Identification and phylogenetic analysis of marine sponges in the Jordanian Gulf of Aqaba using DNA barcoding
Zeinab Arabeyyat, Mais Sweiss, Abdalmajeed Alajlouni, Neda'a Al-Ajlouni, Marwan Mahmoud, Sura Shartooh, Farah Alsoqi, Maysoon Kteifan

TL;DR
This study uses DNA barcoding to identify and classify marine sponges in the Jordanian Gulf of Aqaba, revealing five sponge genera and an unidentified one.
Contribution
This is the first morpho-molecular study of marine sponges in the Jordanian Gulf of Aqaba using DNA barcoding markers.
Findings
Five sponge genera were identified: Axinella, Negombata, Siphonochalina, Diacarnus, and an unidentified genus in Haplosclerida.
DNA barcoding and phylogenetic analysis provided a reliable method for sponge identification due to limited morphological traits.
This study establishes a new protocol for sponge identification at the genus level in the region.
Abstract
Sponges (Porifera) are the largest biomass component of coral reefs benthic fauna among marine organisms and are very morphologically diverse. In the present work, we aimed to identify marine sponges in the Jordanian Gulf of Aqaba using the partial 18S rRNA and the 28S rRNA genes as DNA barcoding markers. A total of nine morphologically different marine sponge samples from 6.6m to approximately 22m depth were collected. Sponge fragments were frozen at −80 °C prior to DNA extraction. The sponge's DNA was extracted using a commercial kit and subjected directly to PCR amplification for the 18S rRNA and 28S rRNA genes. The DNA sequences were analyzed using the Basic Local Alignment Search Tool (BLAST) to determine the sponge's identity, and phylogenetic trees were constructed to clarify the relationship among the samples. The results obtained revealed the presence of the following genera:…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
