Shigella sonnei and Shigella flexneri infection in Caenorhabditis elegans led to species-specific regulatory responses in the host and pathogen
Bao Chi Wong, Hock Siew Tan

TL;DR
This study shows that Shigella sonnei and Shigella flexneri cause different responses in the host and pathogen during infection in C. elegans.
Contribution
The study reveals species-specific regulatory responses in both host and pathogen during Shigella infections.
Findings
S. sonnei infection downregulates sphingolipid metabolism, cadmium ion response, and xenobiotic response in C. elegans.
S. sonnei upregulates biofilm formation and energy generation/conservation compared to S. flexneri.
Nematode defense response is downregulated during both Shigella infections.
Abstract
In recent decades, Shigella sonnei has surpassed Shigella flexneri as the leading cause of shigellosis, possibly due to species-specific differences in their transcriptomic responses. This study used dual RNA sequencing to analyse the transcriptomic responses of Caenorhabditis elegans and the two Shigella species at early (10 minutes) and late (24 hours) stages of infection. While the nematode defence response was downregulated during both Shigella infections, only infection by S. sonnei led to downregulation of sphingolipid metabolism, cadmium ion response and xenobiotic response in C. elegans. Furthermore, S. sonnei upregulates biofilm formation and energy generation/conservation during infection, acid resistance-related genes and biofilm regulators compared to S. flexneri. These findings highlight species-specific responses during C. elegans infection.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
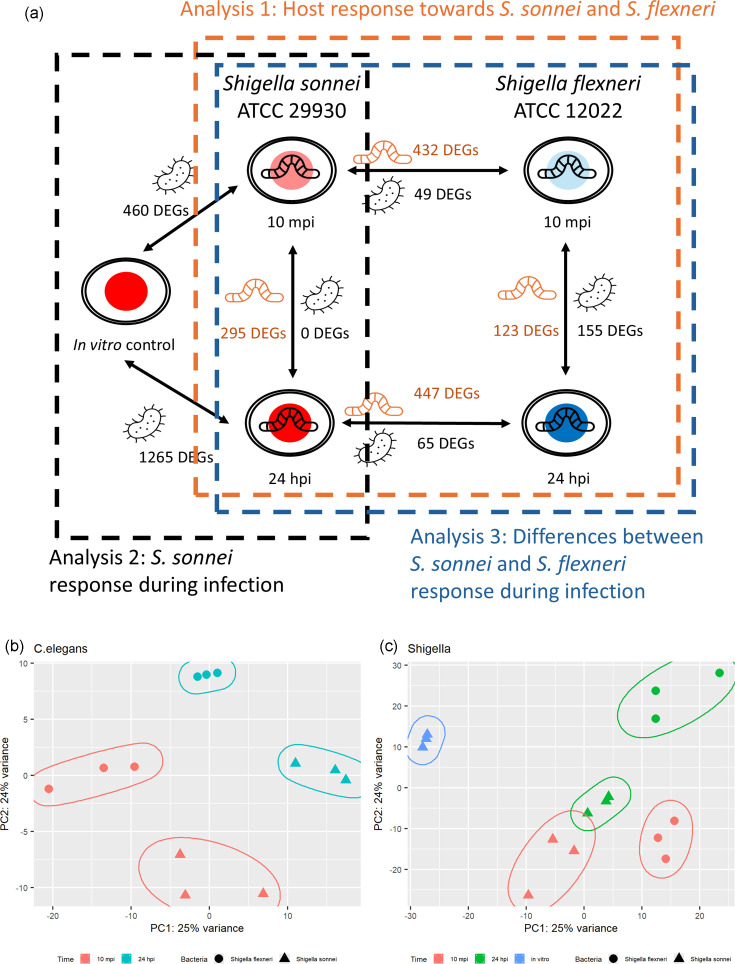 Fig. 1
Fig. 1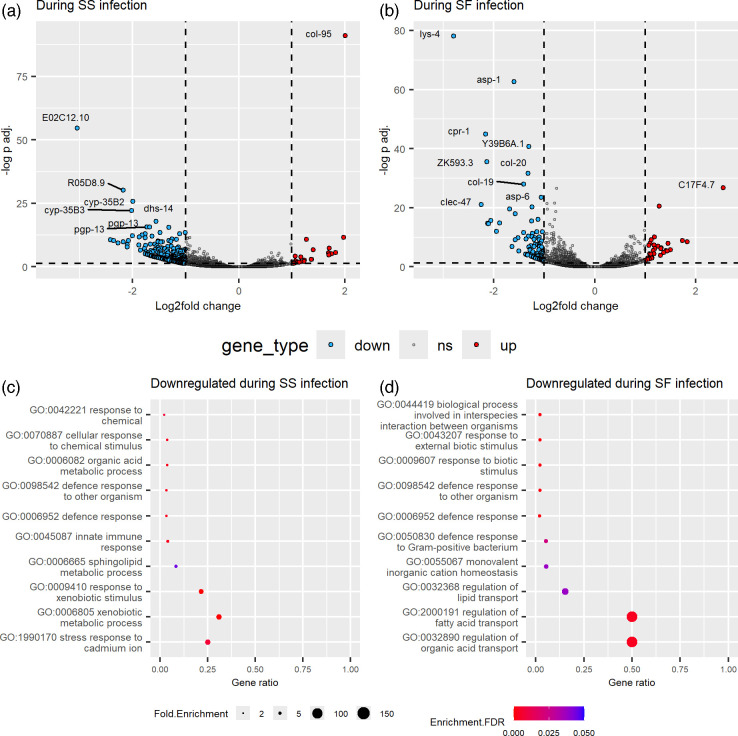 Fig. 2
Fig. 2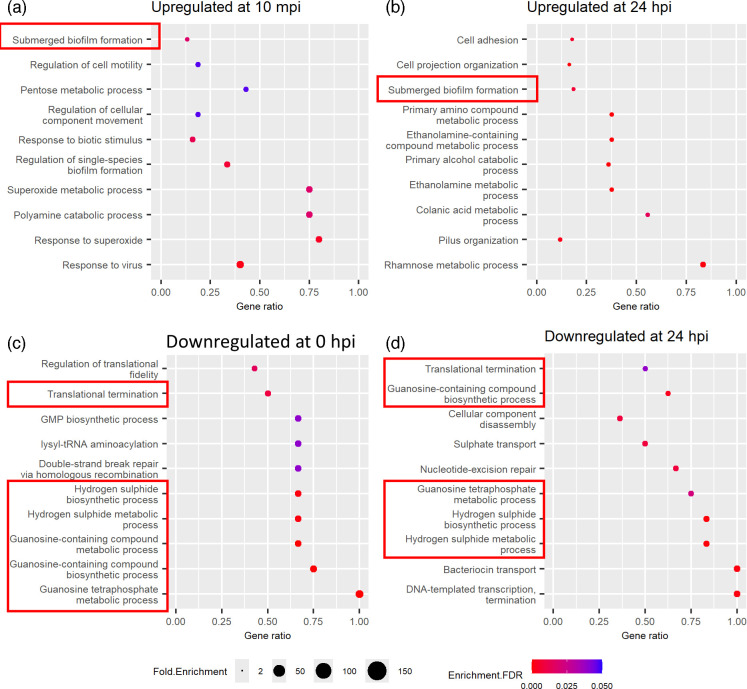 Fig. 3
Fig. 3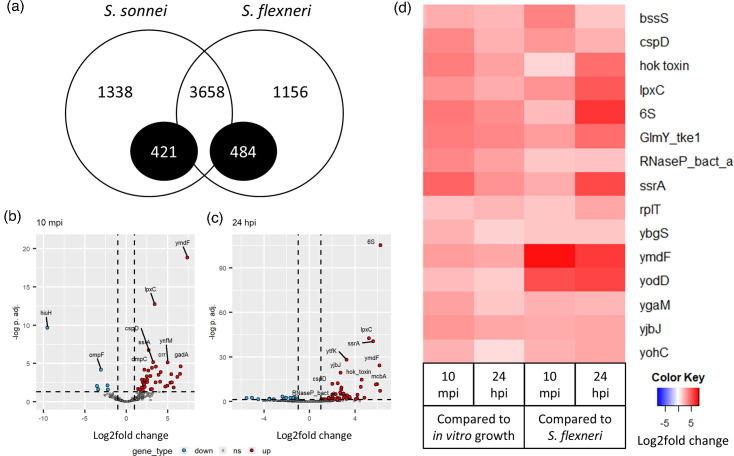 Fig. 4
Fig. 4- —Monash University Malaysia High Impact Research Support Fund Award 2022
- —Ministry of Higher Education Malaysia Fundamental Research Grant Scheme
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEscherichia coli research studies · Gut microbiota and health · Clostridium difficile and Clostridium perfringens research
Data Availability
Trimmed paired-end data of the dual RNA sequencing (Caenorhabditis elegans–Shigella sonnei and Caenorhabditis elegans–Shigella flexneri) are available from the Sequencing Read Archive under BioProject PRJNA1128419 [10 minutes post-infection (mpi)] and PRJNA1147013 [24 hours post-infection (hpi)]. Raw sequencing data of S. sonnei on nematode growth medium (NGM) agar (in vitro) are available under BioProject PRJNA1148041. Other datasets used and analysed during the current study are available from the corresponding author upon reasonable request.
