Morphological characteristics and phylogenetic analyses revealed five new species (Basidiomycota) from Southwestern China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
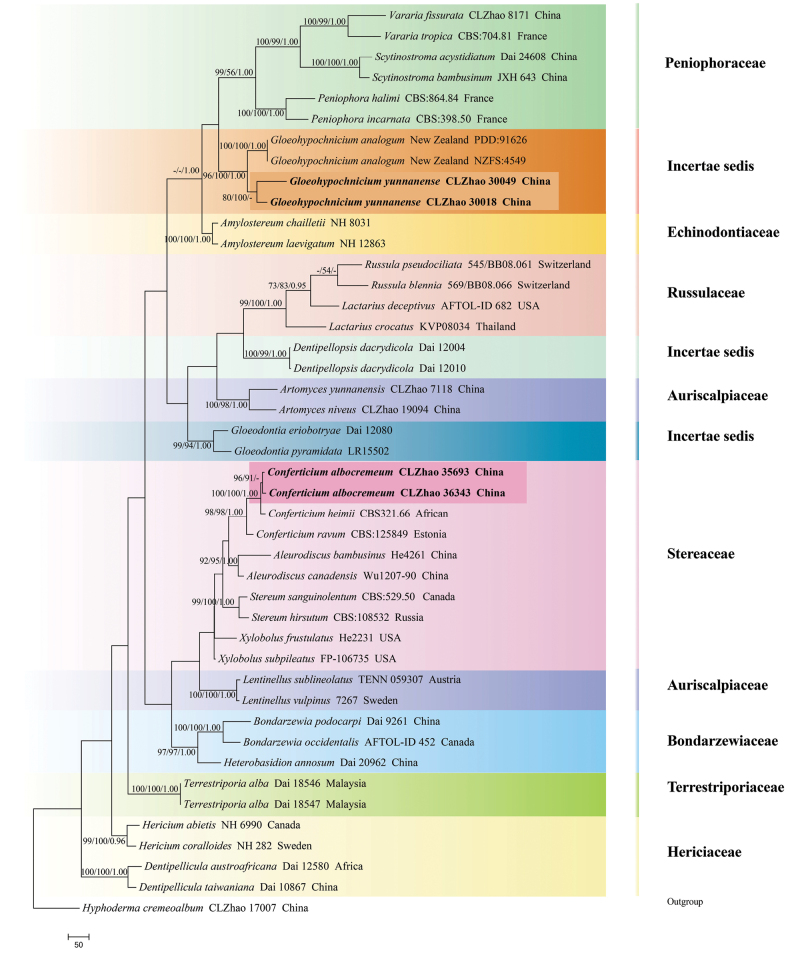 Figure 1
Figure 1 Figure 2
Figure 2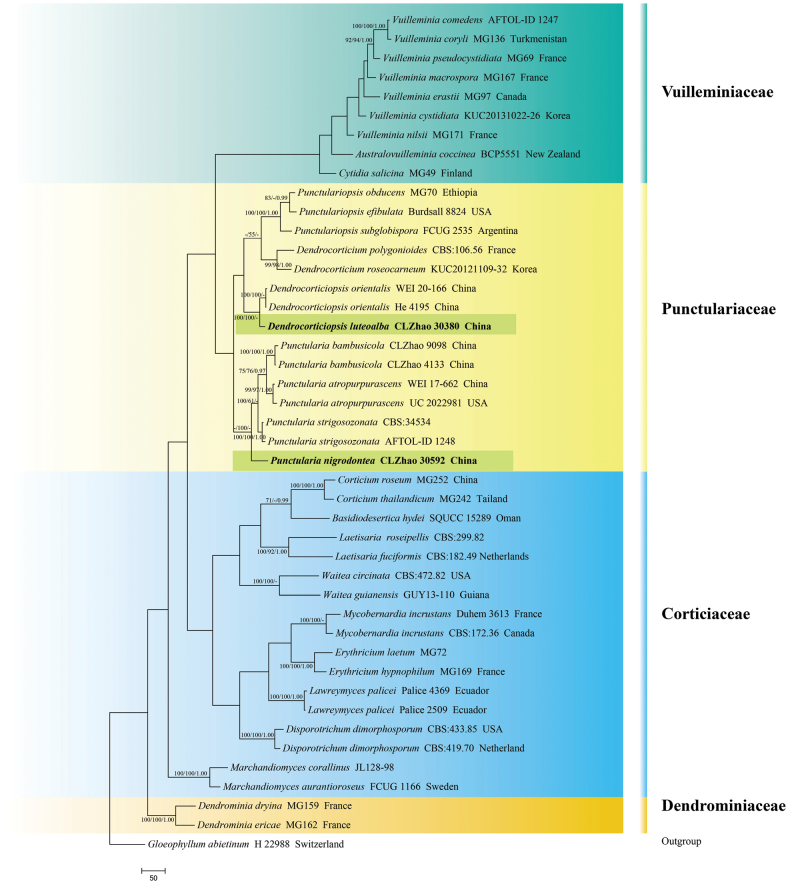 Figure 3
Figure 3 Figure 4
Figure 4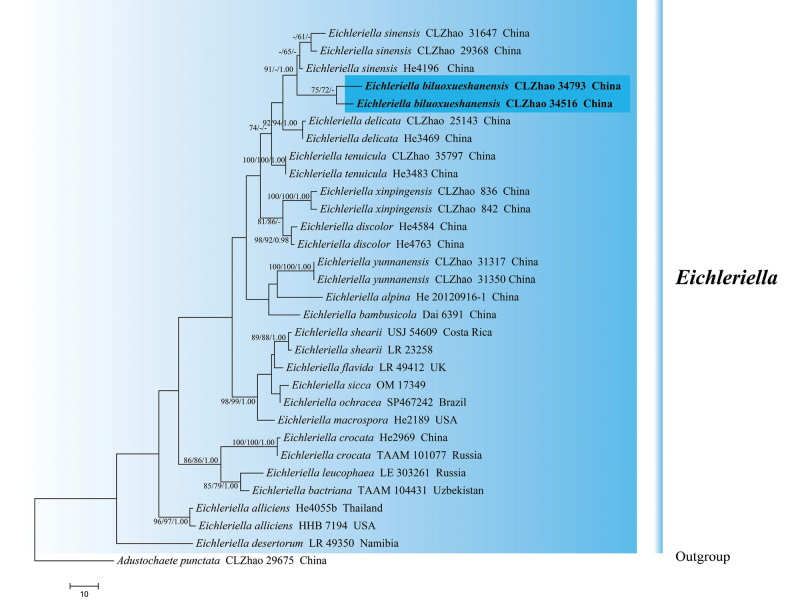 Figure 5