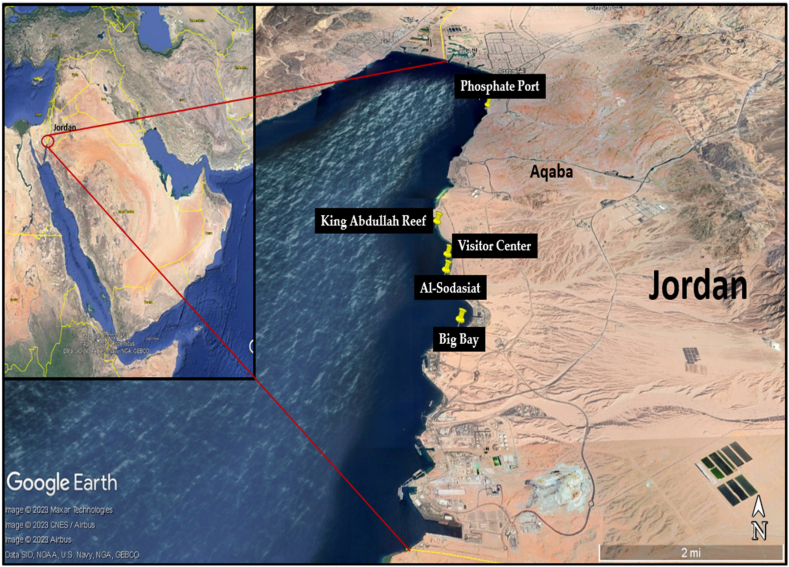 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3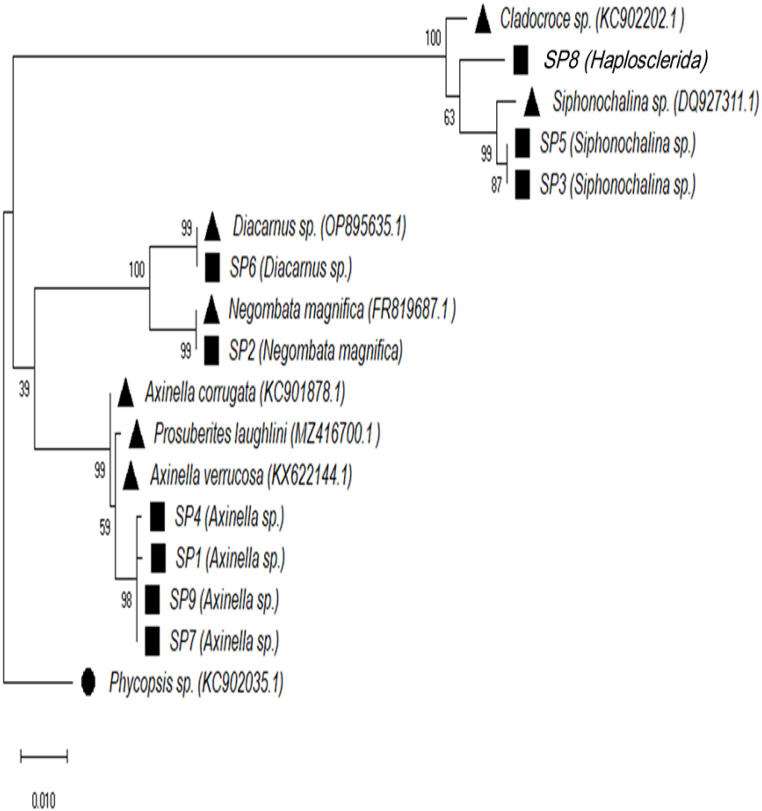 Figure 4
Figure 4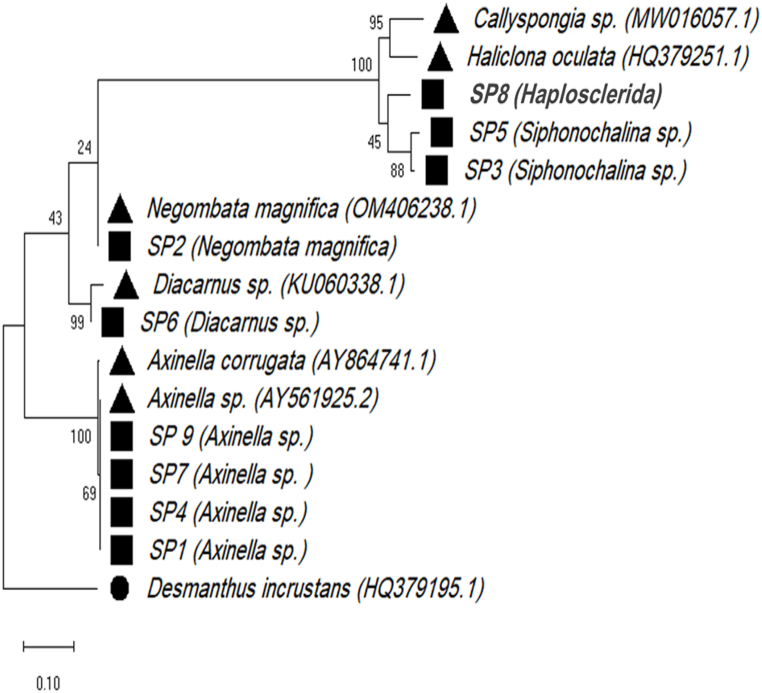 Figure 5
Figure 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMarine Sponges and Natural Products · Marine Ecology and Invasive Species · Aquaculture disease management and microbiota
Introduction
1
Coral reefs are important habitats with the most diverse of all marine ecosystems. They provide a home for at least a fourth part of marine fauna, including sponges, mollusks, echinoderms, fishes, cnidarians, crustaceans, and others [1]. Coral reefs are known worldwide as fragile ecosystems that are exposed to many threats such as climate change, ocean acidification, illegal fishing, and harmful land use practices [2]. The coral reefs in the northern Red Sea are also vulnerable to human impacts such as tourism, shipping, pollution, and port activities [3].
One of the largest biomass components of coral reefs benthic fauna is sponges in the Caribbean sponges [4]. While sponges (Phylum Porifera) are essential for the functioning of the coral reef ecosystem [5], relatively little is known about sponge species diversity in the Red Sea [6]. Our current knowledge of the Red Sea sponge is based on Wooster and his colleagues [7] and Erpenbeck and his colleagues [8,9], where it harbors a diverse understudied community of sponge species with around 261 valid sponge species (representing 114 genera) in the Red Sea [7]. However, the Jordanian Gulf of Aqaba is still considered an underexplored Gulf region. This gulf is part of a semi-enclosed basin that is separated from the Red Sea at the Straits of Tiran, spans 27 km in length, and is known as one of the unique aquatic ecosystems that has one of the world's richest coral communities [10].
