Allelic frequency of msp2 and glurp genes in Plasmodium falciparum isolates from Awka, Anambra, Nigeria
Moses Ikegbunam, Abone Harrison, Chukwudi Egbuche, Nwasolu Obidi, Judith Mbamalu, Enyi Emmanuel, Offojebe Kosisochukwu, Mercy Ezeunala, Nzeukwu Chibumma, Ifeyinwa Onochie-Igbinedion, Joy Igwe, Joy Nnanna, Dorothy Ezeagwuna, Vincent Duru, Frances Nworji, Charles Esimone

TL;DR
This study analyzed the genetic diversity of malaria parasites in Nigeria, focusing on the msp2 and glurp genes to inform malaria control strategies.
Contribution
The study provides baseline data on allelic frequencies and genetic diversity of P. falciparum in Awka, Nigeria.
Findings
The 3D7 msp2 allele was predominant, with high polyclonal infection rates observed.
Twenty-one distinct msp2 alleles and nine glurp alleles were detected, showing significant genetic diversity.
The study found a high multiplicity of infection and heterozygosity, indicating a need for targeted malaria interventions.
Abstract
The genetic diversity of Plasmodium falciparum correlates with its pathogenicity, therefore design of evidence-based intervention strategies to eradicate malaria requires genetic diversity surveillance. This study characterised the allelic frequencies and genetic diversity of P. falciparum parasites isolated from Awka, Nigeria. Genomic DNA was extracted from 177 P. falciparum isolates and the polymorphic regions of the msp2 and glurp genes were genotyped by nested polymerase chain reaction (PCR). Two msp2 alleles (3D7 and FC27) were analysed. The 3D7 (93.55%) msp2 allelic family was predominant in msp2 positivie isolates. Polyclonal msp2 infection was observed in 24 (38.71%) isolates. Twenty-one distinct msp2 alleles were detected, with fragment sizes ranging from 200 bp to 1200 bp. The 300 bp allelic fragment (26.83%) was predominant for the 3D7 allele, while the 400 bp allelic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
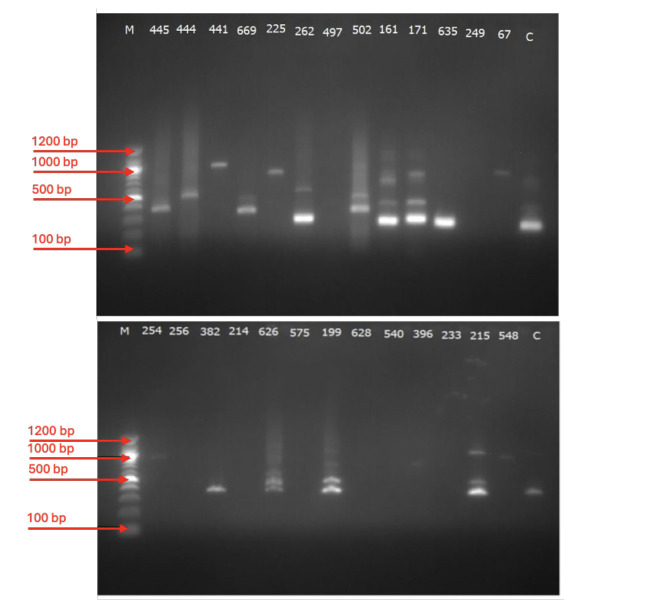 Figure 1
Figure 1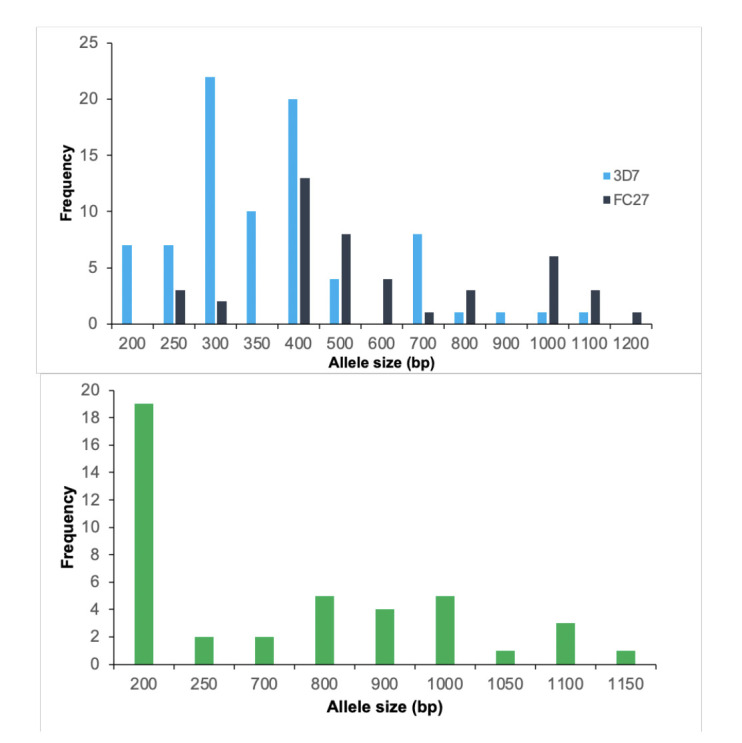 Figure 2
Figure 2| Gene | Primers |
|---|---|
|
| F: 5'-ATGAAGGTAATTAAAACATTGTCTATTATA-3' |
| R: 5'-TTATATGAATATGGCAAAAGATAAAACAAG-3' | |
|
| F: 5'-GCAAATGAAGGTTCTAATACTAATAG-3' |
| R: 5'-GCTTTGGGTCCTTCTTCAGTTGATTC-3' | |
|
| F: 5'-GCAGAAAGTAAGCCTCTACTGGTGCT-3' |
| F: 5'-GATTGTTTCGGCATTATTATGA-3' | |
|
| F: 5'-TGAATTTGAAGATGTTCACACTGAAC-3' |
| R1: 5'-ACATGCAAGTGTTGATCCTGAAG-3' | |
|
| F: 5'-TGTTCACACTGAACAATTAGATTTAGATCA-3' |
| R2: 5'-TGTAGGTACCACGGGTTCTTGTGG-3' |
| Markers | Frequency (%) | Allele size (bp) | Total No of Alleles | Distinct alleles |
|---|---|---|---|---|
|
| 28 (45.16%) | 250 -1200 | 44 | 10 |
|
| 58 (93.55%) | 200 -1100 | 82 | 11 |
|
| 24 (38.71%) | |||
|