Introduction
Shigella is the bacterial species responsible for causing shigellosis, an intestinal infection associated with aggressive and bloody diarrhoea [1]. Among the four species of Shigella, Shigella flexneri is associated with resource-poor countries, while Shigella sonnei is associated with developed countries [23]. As more countries undergo significant industrialization, there is an increase in the proportion of S. sonnei isolates compared to S. flexneri [38]. Much information on the virulence and pathogenesis of Shigella was obtained from S. flexneri studies and generalized to other Shigella species. However, recent studies have demonstrated the species-specific differences between S. flexneri and S. sonnei. For instance, it is commonly reported that S. flexneri uses the type III secretion system (T3SS) during macrophage internalization [910]. However, S. sonnei’s internalization into macrophages is independent of T3SS [11]. The O-antigen of S. sonnei is also different from other Shigella [12], which helps decrease macrophage internalization and pyroptosis [11] and increases virulence in the zebrafish model [13]. Other than that, S. sonnei also uses colicins to outcompete other bacteria during infection [1415]. These studies demonstrated that S. sonnei studies are necessary due to the species-specific differences from S. flexneri.
Caenorhabditis elegans is a soil-dwelling nematode that has been used as an animal model for bacterial infection for multiple bacterial species including Pseudomonas aeruginosa [1617], Staphylococcus aureus [1819] and Shigella [2023]. The intestinal epithelial cells of the nematodes are similar to human intestinal cells, making them a valuable in vivo model for preliminary studies of host–pathogen interactions [2425]. In contrast, multiple studies were performed on the Shigella–C. elegans model, and they were focused on the host response towards Shigella infection [21232627]. To our knowledge, this study is the first to analyse the Shigella response during C. elegans infection.
This study aimed to investigate both the host response and the pathogen response during the infection through dual RNA sequencing, where the transcriptomic data of both organisms are analysed during the bacterial infection. This study demonstrated that the infection by S. sonnei and S. flexneri can lead to different responses in the C. elegans host at early (10 minutes post-infection, or 10 mpi) and later (24 hours post-infection, or 24 hpi) stages of infection. The two Shigella also had different responses during C. elegans infection.
Methodology
Strains, plasmids and growth conditions
The C. elegans wild-type N2 Bristol strain was cultured on nematode growth medium (NGM) agar plates seeded with UV-killed Escherichia coli OP50 and maintained at 22 °C [28]. E. coli OP50 was used as the standardized food source for C. elegans in laboratory conditions and is therefore non-pathogenic [28]. S. sonnei ATCC 29930 and S. flexneri ATCC 12022 were obtained from the American Type Culture Collection (ATCC), and they are pathogenic towards C. elegans. Unless stated otherwise, all bacterial strains were grown on Luria-Bertani agar and broth (Himedia, India) and incubated overnight at 37 °C.