The Red Sea is known as a high biodiversity and endemism region [7]. However, little is still known about the biodiversity of Red Sea sponges [8], especially regarding data on their abundance, coverage, and species composition that remain scarce [6]. This limitation in data is due to limited taxonomic knowledge, the cryptic growth forms of sponges, and the challenges of identifying them in the field [11].
Morphological identification of sponges based only on spicule identification is a difficult task, as the morphology of sponge spicules can be simple, complex, or even absent [12]. Therefore, genetic analysis of sponges can shed light on their unique evolution, and sponges may prove to be ideal model organisms for addressing questions of evolutionary and ecological genetics [6]. Furthermore, molecular tools enable the assessment of phylogenetic relationships in sponges, ranging from intra-species to phylum-level relationships [8], despite the absence of a distinct barcoding gap and the lack of a genetic distance threshold-based species concept for sponges [8,13].
Marine sponges have unique phylogenetic relationships and genetic diversity that can be effectively explored through the combined use of morphological analysis and molecular barcoding. In the present study we therefore assessed the morphological and molecular identification of sponges using two nuclear ribosomal rRNA genes, the large subunit 28S rRNA and the small subunit 18S rRNA. These two markers were utilized for molecular barcoding and phylogenetic analysis. Additionally, to our knowledge, no prior study has focused on clarifying sponge with morphological and molecular identification of sponges in the Jordanian Gulf of Aqaba.
Materials and methods
2
Sponge samples collection
2.1
Sponge samples were collected from an inshore area in Jordan's marine environment by SCUBA diving. The Jordanian Gulf of Aqaba is known for its small coral reef ecosystems that cover only 4 km^2^ in total along the 27 km of the entire coastline. Nine sponge samples were collected from five inshore reefs (Fig. 1) along the short coast of the Jordanian Gulf of Aqaba (29°31′36.01"N 35°00′28.01"E). Fragments of sponge individuals were collected at a depth range of 6.6–22 m. The geographic coordinates of the five sampling sites are shown in Table 1. Sponges were collected during December 2022. The temperature of the water was around 25 °C, and all sponges were emergent species living in an area that was exposed to light. Sponge fragments were collected and placed separately into a sterile plastic bag; the bags were then stored on ice for transport to the laboratory. In the laboratory, all samples were immediately placed at −80 °C prior to further analysis.Fig. 1. Collection sites of samples used in this study. Map created with Google Earth Pro (Google LLC., Mountain View, CA, USA).Fig. 1. Table 1Geographic coordinates and depth of sponge sampling sites.Table 1. Sampling siteSample IDsDepth (m)CoordinatesBig BaySP117.829°24′18.97"N/34°58′23.79"ESP218SP313.3Visitor CenterSP46.629°25′36.41"N/34°58′17.14"EPhosphate PortSP516.429°30′9.48"N/34°59′30.81"EKing Abdullah ReefSP616.529°26′23.34"N/34°58′8.80"ESP716.7SP822Al-SodasiatSP99.529°25′16.42"N/34°58′14.45"E
Morphology of sponge samples
2.2
The morphological identification methods using the external morphology and spicules of sponges were used to identify collected sponges in this study based on the descriptions of various sponges from different regions available in the literature [7,[14], [15], [16], [17]]. Each sponge species has one or more types of spicules, each with a specific morphology [12]. In the present study, eight sponge samples out of the original nine samples were identified by determining the structure of their spicules (SP7 is not included here, but was tested for molecular identification). Spicule analysis was carried out using the technique described in Hooper (2000) using a rapid tissue digestion in bleach [18]. Sponge fragments of approximately 5–10 mm were placed in 15 ml conical tubes and digested with 5 ml active bleach (Spartan©, Jordan) and incubated until all tissue dissolved. Bleach was diluted with water and then replaced with ethanol. A clean spicule suspension was aspirated and pipetted onto a microscope slide. Slides were analyzed under magnifications of 50–400x on an inverted microscope (AX10 Vert.A1, Zeiss, Germany).