Figure 5 Figure 6
Figure 6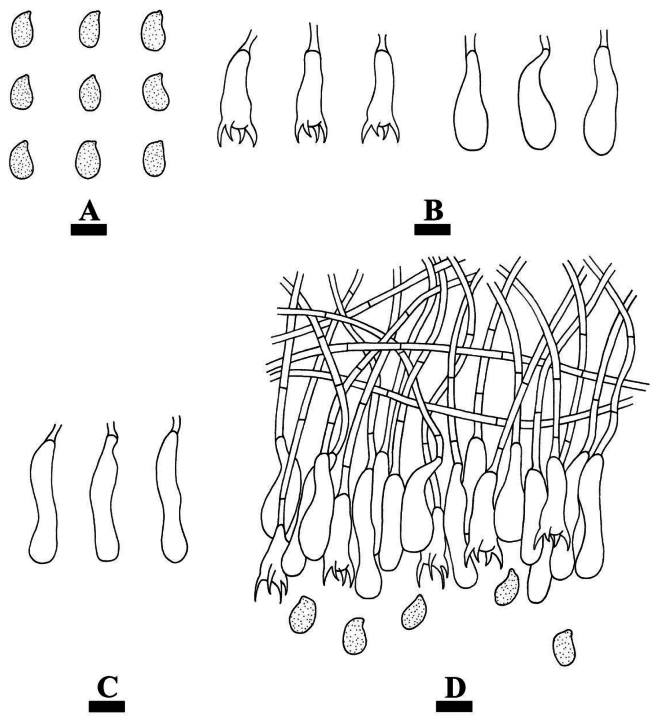 Figure 7
Figure 7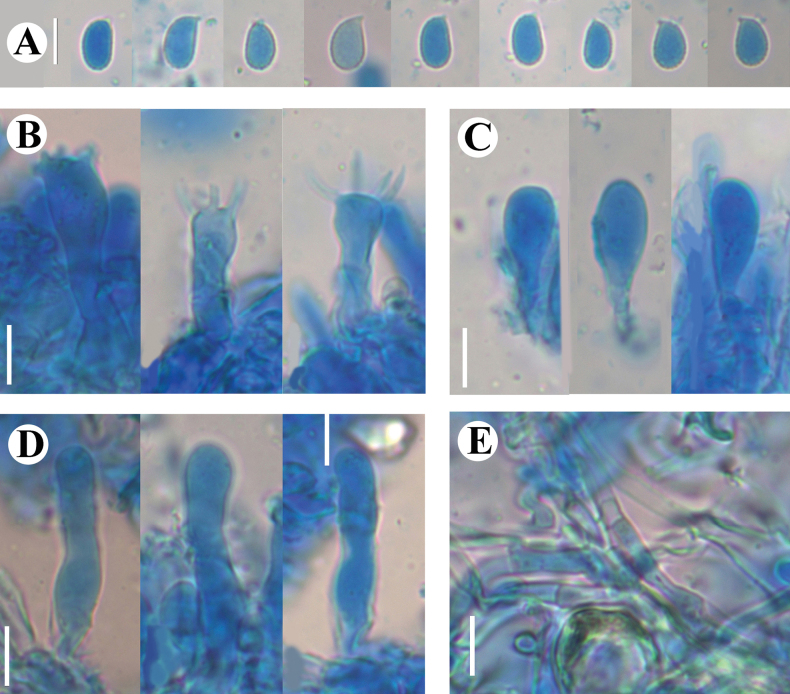 Figure 8
Figure 8 Figure 9
Figure 9 Figure 10
Figure 10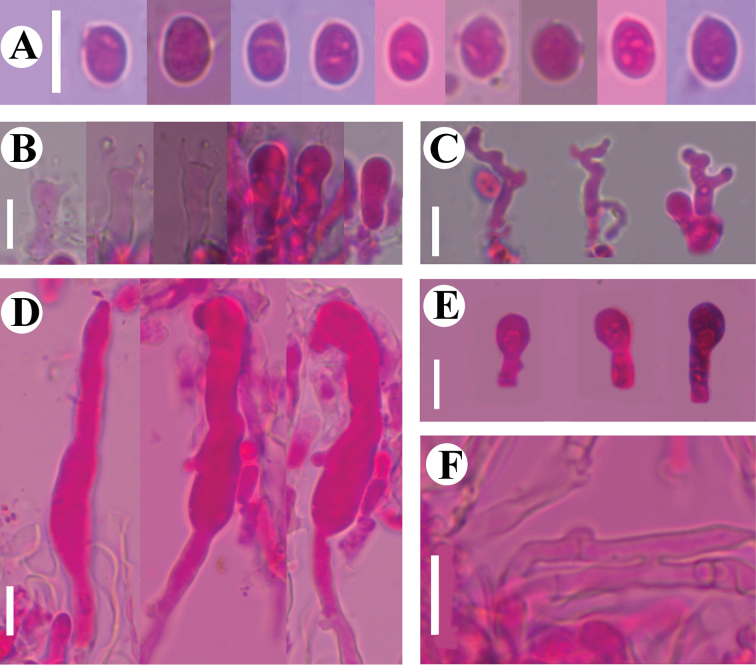 Figure 11
Figure 11 Figure 12
Figure 12 Figure 13
Figure 13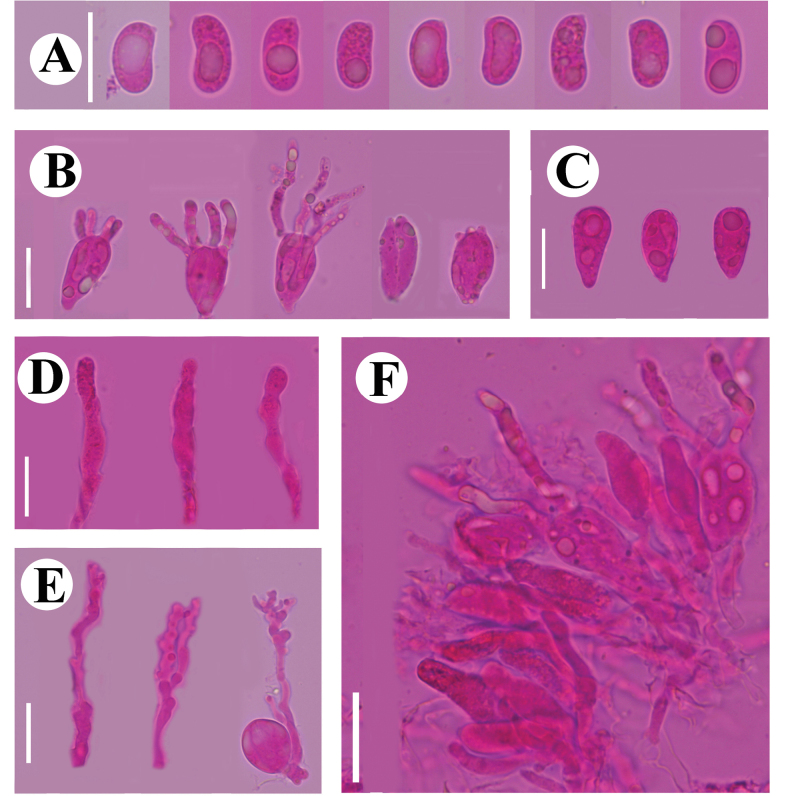 Figure 14