Exposure of C. elegans to bacterial strains, RNA extraction and sequencing
All infection assays were carried out as described previously [29]. The L4 stage of nematodes was exposed to S. sonnei and S. flexneri lawns. One hundred worms were harvested at two time points, 10 min (10 mpi) and 24 h post-infection (24 hpi). Tri-RNA (Favorgen), in combination with the Monarch Total RNA Miniprep kit (New England Biolabs, Massachusetts, USA), was used according to the manufacturer’s instructions to purify the RNA. In vitro S. sonnei 29930 cells grown on NGM agar overnight were also harvested for RNA extraction.
Transcriptome bioinformatic analyses
All RNA library samples were prepared as described [29] and sequenced on the Illumina Novaseq 6000 system (150 bp paired-end). The raw reads were trimmed using Trimmomatic v0.39 [30] and mapped to the C. elegans genome (NCBI RefSeq assembly GCF_000002985.6) using hisat2 v2.2.1 [31]. The unmapped reads were then mapped to S. sonnei ATCC 29930 or S. flexneri ATCC 12022 genome obtained from the ATCC Genome Portal [32]. The mapped reads were quantified using htseq-count [33]. The differentially expressed genes (DEGs) were determined with DESeq2 [34], using a threshold of FDR<0.05 and a log2fold change of ±1. Single orthologs between the two Shigella strains were obtained using ProteinOrtho [35], with the functional nucleotide sequences as input. Functional enrichment was performed using the online tool ShinyGo v0.80 [36].
Knockout mutants of S. sonnei ATCC 29930
Knockout mutants of the target genes in the S. sonnei ATCC 29930 wild type (WT) were constructed using homologous recombination as previously described [37] using the chloramphenicol resistance cassette as a selection marker. These genes are cspD and hok toxin genes. After electroporation, the bacteria were plated on agar containing 15 µg ml^−1^ of chloramphenicol for selection. Successful knockout mutants were verified by colony PCR.
Growth curve of mutants
The growth curves of the S. sonnei mutants and WT were obtained and compared. The overnight cultures of the bacteria were diluted to 0.05 OD and added to the microplate. The plate was incubated at 37 °C, and the OD 600 nm reading was obtained every 30 min. The maximum specific growth rate was determined using a growth curve fitting the Gompertz model, and the results from five replicates were analysed for any statistical significance using one-way ANOVA with the nonparametric Kruskal–Wallis test and Dunn’s multiple comparison tests.
Killing assay and bacterial colonization assay of nematodes
The virulence of the S. sonnei mutants was determined using the nematode killing assay and bacterial colonization assay as described previously [202938].
Results and discussion
Dual RNA sequencing of the Shigella-infected C. elegans
S. sonnei ATCC 29930 and S. flexneri ATCC 12022 do not contain the invasion plasmid required for virulence. However, the nematodes infected with these two Shigella had a significant decrease in lifespan compared to the non-pathogenic E. coli [29]. Additionally, there is a higher amount of Shigella obtained from the nematode gut after infection than E. coli OP50. A previous study had only investigated the bacterial response during C. elegans infection at 72 hpi. To further investigate the bacterial and host transcriptional response during infection, the total RNA from infected C. elegans at two time points (10 mpi and 24 hpi) was sequenced (Fig. 1). The reads were first mapped to C. elegans. Next, the unmapped reads were mapped to either S. sonnei or S. flexneri. An average of 66M reads was obtained from the S. sonnei–C. elegans samples, while 105M reads were obtained from the S. flexneri–C. elegans samples. More than 83% of reads remained after trimming and quality filtering, while the remaining reads did not pass quality filtering. For dual RNA-sequencing samples, more than 98% of the reads mapped to the C. elegans genome, while 0.02–0.47% of the reads mapped to the bacterial genome (Table S1, available in the online version of this article). The low amount of bacterial reads was expected due to the different genome sizes between the host and the bacteria [39]. All samples from the same conditions were clustered in the principal component assay plot (Fig. 1b, c). However, S. sonnei response at both time points was clustered closer to each other than S. flexneri (Fig. 1c), suggesting that their response at 24 hpi is similar to the beginning of infection.