Total genomic DNA extraction and PCR amplification
2.3
Total genomic DNA of nine sponge samples was extracted using the G-spin™ Genomic DNA Extraction Kit (iNtRON Biotechnology, South Korea) following the manufacturer's protocol. The quality of the DNA extracts was determined by 1.5 % agarose gel electrophoresis. The 28S rRNA partial gene was amplified using a sponge universal forward primer C1’ (5′-ACCCGCTGAATTTAAGCAT-3′), and reverse primer D2’ (5′-TCCGTGTTTCAAGACGGG-3′) [19]. Thermocycler conditions for the C1′-D2′ primer pair were as follows: the first cycle is 94 °C for 4 min, annealing at 57 °C for 2 min, and extension at 72 °C for 2 min, followed by 30 cycles, each consisting of 1 min at 94 °C, 1 min at the annealing temperature, and 1 min at 72 °C. Followed by a final extension step of 4 min at 72 °C [19]. The 18S rRNA gene fragment was amplified using the forward primer SS5 (5′-GGTTGATCCTGCCAGTAGTCATATGCTTG-3′), and reverse primer SS3 (5′-GATCCTTCCGCAGGTTCACCTACGGAAACC-3′) [20]. The thermocycler conditions for the 18S-rRNA primers were as follows: the first cycle is 94 °C for 2:30 min, annealing at 56 °C for 1 min, and extension at 72 °C for 2 min, followed by 29 cycles, each consisting of 1 min at 94 °C, annealing at 56 °C for 1 min, and 2:30 min at 72 °C. Followed by a final extension step of 4 min at 72 °C ended the reaction [21]. PCR products were analyzed on 1 % agarose gels using RedSafe™ nucleic acid staining solution (iNtRON Biotechnology, South Korea). The expected PCR product sizes were 800–900 bp for 28S rRNA (C1’ – D2′ primers) and ∼1800 bp for 18S rRNA with (SS5 – SS3 primers).
Sequencing and phylogenetic analysis of amplified PCR products
2.4
Direct sequencing of the PCR products for the 18S and 28S rRNA genes of the sponge was conducted using the Sanger sequencing service from Macrogen (Macrogen Inc., Korea). Sequences were edited by removing low-quality sequences at the beginning and the end of the chromatogram. Edited sequences were used for similarity search using the BLASTn tool at the National Center for Biotechnology Information (NCBI) [22]. GenBank accession numbers for sponges (n = 9) collected in this study were submitted to GenBank (NCBI) and have the following accession numbers: 18S rRNA (accession no. OR532378–OR532386) and 28S rRNA (accession no. OR532387–OR532395). To clarify the phylogenetic relationship among the collected samples and the sponge genera in GenBank, sequences were aligned using the ClustalW algorithm, with a sequence length after trimming of 991 nt for the 18S rRNA and 735 nt for the 28S rRNA. The bootstrapping value was 10000 replicates. The maximum likelihood phylogenetic tree was constructed based on the Tamura-Nei model [23] using MEGA X [24]. Phycopsis sp. (KC902035.1) and Desmanthus incrustans (HQ379195.1) were outgroups for the 18S rRNA and 28S rRNA phylogenetic trees, respectively.
Results
3
Identification of sponges
3.1
Based on the external characters [14], SP1 grows as a massive sponge; SP2 is like an erect and branched sponge; SP3 includes cups and thin cylindrical branches morphologies; SP4 is like a composite massive sponge; SP5 has cup- and barrel-shaped forms; SP6 is like a thick, erect, branched sponge; SP7 has a thick, erect, and rounded branch; SP8 has a massively encrusting shape. While SP9 has a flabellate shape.
