OriginFixed170.440.710.4290 SexFixed1536136.19727.77**< 0.0001** TemperatureFixed15362.3912.770.0004 NitrogenFixed15360.180.950.3296 Population (Origin)Random6630.682.520.0298 Family (OriginPop)Random315360.331.770.0069 Error5360.19Wing length OriginFixed11028.7536.560.0001 SexFixed15262.694.700.0306 TemperatureFixed15268.1614.260.0002 NitrogenFixed1526< 0.01< 0.010.9667 OriginSexFixed15264.638.090.0046 OriginTempFixed15260.480.850.3580 SexTempFixed15262.294.000.0460 OriginNitroFixed15261.051.840.1755 SexNitroFixed15260.220.390.5344 TempNitroFixed15263.586.250.0127 OriginTempNitroFixed15263.165.520.0192 Population (Origin)Random6520.840.760.6010 Family (OriginPop)Random315261.502.62**< 0.0001** Error5260.57Wing area OriginFixed192724.4421.460.0011 SexFixed152910,610.74125.16**< 0.0001** TemperatureFixed1529393.024.640.0318 NitrogenFixed1529261.253.080.0798 TempNitroFixed1529872.5410.290.0014 OriginTempNitroFixed3529246.842.910.0340 Population (Origin)Random655138.860.950.4643 Family (OriginPop)Random31529191.552.260.0002 Error52984.78Wing loading OriginFixed1280.028.960.0057 SexFixed15270.55115.06**< 0.0001** TemperatureFixed15270.74155.66**< 0.0001** NitrogenFixed15270.047.870.0052 OriginSexFixed15270.0715.310.0001 OriginTempFixed15270.73153.70**< 0.0001** SexTempFixed15270.035.850.0159 Population (Origin)Random652< 0.010.180.9805 Family (OriginPop)Random315270.012.66**< 0.0001** Error527< 0.01Wing aspect ratio OriginFixed1132.3310.700.0063 SexFixed153248.5799.97**< 0.0001** TemperatureFixed15324.088.390.0039 NitrogenFixed15322.815.790.0165 SexTempFixed15322.194.500.0344 Population (Origin)Random6710.170.280.9428 Family (OriginPop)Random315320.691.430.0643 Error5320.49Fat content OriginFixed171735.7211.440.0110 SexFixed1315311.613.690.0556 TemperatureFixed1315119.531.420.2349 NitrogenFixed1315141.321.670.1966 Population (Origin)Random6105166.852.230.0456 Family (OriginPop)Random2731566.580.790.7662 Error31584.39Minimum adequate models were constructed by sequentially removing non-significant interaction terms, as far as no higher order interactions were significant. Significant p-values are given in bold
The above general patterns were modulated by interactions with other factors (Table 1). For six traits, the interaction between origin and rearing temperature was significant. These interactions show for larval time and larval growth rate that differences among altitudes were restricted to or more pronounced at the lower temperature (Fig. 1). Adult, thorax and abdomen mass were higher in high-altitude populations at the lower rearing temperature, but lower at the higher rearing temperature. Accordingly, wing loading was higher in high- than in low-altitude populations at the lower temperature, but the other way around at the higher temperature (Fig. 1f). The significant origin by nitrogen interactions for larval time and larval growth rate show that the high-altitude populations responded much more strongly to the high nitrogen treatment, prolonging larval time and reducing larval growth rates (Fig. 2).Fig. 1. Larval time (a), larval growth rate (b), adult mass (c), thorax mass (d), abdomen mass (e), and wing loading (f) in relation to origin (high vs. low altitude) and rearing temperature (19 vs. 25 °C) in the butterfly Lycaena tityrus. Given are means ± SEFig. 2Larval time (a) and larval growth rate (b) in relation to origin (high vs. low altitude) and host-plant nitrogen fertilization (low N vs. high N) in the butterfly Lycaena tityrus. Given are means ± SE
All measured traits differed significantly among sexes except from fat content, for which there was only a tendency towards a lower fat content in females (12.5 ± 0.8 vs. 14.4 ± 0.7%). Females compared with males had longer larval (32.2 ± 0.4 vs. 29.6 ± 0.4 d) and pupal times (11.2 ± 0.1 vs. 10.7 ± 0.1 d), higher pupal masses (126.5 ± 1.3 vs. 118.2 ± 1.2 mg), lower larval growth rates (4.57 ± 0.10 vs. 4.77 ± 0.10 mg/d), higher adult (38.2 ± 0.6 vs. 28.5 ± 0.5 mg), thorax (15.5 ± 0.3 vs. 14.7 ± 0.3 mg) and abdomen masses (15.6 ± 0.3 vs. 7.9 ± 0.2 mg), lower thorax-abdomen ratios (1.04 ± 0.03 vs. 2.00 ± 0.03), longer wings (15.0 ± 0.1 vs. 14.8 ± 0.1 mm), larger wing areas (96.6 ± 0.7 vs. 87.9 ± 0.7 mm^2^), higher wing loadings (0.395 ± 0.005 vs. 0.323 ± 0.005 mg/mm^2^), and lower wing aspect ratios (9.33 ± 0.05 vs. 9.99 ± 0.05). The interaction between origin and sex was significant for thorax mass, wing length, and wing loading, showing that sex differences were restricted to or more pronounced in high-altitude (thorax mass, wing loading) or low-altitude populations (wing length; Table S1b, supplementary material).