| 69 (38.98%) | 200-1150 | 79 | 9 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
Topicsthermodynamics and calorimetric analyses
Introduction
Nigeria has the highest burden of malaria globally. It is a major public health concern in Nigeria with about 200,000 deaths from the disease annually, the main victims being children <5 yrs and pregnant women [1]. Plasmodium falciparum is the deadliest and most prevalent plasmodium species associated with malaria infection in sub-Saharan Africa [1,2]. The extensive genetic diversity of P. falciparum strains poses a challenge to efforts to eradicate malaria and possibly contributes to malaria pathology by suppressing acquired immunity and prompting the emergence of drug resistance and insecticide resistance variants [3,4,5]. Therefore, genomic surveillance of the parasite is essential to develop an effective control strategy to eradicate malaria in Nigeria.
An established method to survey the parasite is by targeted genotyping of merozoite surface protein (msp-2) and glutamate-rich protein (glurp) genes as they contain polymorphic regions that are used as markers for defining genetic variation in P. falciparum malaria and determining the multiplicity of infection (MoI) [4,6,7]. The msp-2 glycoprotein is an asexual blood stage antigen that consists of 5 polymorphic blocks with block 3 being the most polymorphic. The two main allelic families of msp-2, FC27 and 3D7, are based on the polymorphic regions of the central repeat sequences [8,9]. The glurp protein is an antigen expressed in both the pre-erythrocytic and erythrocytic stages of the parasite, as well as on the surface of newly released merozoites. This antigen consists of three regions, with the immunodominant C-terminal repetitive region (R2) being the most polymorphic [10]. The msp-2 and glurp proteins are also considered promising candidate antigens for the development of a malaria vaccine as they are targeted by cytophilic antibodies and are associated with natural immune protection against clinical malaria [11,12],
Understanding the genetic diversity of P. falciparum in different geographical regions of Nigeria is essential for developing new and effective malaria control interventions. Currently, there is limited information on the genetic diversity and multiplicity of P. falciparum infection in southeast Nigeria. This study aimed to evaluate the allelic frequency of msp2 and glurp genes in P. falciparum parasites isolated in Awka, an urban city in southeast Nigeria. This research will contribute information on disease pathogenesis and immunity acquisition in Nigeria. It also provides information that is benefcial for the development of an effective malaria vaccine.