For the morphologies of spicules, only eight sponge spicule samples were analyzed for sponge identification using the inverted microscope (only sample SP7 was missing when analyzed). Examples of the spicule morphology of the preparations analyzed are presented in Fig. 2. The shape and sizes of spicules in each sponge are variable. Based on the presence of only style spicules [16], we concluded that samples SP1, SP4, and SP9 belong to the same genus, Axinella. The spicules from sample SP2 are oxea, strongyle, and sanidaster spicules that match the description of Negombata magnifica [25]. Spicules from sample SP3 are spheroxyasters, acanthoxea spicules, styles, and tripod spicules. While spicules from sample SP5 are oxea and strongyle spicules from the Siphonochalina genus [26], spicules from sample SP6 are oxea spicules of the Diacarnus genus [25], and spicules from sample SP8 were only style spicules with one end pointed and the other rounded. Based only on morphological characters, it was not possible to determine the genera for samples SP3 and SP8.Fig. 2. Representative spicules of sponge samples SP1 – SP9. SP7 was not available in this analysis.Fig. 2
For molecular identification, a total of nine 18S and nine 28S sequences were successfully obtained from nine samples (see Fig. 3 for sponge samples). The partial nuclear 18S and 28S rRNA sequences have been analyzed and NCBI BLASTN [22] was used to find similar sequences.Fig. 3. Underwater photographs of all sponges sampled in the present study. Sponges photographs were collected by scuba divers. These photographs have been color corrected using an algorithm developed by Nikolaj Bech Andersen [31].Fig. 3
The novelty of this study is that it is the first morphological and molecular identification of marine sponges in the Jordanian Gulf of Aqaba, which has yet to be studied much by previous researchers. In the current study, the morphology characteristics support results of the molecular analysis and match those of SP1 (Axinella sp.), SP2 (Negombata magnifica), SP4 (Axinella sp.), SP5 (Siphonochalina sp.), SP6 (Diacarnus sp.), and SP9 (Axinella sp.) [[27], [28], [29], [30]]. However, it was difficult to identify the collected sponges to the species level, except for SP2.
Based on the sequencing analysis, all sequences from sponge samples were identified and classified as belonging to the following five genera: Axinella, Negombata, Siphonochalina, Diacarnus, and an unidentified genus within the order Haplosclerida, with identity percentages ranging from 98.29 to 100. The genus Axinella was predominant in 4 out of 5 sampling sites. Identification at the species level was possible for only one sample (SP2). The samples were identified at the genus level because they had identical sequences (the markers did not vary), and it was not possible to determine the species. For SP8, the highest identity based on the 28S rRNA sequence was with Haliclona oculata HQ379251.1 and Callyspongia sp. 1 JV-2020 MW016057.1; but the low identity (around 86 %) suggests that SP8 does not belong to these genera. However, the highest identity based on the 18S RNA was around 98.29 % for the Cladocroce sp. 0CDN9562-C and Calyx sp. NIWAKD1132, supporting its classification within these genera. Therefore, we recommend classifying SP8 within a genus of the order Haplosclerida. Additionally, some sequences of the 28S rRNA gene have very low identity with sequences available on GenBank (such as SP3, SP5, and SP8), and that could be due to the lack of information in the GenBank database. At 86.63 % identity, SP3 has the lowest identity for the 18S rRNA sequences (Table 2).Table 2BLAST analysis of PCR amplicons from marine sponge isolates according the highest similarity to 18S and 28S rRNA sequences in the GenBank nucleotide sequence database.Table 2. Sponge IDsLocusSize (nt)Closet organism in GenBank & NCBI accession no.Max scoreQuery (%)Identity (%)E-valueGenBank accession no.bSP128S700aAxinella sp. (AY561925.2)12421001000.00OR532387aAxinella corrugate (AY864741.1)1214100990.0018S950aAxinella verrucosa (KX622144.1)162010099.110.00OR532378Axinella corrugate (KC901907)162010099.110.00aAxinella corrugate (KC901878.1)1615100990.00aProsuberites laughlini (MZ416700.1)1615100990.00SP228S763aNegombata magnifica (OM406238.1)9941001000.00OR532388Negombata sp. (OM406250.1)98310099.630.0018S1050aNegombata magnifica (FR819687.1)193610099.900.00OR532379Neopodospongia cf. normani (KC902112.1)188610099.050.00Diacarnus