All traits except from pupal mass and abdomen fat content differed among rearing temperatures. At the lower rearing temperature, larval (41.9 ± 0.5 vs. 20.0 ± 0.3 d) and pupal times (13.4 ± 0.1 vs. 8.5 ± 0.1 d) were longer, larval growth rate (3.06 ± 0.11 vs. 6.28 ± 0.06 mg/d) was lower, adult (37.5 ± 0.7 vs. 29.2 ± 0.4 mg), thorax (16.7 ± 0.3 vs. 13.5 ± 0.2 mg) and abdomen masses (13.6 ± 0.3 vs. 9.9 ± 0.2 mg) were higher, thorax-abdomen ratio (1.42 ± 0.04 vs. 1.59 ± 0.02) was lower, wings (15.0 ± 0.1 vs. 14.7 ± 0.1 mm) were longer, wing area (93.3 ± 0.8 vs. 91.1 ± 0.5 mm^2^) was larger, and wing loading (0.402 ± 0.006 vs. 0.316 ± 0.004 mg/mm^2^) and wing aspect ratio (9.77 ± 0.06 vs. 9.54 ± 0.04) were higher. The reductions in abdomen mass and concomitantly wing loading at the higher temperature were more pronounced in females than in males. However, the reductions in wing length and wing aspect ratio at the higher temperature were more pronounced in males than in females (significant sex by temperature interactions; Table S1a, supplementary material). The reduction in thorax mass, wing length and area at the higher temperature were more pronounced in the high nitrogen treatment (significant temperature by nitrogen treatment interactions; Fig. 3). For the latter two traits, this effect was especially pronounced in high altitude populations (significant three-way interactions; Table S2, supplementary material).Fig. 3. Thorax mass (a), wing length (b), and wing area (c) in relation to temperature (19 vs. 25 °C) and host-plant nitrogen treatment (low N vs. high N) in the butterfly Lycaena tityrus. Given are means ± SE
Host-plant nitrogen treatment significantly affected larval time, adult mass, abdomen mass, wing loading, and wing aspect ratio. At the high as compared with the low nitrogen treatment larval time (32.0 ± 0.4 vs. 29.9 ± 0.4 d) was longer, adult (33.1 ± 0.6 vs. 33.6 ± 0.5 mg) and abdomen masses (11.6 ± 0.3 vs. 12.0 ± 0.2 mg) were lower, and wing loading (0.351 ± 0.006 vs. 0.368 ± 0.005 mg/mm^2^) and wing aspect ratio (9.54 ± 0.05 vs. 9.76 ± 0.05) were lower. Additionally, larval growth rate tended to be lower in the high nitrogen treatment (4.62 ± 0.10 vs. 4.72 ± 0.08 mg/day). The significant origin by temperature by nitrogen interaction indicates that nitrogen fertilization had generally little impact on pupal mass, except from the high-altitude populations at the lower rearing temperature, where high nitrogen resulted in heavier pupae (Table S2, supplementary material).