C. elegans–Shigella host–pathogen infection samples. (a) A total of five conditions were used to collect RNA for sequencing (S. sonnei in vitro, S. sonnei-infected C. elegans 10 mpi, S. sonnei-infected C. elegans 24 hpi, S. flexneri-infected C. elegans 10 mpi and S. flexneri-infected C. elegans 24 hpi). The number of differentially expressed genes (DEGs) (P-value<0.05, fold change >2) are labelled between the conditions compared (orange indicates DEGs by C. elegans; black indicates DEGs by Shigella). Boxes indicate the samples used in the three different parts of the analysis. (b) The principal component analysis (PCA) plot of C. elegans during S. sonnei or S. flexneri infection at 10 mpi and 24 hpi. (c) PCA plot of the S. sonnei or S. flexneri response during C. elegans infection at 10 mpi and 24 hpi.
C. elegans has different responses towards S. sonnei or S. flexneri infection
S. sonnei-infected C. elegans differentially regulates 295 genes at the later infection stage (Fig. 2a), while S. flexneri-infected C. elegans differentially regulates 123 genes (Fig. 2b; DEGs listed in Tables S2 and S3, available in the online version of this article). Multiple pathways were enriched within the downregulated genes in C. elegans during the infection of S. sonnei (Fig. 2c) and S. flexneri (Fig. 2d). First, the infection of both S. sonnei and S. flexneri led to the downregulation of several genes involved in C. elegans defence response at 24 hpi. S. sonnei-infected C. elegans also downregulate genes involved in the stress response to cadmium ion (cdr-1, cdr-4 and pgp-3), response to xenobiotic stimulus (cyp-33C1, cyp-33C7, cyp-34A10, cyp-34A7 and nine cyp-35 genes) and sphingolipid metabolic process (asah-1, asah-2, asm-2, cgt-1 and gba-2). In contrast, S. flexneri-infected C. elegans downregulates genes involved in monovalent inorganic cation homeostasis (amt-1, nhx-2 and pept-1).
C. elegans response during S. sonnei or S. flexneri infection at 10 mpi and 24 hpi. (a) Volcano plot of DEGs in C. elegans during S. sonnei infection. (b) Volcano plot of DEGs in C. elegans during S. flexneri infection. (c) The enriched gene ontology (GO) biological processes identified among the downregulated DEGs in S. sonnei-infected nematodes. (d) The enriched GO biological processes identified among the downregulated DEGs in S. flexneri-infected nematodes.
Similar to this study, the nematode defence response is commonly enriched during bacterial infection [40]. For instance, the defence response genes were upregulated by the S. flexneri-infected nematodes compared to non-infected nematodes at the same time of infection [27]. However, our study demonstrated that the defence response was downregulated as the infection progressed from early to later stages. The downregulation of the host defence response, or host immune suppression, can be induced by the bacteria as a part of their pathogenesis repertoire, including Shigella [4143]. However, these are mainly mediated by the plasmid-encoded T3SS [4445]. As the studied Shigella strains do not have the pINV plasmid, this suggests that S. sonnei and S. flexneri may be suppressing these nematode defence response genes with other mechanisms.
During S. sonnei infection, C. elegans downregulated genes related to its xenobiotic response. The C. elegans xenobiotic response has been shown to interact with the immune response towards bacterial infection [46]. However, their specific role during S. sonnei infection is unclear. These genes may be induced at the early stages of infection and then downregulated after they are no longer needed. In addition, these genes are only downregulated in C. elegans during S. sonnei infection, suggesting that these may be responses against S. sonnei specifically. In contrast, S. flexneri-infected nematodes downregulate monovalent inorganic cation homeostasis genes, which are pept-1 and nhx-2. The peptide transporter PEPT-1 regulates the uptake of free fatty acids along with the sodium-proton exchanger NHX-2, and PEPT-1 loss leads to higher free fatty acid uptake [47]. While these proteins were not directly linked to the nematode response towards Shigella or any bacterial infection, they were downregulated in this study. This demonstrated that there are differences in C. elegans response towards S. sonnei or S. flexneri infection, which is similarly reported in other studies [4849].