sp. (OP895635.1)184510098.380.00SP328S700aCallyspongia sp. (MW016057.1)7679986.630.00OR532389Haplosclerida sp. (MW016166.1)7619986.630.00aHaliclona oculate (HQ379251.1)72810085.820.0018S1000aSiphonochalina sp. (DQ927311.1)181410099.40.00OR532380aCladocroce sp. (KC902202.1)a175110098.30.00Calyx sp. (DQ927313.1)175110098.30.00SP428S722aAxinella sp. (AY561925.2)12511001000.00OR532390aAxinella corrugate (AY864741.1)122310099.260.0018S1000aAxinella verrucosa (KX622144.1)180310099.20.00OR532381Axinella corrugate (KC901907.1)180310099.10.00aProsuberites laughlini (EF654529.1)179710099.10.00SP528S800aCallyspongia sp. (MW016057.1)7619786.090.00OR532391Haplosclerida sp. (MW016166.1)7549785.950.00aHaliclona oculata (HQ379251.1)7509786.070.0018S1000aSiphonochalina sp. (DQ927311.1)181410099.40.00OR532382aCladocroce sp. (KC902202.1)175110098.30.00Calyx sp. (DQ927313.1)175110098.30.00SP628S764aDiacarnus sp. (KU060338.1)7375599.750.00OR532392Negombata sp. (OM406250.1)7698589.680.0018S1000aDiacarnus sp. (OP895635.1)18471001000.00OR532383Diacarnus bismarckensis (KC902137.1)183610099.80.00Neopodospongia cf. normani (KC902112.1)177710098.70.00SP728S709aAxinella sp. (AY561925.2)12431001000.00OR532393aAxinella corrugate (AY864741.1)121610099.260.0018S1000aAxinella verrucosa (KX622144.1)181410099.40.00OR532384Axinella corrugate (KC901907.1)181410099.40.00aProsuberites laughlini (EF654529.1)180810099.30.00SP828S750aHaliclona oculata (HQ379251.1)8119787.620.00OR532394Haliclona melissae (OP526584.1)7919587.590.00aCallyspongia sp. (MW016057.1)7609786.150.0018S1050aCladocroce sp. (0CDN9562-C) (KC902202.1)183610098.290.00OR532385Calyx sp. (DQ927313.1)183610098.290.00Cladocroce sp. (KT900335.1)183110098.190.00SP928S700aAxinella sp. (AY561925.2)12341001000.00OR532395aAxinella corrugate (AY864741.1)120610099.250.0018S900aAxinella verrucosa (KX622144.1)164010099.560.00OR532386Axinella corrugate (KC901907.1)164010099.560.00aProsuberites laughlini (EF654529.1)163510099.440.00Please note that before doing BLAST, all the sequences were trimmed from the beginning and the end to remove the bad quality and low signal sequences.aThese sequences are the reference sequences used in constructing the phylogenetic tree.bGenBank accession number of all samples used in this study.
Phylogenetic analysis
3.2
Two maximum likelihood phylogenetic trees were constructed for the collected samples (SP1–SP9) as well as representatives of related genera from GenBank. The phylogenetic trees were constructed based on 18S rRNA (Fig. 4) and 28S rRNA (Fig. 5). The phylogenetic analysis of all isolates from each cluster revealed that there were at least five different genera of marine sponges (Axinella, Negombata, Siphonochalina, Diacarnus, and an unidentified genus within the order Haplosclerida).Fig. 4. The maximum likelihood phylogenetic tree of the 18S rRNA gene. The tree was constructed using Maximum Likelihood method based on the Tamura-Nei model [23]. The boot-strapping value was 10000 and the number next to the branch indicates the percentage of trees in which the associated taxa clustered together. The scale bar is the scale to which the tree was drawn to. Evolutionary analyses were conducted in MEGA X [24]. Black circle, outgroup to which the tree was rooted to; black triangles; representative sequences from the GenBank and their accession numbers; black squares, the collected samples.Fig. 4. Fig. 5The maximum likelihood phylogenetic tree of the 28S rRNA gene. The tree was constructed using Maximum Likelihood method based on the Tamura-Nei model [23]. The boot-strapping value was 10000 and the number next to the branch indicates the percentage of trees in which the associated taxa clustered together. The scale bar is the scale to which the tree was drawn to. Evolutionary analyses were conducted in MEGA X [24]. Black circle, outgroup to which the tree was rooted to; black triangles; representative sequences from the GenBank and their accession numbers; black squares, the collected samples.Fig. 5
Discussion
4
Little information is available about Red Sea sponge [6,7], while no official data were found for the Jordanian Gulf of Aqaba sponges [7]. To our knowledge, no prior study has focused on clarifying sponge with morphological and molecular taxonomy of marine sponges in the Jordanian Gulf of Aqaba. The results obtained, using partial 18S rRNA and the 28S rRNA genes as DNA barcoding markers, revealed the presence of genera like Axinella, Negombata, Siphonochalina, Diacarnus, and a genus from the order Haplosclerida.