Significant variation between replicate populations were found in larval time, larval growth rate, thorax-abdomen ratio, and fat content, and between families in pupal time, pupal mass, adult mass, thorax mass, abdomen mass, thorax-abdomen ratio, wing length, wing area, and wing loading (Table 1). This suggests genetic variation in development time, body size, storage reserves, and the allocation to dispersal versus reproduction among populations and / or families.
Survival rates, finally, only differed significantly among rearing temperatures (19 °C: 55.1%, 25 °C: 85.2%; z = − 4.55, p < 0.0001), but not in relation to origin (low altitude: 84.0%, high altitude: 72.2%; z = − 1.37, p = 0.1715) or nitrogen treatment (low N: 84.2%, high N: 71.1%; z = − 1.06, p = 0.2884). All interactions were non-significant (all p-values > 0.14).
Discussion
Our study revealed evidence for local genetic adaptation in high- and low-altitude populations of the butterfly Lycaena tityrus. Specifically, low-altitude populations showed an overall shorter development time associated with higher larval growth rates (cf. Karl et al. 2008), presumably as an adaptation to having 2–3 generations per year in contrast to the high-altitude populations which are univoltine (Tolman and Lewington 2008). Thus, the time constraints imposed by having to fit in additional generations a year seem to be much more severe than those imposed by the shorter vegetation period in alpine environments (Roff 1980; Nylin and Gotthard 1998; Burke et al. 2005; Karl et al. 2008). Interestingly, Estonian populations of L. tityrus also show shorter development times (and smaller body size) compared to German populations, presumably as an adaptation to the short season length in Estonia (Reim et al. 2018b). However, Estonian populations have two generations a year, supporting the above conclusions (Reim et al. 2018b). Interestingly, the differences in larval development time were restricted to the lower rearing temperature, at which also the difference in larval growth rate was more pronounced. This may indicate that the low-altitude populations speed up their development as much as possible if ambient conditions are challenging for quick development (Nylin and Gotthard 1998), a pressure univoltine high-altitude populations do not face.
Further evidence for local adaptation stems from the fact that high-altitude populations gained a higher body mass than the low-altitude ones at the lower rearing temperature, but vice versa at the higher rearing temperature. Thus, animals were most efficient under the thermal conditions they are presumably better adapted to, probably reflecting metabolic cold adaptation and associated costs at warmer temperatures in high-altitude populations (see Addo-Bediako et al. 2002; Terblanche et al. 2009).
High-altitude populations seem to invest less into dispersal, having smaller wings (despite a similar body mass) and concomitantly higher wing loadings, lower wing aspect ratios and relative abdomen fat contents, and tending to have lower thorax masses as compared with low-altitude populations. Typically, large thoraxes and wings as well as low wing loadings and aspect ratios indicate a high dispersal ability and an aerodynamically- and cost-efficient flight (Berwaerts et al. 2002; Almbro and Kullberg 2012; Lion et al. 2023), while abdominal fat is used to fuel dispersal (Arrese and Soulages 2010; Toprak et al. 2020). However, in general insects seem to have increased rather than decreased wing sizes at higher altitudes (Dillon et al. 2006). The opposite pattern revealed in our study, i.e., smaller wings and a lower investment into dispersal in high-altitude populations, might result from inhabiting relatively continuous habitats along alpine streams (Trense et al. 2021), and/or from a lower dispersal propensity due to an exposure to stronger winds at higher altitudes (Hodkinson 2005).
In general, insect herbivores are often nitrogen limited, typically referred to as the nitrogen limitation hypothesis (White 1993). However, in our study, high levels of host plant fertilization as used in modern agriculture, thus exceeding natural nutrient levels, negatively affected fitness components in L. tityrus, reducing growth rates, body mass and concomitantly wing loading, and prolonging larval time (cf. Fischer and Fiedler 2000; Kurze et al. 2018; Raharivololoniaina et al. 2023). Note that low levels of host plant nitrogen fertilization often increases insect fitness in line with nitrogen limitation hypothesis (White 1993; Kurze et al. 2017; Raharivololoniaina et al. 2021), while high levels may exert negative effects. Thus, herbivore performance in response to host plant nitrogen levels likely follows a hump-shaped curve (Han et al. 2014; Tao et al. 2014; Lebigre et al. 2018). Such negative effects may even include increased mortality (Fischer and Fiedler 2000), though here we could only find sublethal effects.