Materials and Methods
This survey was conducted to assess the allelic frequency of P. falciparum msp2 and glurp genes. A total of 179 participants, aged 6 months to 68 yrs, were recruited between February 2019 and January 2020. Participants were included if they presented with malaria-related symptoms (e.g., fever, chills, headache) at Chukwuemeka Odumegwu Ojukwu University Teaching Hospital, Awka, Anambra State, Nigeria. The recruitment was based on clinical suspicion of malaria, and participants were screened using a rapid diagnostic test (RDT) for malaria prior to enrolment. Written informed consent was obtained from all participants or their legal guardians before inclusion in the study.
Study site and population
The study was conducted in Awka, Anambra State, Nigeria, a region with a tropical climate characterised by a wet season from April to October and a dry season from November to March. The area experiences an average annual rainfall of 1,200 mm and a temperature range of 25–32°C. Malaria transmission in this region is perennial, with peaks during the rainy season. Control measures include the use of insecticide-treated nets (ITNs) and intermittent preventive treatment for pregnant women (IPTp).
Participants were recruited from outpatient clinics, with inclusion criteria as follows: Age between 4 months and 60 years, presentation with clinical signs and symptoms suggestive of malaria and positive RDT for malaria prior to study enrolment. Exclusion criteria included participants on malaria treatment within the last two weeks or those unwilling to provide informed consent.
Sample collection, preparation and parasite detection
Venous blood samples (2 mL) were collected into EDTA tubes from each participant. RDT was performed using the SD Bioline Malaria Ag Pf/Pan (South Korea), which detects P. falciparum and non-falciparum species. Samples positive by RDT were preserved at −20°C until genomic DNA extraction. DNA extraction was performed using the Quick-DNA Miniprep kit (Zymo Research, USA) following the manufacturer’s protocol.
Molecular genotyping of P. falciparum msp2 and glurp genes
The polymorphic regions of the msp2 and glurp genes were amplified in a nested PCR reaction. The primary PCR conditions for both the msp2 and glurp genes were an initial denaturation step of 95 °C for 5 min followed by 30 cycles of 95 °C for 1 min, 54 °C for 1 min, 72 °C for 1 min, and a final extension of 72 °C for 5 min. The nested PCR parameters for the glurp gene were identical to the primary reaction; only the annealing temperature was adjusted to 59 °C. For the msp2 gene, the nested PCR conditions were initial denaturation at 94°C for 5 min, followed by 30 cycles at 94 °C for 10 s, 57 °C for 30 s, and 72 °C for 40 s. The final cycle had a prolonged extension at 72 °C for 3 min. The primers targeting the RII region of glurp [13] and the 3D7 and FC27 regions of msp2 [14] are shown in Table 1. The PCR products were separated on a 1.5% agarose gel, stained with ethidium bromide, and visualised using a UV transilluminator (Vilber, France).
Heterozygosity and multiplicity of infection
The expected heterozygosity index (He) was calculated using the formula:
He=[nn−1][1−∑Pi2]
Where n = number of isolates analysed and Pi = the frequency of the i^th^ allele in the population. The multiplicity of infection (MoI) was calculated by dividing the total number of fragments detected in an antigenic marker by the number of samples positive for that same marker. Samples where only one genotype was detected per allelic family were monoclonal, while samples with two or more genotypes per allelic family were polyclonal P. falciparum infections.
Data analysis
Heterozygosity index (He) and multiplicity of infection (MoI) were calculated. The presence of polyclonal infections was determined based on the detection of multiple alleles within the same sample.
Ethical approval
The Chukwuemeka Odumegwu Ojukwu University Teaching Hospital's Ethics Review Board granted ethical approval for the study. (COOUTH/CMAC/ETH.C/Vol.1/0035). Informed consent was obtained from the parent or legal guardian of each child before being included in the study.
Results
The study population consisted of 53.03% female and 46.97% male patients aged 6 months to 68 years The mean age of participants was 18.67 ± 0.32 yrs. A total of 179 blood samples were analysed for Plasmodium spp. using a malaria RDT kit which confirmed the presence of P. falciparum in 177 (99%) samples.