S. sonnei upregulates biofilm formation and energy generation/conservation during in vivo growth in C. elegans
There are 460 S. sonnei genes differentially regulated after 10 min of infection, while 1265 genes were differentially regulated after 24 h of infection (DEGs listed in Tables S4 and S5). Functional enrichment analysis was performed on these genes to obtain the top ten pathways with the highest fold enrichment, and multiple pathways were commonly enriched at both time points of infection (Fig. 3). The submerged biofilm formation genes (ybgD, bssS, ychH and yfcV genes) are commonly upregulated at both time points (Fig. 3a, b). In contrast, the translation termination genes (frr, prfA and prfB), guanosine-containing compound biosynthetic process genes (guaA, guaB, relA, spot and gppA) and hydrogen sulphide metabolic process genes (cysD, cysI, cysJ and cysN) were downregulated by S. sonnei at both time points (Fig. 3c, d).
Enriched GO biological processes identified among the DEGs in S. sonnei during in vivo C. elegans infection at 10 mpi and 24 hpi. (a) Enriched within DEGs upregulated at 10 mpi. (b) Enriched within DEGs upregulated at 24 hpi. (c) Enriched within DEGs downregulated at 10 mpi. (d) Enriched within DEGs downregulated at 24 hpi. Red boxes indicate the common pathway enriched between two time points of infection.
Interestingly, an intermembrane transport protein gene, yebS, is upregulated at 10 mpi but downregulated at 24 hpi (which appears bold in Table S4 and S5). This gene contributes to membrane integrity in E. coli. However, its exact function is unknown [50]. As YebS has not been linked with bacterial pathogenesis before, its role in S. sonnei at early and later stages of C. elegans infection is currently unknown.
Biofilm formation genes were upregulated during both time points of infection (Fig. 3a, b). Biofilm formation is associated with host immune system evasion and survival during adverse conditions such as low pH in the gut [51]. Biofilm is also induced in S. flexneri by bile salts, indicating its role during gut infection [5255]. Other related genes, such as cell motility and biofilm regulatory genes, are also upregulated at 10 mpi, including bssR and bssS [56], adrA [57], glgS [58] and uspG [59], suggesting that fine-tuning biofilm formation during the early C. elegans infection is crucial for S. sonnei. Additionally, S. sonnei upregulates genes in pilus organization, cell projection organization and cell adhesion at 24 hpi, including multiple flagellar genes such as flg and fli genes. As biofilm formation requires cell aggregation and adhesion, cell projections such as pili and flagella help support the initial interactions between bacterial cells to form microcolonies [6061]. Thus, even though Shigella do not have functional flagella [62], the upregulation of biofilm regulatory genes may indirectly lead to increased flagellar gene expression.
S. sonnei also prioritizes energy generation or conservation during in vivo infection (Fig. 3). S. sonnei upregulates catabolic processes and energy derivation/generation and downregulates biosynthesis and DNA repair/replication during C. elegans infection. This includes the upregulation of polyamine catabolism at 10 mpi and the upregulation of rhamnose, colanic acid and other general catabolism at 24 hpi. S. sonnei also downregulates genes involved in hydrogen sulphide and guanosine-containing compound metabolism, sulphate and protein transport, as well as DNA replication and repair during C. elegans infection. Additionally, carbohydrate transport was upregulated at 24 hpi, providing nutrients and supporting bacterial growth in the host through increased glucose uptake [63]. Due to multiple challenges during C. elegans gut infection, such as insufficient nutrients, low pH and the host’s immune system [64], the generation and conservation of energy can support S. sonnei’s continued survival.
The membrane lipid metabolism is differentially regulated by both C. elegans and S. sonnei at 24 hpi simultaneously
The dual RNA sequencing and analysis allow the simultaneous observation of the host and pathogen response simultaneously. In this study, C. elegans downregulates sphingolipid metabolism genes (asah-1 and asah-2) (Fig. 2c), while S. sonnei upregulates ethanolamine metabolism genes (ccmL, eutE, eutN, eutP, eutQ, eutS, eutT, pduB and pudU) at 24 hpi (Fig. 3b). The simultaneous regulation of the membrane lipid metabolism in C. elegans and S. sonnei during 24 h of infection demonstrated that these membrane lipids have roles in both host defence and bacterial pathogenesis. Interestingly, the sphingolipid metabolism genes were not differentially regulated in S. flexneri-infected nematodes.