Identification of sponge species is difficult due to the scarcity of diagnostic morphological characters [32]. There are many sponges with little or no spicule diversity, and some sponge families showed a high degree of similarity in their spicules [33]. Therefore, this would preclude discrimination of closely related sponge species. In the current study, we conducted both morphological and molecular methods to identify the collected sponge samples. Based only on spicule type, it was not possible to identify SP1, SP2, SP3, SP4, SP6, SP7, SP8, and SP9. The sample SP5 was similar to the description of genus Siphonochalina [29,34] based on external morphology.
DNA barcoding is a powerful tool for sponge identification, although, according to literature studies about Poriferan DNA markers, there is no single ideal marker for all sponge species [[35], [36], [37], [38]]. While morphological identification of sponges is impossible with only loose identification, DNA fragments can be used for targeting DNA barcodes such as 18S rDNA, 28S rDNA, rDNA ITS, and fragment mtDNA cytochrome oxidase subunit 1 (CO1) [32]. The incomplete sequence entries in GenBank are also limiting sponge species identification besides the potential for misidentification [39]. Study of the molecular biodiversity of demosponges of the Red Sea using a partial sequence of the 28S rRNA and mtDNA, the standard animal barcoding fragment revealed that 28S sequences were more variable in Order Haplosclerida and can be useful to discriminate the OTUs in order to detect the genetic variation in specimens with identical genotypes for COI mtDNA [8]. Opposing this data, in the present study, the 18S rRNA gene was shown to be more variable than the 28S rRNA.
Two DNA markers were used in this study to identify the collected sponge samples at the molecular level. The 18S rRNA helps to identify sponge samples to genus level for SP3, SP5, and SP8 samples compared to the 28S rRNA results. This could be due to the sequences submitted in the database for the 28S being limited and not covering all the genera of sponges, while the 18S rRNA may cover those sponge genera.
The genus Axinella was detected in four out of five sampling sites in this study. Genus Axinella (order: Halichondrida; family: Axinellidae) was found and reported as endemic in the Red Sea [8,40]. The sponge Negombata magnifica was detected in single samples collected in the Big Bay site. This toxic finger sponge, N. magnifica, was reported as an endemic species in the Red Sea under the previous name Latrunculia magnifica [40]. N. magnifica was also reported in the Red Sea, Eilat in 2012 by Belinky and his colleagues in 2012 [41]. Moreover, the sponge Siphonochalina sp. was detected in two samples from two different sites (Phosphate Port and Big Bay sites). Genus Siphonochalina was among the subjects of secondary metabolite research in the Red Sea [42]. While Callyspongia (Callyspongia) siphonella, Callyspongia (Callyspongia) tubulosa, Callyspongia (Euplacella) communis, and Callyspongia reticulata were reported from the Red Sea under the following previous names: Siphonochalina siphonella, Siphonochalina tubulosa, Siphonochalina communis, and Siphonochalina reticulata, respectively [40,43,44].
In this study, the Diacarnus genus was detected in single samples in the King Abdullah Reef site. The distribution of the genus Diacarnus (order: Poecilosclerida; family: Podospongiidae) includes the Red Sea [30]. Diacarnus erythraeanus and Diacarnus spinipoculum were also reported and studied for secondary metabolites in the Red Sea [[45], [46], [47], [48], [49]]. While a marine sponge from an unidentified genus within the order Haplosclerida was detected in the SP8 sample from Al-Sodasiat site in this study. Notably, our results showed that the highest identity based on the 28S rRNA sequence was with Haliclona oculata HQ379251.1 and Callyspongia sp. 1 JV-2020 MW016057. Meanwhile, the highest identity based on the 18S RNA (around 98.29 %) was with the Cladocroce sp. 0CDN9562-C and Calyx sp. NIWAKD1132. These results suggest that the genus of this sample could not be determined based on the available data. Therefore, in this study, we classify it as an unidentified genus within the order Haplosclerida. Additionally, the lowest identity values were observed for the sequences of SP3 (86.63 %), SP5 (87.05 %), and SP8 (86.6 %), respectively (Table 2), and this could be due to the lack of information in the GenBank database. The results of phylogenetic analysis based on the 18S and 28S rRNA gene sequences imply recognition of five genera (Axinella, Negombata, Siphonochalina, Diacarnus, and an unidentified genus within the order Haplosclerida). It is important for sponge identification to consider not only spicules and the outer morphology but also to apply histological techniques and molecular analysis with multiple DNA markers. Additionally, analyzing spicule morphology in more detail and expanding molecular barcoding with additional markers could provide a more comprehensive understanding of the order-level richness and diversity of sponges in the Red Sea region.