Consequently, L. tityrus seems highly vulnerable to agricultural intensification not only due to more frequent grassland cutting or more intensive grazing, but also due to increasing nutrient loads. Possible mechanisms behind negative effects of increased nitrogen fertilization on insect herbivores include host-plant changes in elemental stoichiometry, nutritional geometry, essential micronutrients, and allelochemicals (Vogels et al. 2023). As predicted, detrimental effects were more pronounced in high-altitude populations, such that these populations, likely adapted to low levels of host-plant nitrogen, are even more vulnerable to agricultural intensification. Note that all alpine populations were sampled in the central Alps on unfertilized meadows above crystalline primary rock, resulting in acidic, nutrient-poor soils. In contrast, sampling locations of low-altitude populations were on partly fertilized meadows above less acidic soils (basalt, marshland). This also suggests that our data probably do not reflect recent adaptations to different levels of nitrogen availability, but rather long-term local adaptation. In any case, our results are in agreement with the notion that herbivores from naturally nutrient-poor environments should be more strongly affected than those from richer environments (Vogels et al. 2023). Furthermore, detrimental effects of nitrogen fertilization were more pronounced at the higher rearing temperature, indicating increasing risks from climate change. Similarly, additive effects between host-plant nitrogen fertilization and drought were found in L. tityrus (Raharivololoniaina et al. 2023).
The effects of sex and temperature on life-history traits were as expected. Female insects are typically selected for large body size and a high investment into the abdomen, which is positively related to fecundity (Honek 1993; Nylin and Gotthard 1998; Teder et al. 2021). Males, in contrast, have a selective premium on rapid development to eclose before the females (protandry) and on flight capacity, both increasing their reproductive success (Wiklund et al. 1991; Berwaerts et al. 2006; Reim et al. 2018b). As generally found in ectotherms, development time was longer and growth rates lower at the lower developmental temperature (Von Bertalanffy 1960; Karl and Fischer 2008). According to the temperature-size rule on plasticity in body size (Atkinson and Sibly 1997; Karl and Fischer 2008), animals were larger at the lower temperature, also resulting in higher wing loadings. The higher mortality rates at the lower rearing temperature are likely associated with the concomitantly much longer development time.
In summary, we show local adaptation in developmental traits in the butterfly L. tityrus. Low-altitude populations were adapted to warmer temperatures and longer seasons, displaying a more rapid development and phenotypes indicative of a high dispersal capacity. Importantly, we also show that L. tityrus is vulnerable to agricultural intensification, responding negatively to agriculturally relevant levels of nitrogen fertilization of its host plant. According effects were particularly pronounced at warmer temperatures and in high-altitude populations. Thus, our study adds to the increasing knowledge that different drivers of global change, here nutrient loading and climate change, may interact and thereby increase the overall level of threat to biodiversity (Raven and Wagner 2021). Furthermore, our study suggests that populations inhabiting nitrogen-poor environments, as can be assumed for the alpine populations investigated here, might be even more vulnerable to agricultural intensification than others, which may be applicable to many species (see also Turlure et al. 2013). A much lower level of nitrogen input in the habitats of the alpine populations investigated here compared to those of the low-altitude populations is very likely, resulting from an absence of any fertilization of these remote alpine meadows in combination with acidic ground rocks and a low level of atmospheric nitrogen deposition in contrast to central and northern Germany (Walter 2007; European Environment Agency 2024).
In L. tityrus, high-altitude populations seem to be especially vulnerable, showing a high sensitivity to high nitrogen levels, reduced dispersal capacity, and reduced plasticity in heat resistance (Karl et al. 2008, 2009b). We think that our findings, specifically the detrimental effects of agriculturally relevant levels of nitrogen fertilization being exaggerated at warmer temperatures and in populations from nutrient-poorer environments, may have important implications for other vulnerable species in the face of rapid environmental change. In particular, nitrogen fertilization and deposition should be reduced especially in naturally nutrient-poor environments. Future research should investigate whether according species, populations, and communities are indeed more vulnerable to nutrient input, and which mechanisms may be responsible for detrimental effects.
Supplementary Information
Below is the link to the electronic supplementary material.Supplementary file1 (DOCX 29 KB)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1European Environment Agency (2024). Risk of eutrophication measured as exceedance of critical loads of nitrogen deposition in Europe, in 2022. www.eea.europa.eu.