Sphingolipids are one of the membrane lipids in eukaryotes, and they have multiple roles in bacterial infection [65]. Both asah-1 and asah-2 genes are involved in sphingolipid catabolism, and the downregulation of sphingolipid catabolism improves host survival during bacterial infection [66]. Other sphingolipid metabolism genes provide protective activity towards P. aeruginosa and Enterococcus faecalis [67]. Thus, the downregulation of these genes in the nematodes suggests that this may be one of the protective responses against S. sonnei infection, but not S. flexneri, at 24 hpi.
Bacteria can either synthesize or exploit the host’s sphingolipids for pathogenesis [68]. Shigella relies on the host sphingolipids for pathogenesis, as S. flexneri invasion is reduced in host cells that are sphingolipid-deficient [69]. Phosphatidyl-ethanolamine, a precursor membrane lipid and a byproduct of sphingolipid catabolism, is important for host cell attachment [7071]. E. coli has been shown to alter the host phospholipid metabolism during infection [72], suggesting that the upregulation of ethanolamine metabolism by S. sonnei at 24 hpi is most likely helpful for pathogenesis.
S. sonnei upregulates acid resistance (AR) and biofilm genes compared to S. flexneri during C. elegans infection
S. sonnei and S. flexneri have 5417 and 5298 total annotated genes, respectively. There are 3658 genes with a single ortholog in each bacterial genome, while other genes were either specific to each strain or have multiple orthologs (Fig. 4a). A high number of these genes are hypothetical proteins. For the direct comparison of the transcriptomic responses of S. sonnei and S. flexneri during C. elegans infection, only the 3658 common genes were analysed. Between the responses of S. sonnei and S. flexneri during infection, 49 genes were differentially regulated at 10 mpi, while 65 genes were differentially regulated at 24 hpi (Fig. 4b, c; DEGs listed in Tables S6 and S7). At 10 mpi, S. sonnei upregulates genes involved in AR compared to S. flexneri, which are genes involved in the glutamate-dependent system (gadA, gadB and gadC) and the acid stress chaperones (hdeA and hdeB). At 24 hpi, S. sonnei also upregulates biofilm-related genes, including the two biofilm regulatory genes bssR and bssS.
Genomic and transcriptomic differences between S. sonnei ATCC 29930 and S. flexneri ATCC 12022 during C. elegans infection. (a) The number of orthologous genes between S. sonnei and S. flexneri was identified using ProteinOrtho. Black circles denote the genes with multiple orthologs in either strain, which are mostly hypothetical proteins. (b, c) Volcano plots of DEGs between S. sonnei and S. flexneri during (b) 10 mpi and (c) 24 hpi of C. elegans infection. (d) Heatmap of the 15 DEGs commonly identified from both analyses.
Shigella can persist in the acidic gut environment due to their glutamate decarboxylase-dependent AR system. The proteins GadA, GadB and GadC and the acid chaperones HdeAB play key roles in maintaining the intracellular pH at near-neutral conditions [7374]. This study suggested that S. sonnei upregulates its AR mechanism at the early stages of C. elegans infection, perhaps responding to the low pH environment earlier than S. flexneri. Additionally, biofilm is also involved in Shigella response towards the gut conditions, as biofilm formation can be triggered by bile acid in S. flexneri [53]. Most studies on AR [73,7578] and biofilm [7980] were performed on S. flexneri, while none were performed on S. sonnei. The upregulation of AR and biofilm in S. sonnei provides possible explanations for the differences in virulence between S. sonnei and S. flexneri. However, further investigation is required to elucidate the differences between these two species.
Knocking out CspD and a hok toxin in S. sonnei did not negatively affect the virulence of S. sonnei
There are 15 common genes upregulated during S. sonnei infection and expressed at significantly higher levels in S. sonnei than in S. flexneri (Fig. 4d). Two genes (cspD and hok toxin gene) were selected for further validation using gene knockout. The CspD protein is linked with persister formation, which protects the bacteria from host immune response [81]. The chromosome-encoded hok toxin prolongs the lag phase of bacteria to allow time for its stress response [82]. While deleting these genes did not affect their in vitro growth, there were also no differences in their virulence during C. elegans infection compared to the WT (Fig. S1). This suggests that while the hok toxin gene and cspD genes are differentially regulated within S. sonnei during the infection of C. elegans at 24 hpi, single gene deletion may not be sufficient to reduce the virulence of S. sonnei for a longer period.