Conclusions
5
This is the first investigation of morphological and molecular identification of marine sponges in the Jordanian Gulf of Aqaba. Giving consideration to the limitations of each of the molecular and morphological approaches, examination of both of the morphological features and the molecular identification is essential to achieve a more reliable identity [39]. Also, the phylogenetic approach will further enrich our understanding of sponge diversity in the region. However, according to previous studies, knowledge of sponge taxonomy in the Red Sea remains in early stages; hence, more research is needed to fully document the sponge species present in the Red Sea. Upgraded databases are urgently needed as guides for future work.
CRediT authorship contribution statement
Zeinab Arabeyyat: Writing – review & editing, Writing – original draft, Visualization, Validation, Supervision, Software, Resources, Project administration, Methodology, Investigation, Funding acquisition, Formal analysis, Data curation, Conceptualization. Mais Sweiss: Writing – review & editing, Validation, Software, Resources, Methodology, Investigation, Formal analysis, Data curation, Conceptualization. Abdalmajeed Alajlouni: Methodology, Conceptualization. Neda'a Al-Ajlouni: Methodology. Marwan Mahmoud: Methodology. Sura Shartooh: Methodology. Farah Alsoqi: Software, Methodology. Maysoon Kteifan: Software, Methodology.
Ethical statement
No ethical statement was reported.
Data availability statement
Data will be made available on request.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1El-Naggar H.A.Human impacts on coral reef ecosystem Natural Resources Management and Biological Sciences. Intech Open, Febvol. 17202110.5772/intechopen.88841 · doi ↗
- 2Wilkinson S.Brodie J.E.Jone.Frew S.Catchment management and coral reef conservation, a practical guide for coastal resource managers to reduce damage from catchment areas based on best practice case studies 2011
- 3Kochzius M.Community structure of coral reef fishes in El Quadim Bay (El Quseir, Egyptian Red Sea coast)Zool. Middle East 4212007899810.1080/09397140.2007.10638251 · doi ↗
- 4Wulff J.Ecological interactions and the distribution, abundance, and diversity of sponges Adv. Mar. Biol.61201227334410.1016/B 978-0-12-387787-1.00003-922560780 · doi ↗ · pubmed ↗
- 5Wilkinson C.R.Net primary productivity in coral reef sponges Science 2194583198341041210.1126/science.219.4583.41017815320 · doi ↗ · pubmed ↗
- 6Berumen M.L.Hoey A.S.Bass W.H.Bouwmeester J.Catania D.Cochran J.E.Khalil M.T.Miyake S.Mughal M.R.Spät J.L.Saenz-Agudelo P.The status of coral reef ecology research in the Red Sea Coral Reefs 32201373774810.1007/s 00338-013-1055-8 · doi ↗
- 7Wooster M.K.Voigt O.Erpenbeck D.Wörheide G.Berumen M.L.Sponges of the Red Sea Voolstra C.Berumen M.Coral Reefs of the Red Sea'. Coral Reefs of the Worldvol. 112019 Springer Cham 9112210.1007/978-3-030-05802-9_6 · doi ↗
- 8Erpenbeck D.Voigt O.Al-Aidaroos A.M.Berumen M.L.Büttner G.Catania D.Guirguis A.N.Paulay G.Schätzle S.Wörheide G.Molecular biodiversity of Red Sea demosponges Mar. Pollut. Bull.105201650751410.1016/j.marpolbul.2015.12.00426776057 · doi ↗ · pubmed ↗