Genetic diversity of P. falciparum infection
PCR amplification was successful in 35.02% (62/177) of the isolates for the msp2 gene and 38.98% (69/177) of the isolates for the glurp gene (Table 2). For msp2, the Fc27 allele had a frequency of 45.16% (28/62) while the predominant 3D7 allele had a frequency of 93.55% (58/62). Monoclonal infections were identified in 38 isolates (61.29%), with 34 isolates (54.84%) positive for the 3D7 allele and 4 isolates (6.45%) positive for the FC27 allele. Polyclonal infections (FC27+IC3D7) were detected in 24 isolates (38.71%). The RII repeat region of the glurp gene was detected in all 69 isolates. Representative gel pictures are presented in Figures 1.
Representative Gel images of samples showing different alleles for 3D7 (top) and FC27 (bottom).
Allelic frequency of msp2 and glurp genes
The allelic genotyping data revealed the polymorphic nature of P. falciparum parasites in Awka. In the msp2 and glurp genes, different allelic types were identified. Alleles of msp2 and glurp were classified according to the size of the amplified PCR bands. Twenty-one alleles were detected in P. falciparum isolates positive for the msp2 gene. This includes 10 FC27 alleles and 11 3D7 alleles with 200–1200 bp fragment sizes. The 300 bp allelic fragment (26.83%) was predominant for the 3D7 allele, while the 400 bp allelic fragment (29.55%) was predominant for the FC27 allele (Figure 2). PCR amplification for the RII repeat region of the glurp gene was successful in 69 isolates. Nine alleles with fragment sizes ranging from 200 to 1150 bp were identified. The most prevalent allelic fragment was 200 bp (45.24%), while both the 1050 bp and 1150 bp allelic fragments were the least prevalent (2.38%) (Figure 2).
Prevalence of P. falciparum 3D7 and FC27 msp2 alleles (top) and glurp alleles (bottom).
Multiplicity of infection (MoI)
The MoI for the msp2 gene (MoI = 2.03) was high as was the heterozygosity (He = 0.34). The MoI and He for the glurp gene were 0.45 and 0.98, respectively.
Discussion
Nigeria is a malaria-endemic country, and an increase in the genetic diversity of P. falciparum strains could lead to more complex infections and the emergence of more virulent or drug-resistant variants, endangering efforts to eradicate the disease [15]. Genetic diversity and polymorphism are key in the acquisition of anti-malaria parasite immunity [16,17]. Therefore, determining the frequency of Plasmodium genotypes in different geographical locations would facilitate the development of effective control strategies. Polymorphic markers in P. falciparum isolates were used to examine the genetic diversity and complexity of parasite populations in patients with symptomatic malaria at the Chukwuemeka Odumegwu Ojukwu Teaching Hospital, Awka, Nigeria.
In this study, allele-specific genotyping of msp2 in P. falciparum isolates reveals high allelic diversity in Awka, Nigeria. The high malaria transmission rate, the incidence of mixed illnesses, and the subsequent exposure of locals to mosquito bites may be responsible for this trend in the research area. For msp2, the 3D7 alleles were predominant, with a 93.54% occurrence. This data is consistent with studies from Kaduna [18], Ibadan [19], Anambra [7], and north-central Nigeria [20, 21] that reported a high prevalence of the 3D7 family, in symptomatic patients, but the results contrast with a study by Ojurongbe et al. [22] in Osogbo, Nigeria, which reported the FC27 allele as the predominant allele. The result is in agreement with other studies conducted in sub-Saharan Africa [23,24], South America [16], and Asia [17,25]. The results suggest that the frequency of 3D7 alleles strongly correlates with symptomatic malaria. The result is conflicting as the 3D7 allele is associated with asymptomatic malaria infections, and it is thought to offer protection against clinical disease [26]. The contrasting observation from various studies indicates a need to understand the influence of human genetic factors on the antigenicity of msp2 alleles as the variations observed in different studies may be attributed to immune selection pressures.
Among the msp2 positive P. falciparum isolates, polyclonal infections were observed in 38.71% of participants. Complex infections marked by multiclonality impacts drug efficacy, severity of disease and the population diversity of the parasite [4]. Polyclonal infections are associated with low levels of protective malaria antibodies, increased prevalence of drug-resistant parasites and possibility of recrudescence [27,28]. More importantly, malaria vaccine efficacy studies on the msp2 gene have shown that vaccination with only one msp2 variant induces an allele-specific response. In one study involving a vaccine that comprised the 3D7 allelic family, vaccinated patients showed an increase in morbidity associated with the FC27 alleles [29]. Subsequent studies have shown that chimeric vaccines [30] and vaccines containing both msp2 allelic families [31] induce a strain-transcending immune response. This necessitates that the malaria vaccine design incorporates both allelic variants of the msp2 gene to account for the increase in polyclonal infections observed in this study.