Limitation of study
This study has multiple limitations. First, C. elegans is a soil nematode with an optimal temperature lower (20–25 °C) than the human body temperature (37 °C). Thus, the infection was performed at a lower temperature than is physiologically relevant. Nevertheless, as the nematodes demonstrated a significant decrease in lifespan after Shigella infection, this study provided some preliminary data on the Shigella–C. elegans infection model. Next, the number of reads that are mapped to the Shigella genome is relatively low compared to other bacterial transcriptomic studies. Thus, this study can only capture highly expressed bacterial genes. Nevertheless, this study provided some pathways and targets of interest involved in Shigella infection within C. elegans gut that are significantly differentially expressed during the infection.
Conclusion
In conclusion, this study provided insights into the response of C. elegans, S. sonnei and S. flexneri during 10 mpi and 24 hpi. C. elegans downregulated its defence response at a later stage of Shigella infection. *S. sonnei-*infected nematodes also downregulated their membrane lipid sphingolipid metabolism. S. sonnei also upregulates the membrane lipid precursor ethanolamine metabolism. S. sonnei also upregulates biofilm formation and prioritizes energy generation/conservation during C. elegans infection. Finally, the two Shigella species have slight transcriptomic response differences during C. elegans infection, as S. sonnei significantly upregulates its biofilm- and acid-resistance-related genes compared to S. flexneri. This study demonstrated that S. sonnei and S. flexneri have species-specific responses during pathogenesis that may affect their virulence.
supplementary material
10.1099/mgen.0.001339Uncited Supplementary Material 1.
10.1099/mgen.0.001339Uncited Supplementary Material 2.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Baker S The HC Recent insights into Shigella Curr Opin Infect Dis 20183144945410.1097/QCO.000000000000047530048255 PMC 6143181 · doi ↗ · pubmed ↗
- 2Thompson CN Duy PT Baker S The rising dominance of Shigella sonnei: an intercontinental shift in the etiology of bacillary dysentery P Lo S Negl Trop Dis 20159 e 000370810.1371/journal.pntd.000370826068698 PMC 4466244 · doi ↗ · pubmed ↗
- 3Anderson M Sansonetti PJ Marteyn BS Shigella diversity and changing landscape: insights for the twenty-first century Front Cell Infect Microbiol 2016610.3389/fcimb.2016.00045 PMC 483548627148494 · doi ↗ · pubmed ↗
- 4Banga Singh KK Ojha SC Deris ZZ Rahman RA A 9-year study of shigellosis in Northeast Malaysia: antimicrobial susceptibility and shifting species dominance Z Gesundh Wiss 20111923123610.1007/s 10389-010-0384-021654922 PMC 3089725 · doi ↗ · pubmed ↗
- 5Qiu S Xu X Yang C Wang J Liang B et al Shift in serotype distribution of Shigella species in China, 2003-2013 Clin Microbiol Infect 20152125210.1016/j.cmi.2014.10.01925658535 · doi ↗ · pubmed ↗
- 6Mao Y Cui E Bao C Liu Z Chen S et al Changing trends and serotype distribution of Shigella species in Beijing from 1994 to 2010 Gut Pathog 201352110.1186/1757-4749-5-2123919811 PMC 3750644 · doi ↗ · pubmed ↗
- 7Das SK Ahmed S Ferdous F Farzana FD Chisti MJ et al Etiological diversity of diarrhoeal disease in Bangladesh J Infect Dev Ctries 2013790090910.3855/jidc.300324334935 · doi ↗ · pubmed ↗
- 8Sousa MÂB Mendes EN Collares GB Péret-Filho LA Penna FJ et al Shigella in Brazilian children with acute diarrhoea: prevalence, antimicrobial resistance and virulence genes Mem Inst Oswaldo Cruz 2013108303510.1590/S 0074-0276201300010000523440111 PMC 3974317 · doi ↗ · pubmed ↗