The high genetic diversity for msp2 is consistent with observations in malaria-endemic regions [4]. The high genetic diversity may correlate with the rate of polyclonal infections and the intensity of malaria transmission in the region [4]. The multiplicity of P. falciparum infections (MoI) for msp2 reported in this study is similar to studies from Nnewi [7] and Ibadan [32] in Nigeria. However, other studies from southwest Nigeria [33,34,35] and Pahang, Malaysia [36] reported a lower MoI, while a study on children living close to a lake in Taabo, Côte d’Ivoire, reported a higher MoI. The multiplicity of infection (MoI), i.e., the number of different P. falciparum strains co-infecting a single host in many malaria-endemic areas is a common feature and has been reported to vary with age, parasite density, immune status, epidemiological settings, and transmission intensity [37]. The MoI observed in this study could indicate high transmission levels of the parasite.
The RII region of the glurp was also shown to have a high prevalence in the study population, with 38.98% occurrence. The study by Ullah et al. [6] showed a higher prevalence of 70% in Pakistan, consistent with another study conducted in south-western Nigeria [32]. Furthermore, a study in Osogbo, Nigeria, reported that the genetic diversity of the R2 polymorphic region of the glurp gene remained diverse despite the implementation of the artemisinin-based combination therapy ACT therapy in the study area [38]. The glurp gene is an antigen of P. falciparum that is highly conserved, present in all stages of the malaria parasite and associated with clinical immunity. These attributes make it a promising biomarker for diagnosis and the development of vaccines against malaria.
Conclusions
The present study shows that there is a high level of polyclonal P. falciparum infections in the population. The P. falciparum parasites harbour multiple gene alleles with high MoI. This indicates the extensive genetic diversity of P. falciparum infection in the study area. The data provides important baseline information to guide malaria control and elimination strategies in the study area and across Nigeria.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1World Health Organization: World malaria report 2023 Geneva World Health Organizationhttps://tinyurl.-com/4h 4jd 9nx (accessed 23 January 2025).
- 2Alemayehu A Biology and epidemiology of Plasmodium falciparum and Plasmodium vivax gametocyte carriage: implication for malaria control and elimination.Parasite Epidemiol. Control 202321 e 00295 Doi: 10.1016/j.-parepi.2023.e 0029536950502 PMC 10025134 · doi ↗ · pubmed ↗
- 3Abukari Z Okonu R Nyarko SB Aminata C Lo The diversity, multiplicity of infection and population structure of P. falciparum parasites circulating in asymptomatic carriers living in high and low malaria transmission settings of Ghana.Genes 201910434 Doi: 10.3390/genes 1006043431181699 PMC 6628376 · doi ↗ · pubmed ↗
- 4Opute AO Akinkunmi JA Funsho AO Obaniyi AK Genetic diversity of Plasmodium falciparum isolates in Nigeria. A review.Egypt. J. Med. Hum. Genet.202223129 Doi: 10.1186/s 43042-022-00340-7 · doi ↗
- 5Ahouidi A Ali M Almagro-Garcia J Amambua-Ngwa A An open dataset of Plasmodium falciparum genome variation in 7,000 worldwide samples.Wellcome Open Res.2021642 Doi: 10.12688/wellcomeopenres.16168.233824913 PMC 8008441 · doi ↗ · pubmed ↗
- 6Ullah I Khan A Israr M Shah M Genomic miscellany and allelic frequencies of Plasmodium falciparum msp-1, msp-2 and glurp in parasite isolates.P Lo S One 202217 e 0264654 Doi: 10.1371/journal.pone.026465435259187 PMC 8903261 · doi ↗ · pubmed ↗
- 7Ikegbunam MN Anagu LO Duru C Nworu CS Genetic diversity and allelic frequency of antigenic markers in Plasmodium falciparum isolates from Nnewi district in Nigeria.J. Infect. Dev. Ctries 202216557563 Doi: 10.3855/jidc.1481535404863 · doi ↗ · pubmed ↗
- 8Beeson JG Drew DR Boyle MJ Feng G Merozoite surface proteins in red blood cell invasion, immunity and vaccines against malaria.FEMS Microbiol. Rev.20164034372 Doi: 10.1093/femsre/fuw 00126833236 PMC 4852283 · doi ↗ · pubmed ↗