Figure 14 Figure 15
Figure 15 Figure 16
Figure 16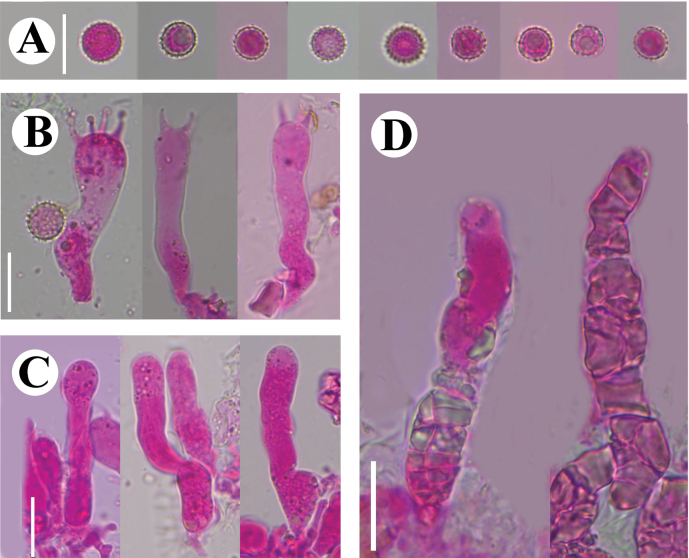 Figure 17
Figure 17 Figure 18
Figure 18 Figure 19
Figure 19 Figure 20
Figure 20| Taxa | Voucher no. | Locality | GenBank accession no. | References | |
|---|---|---|---|---|---|
|
|
| ||||
|
| He2357 | China |
|
|
|
|
| CBS 233.86 | France | – |
|
|
|
| T614 | USA | – |
|
|
|
| FP-100292 | USA | – |
|
|
|
| CLZhao 29675 | China |
|
|
|
|
| He2712 | China |
|
|
|
|
| He4261 | China |
|
|
|
|
| LWZ 20180514-18 | China |
|
|
|
|
| LWZ 20180904-14 | China |
|
|
|
|
| CLZhao 33798 | China |
| – |
|
|
| CLZhao 25093 | China |
|
|
|
|
| CLZhao 8106 | China |
|
|
|
|
| FP-106715 | USA |
|
|
|
|
| HHB 8476 | USA |
|
|
|
|
| NH 8031 | – |
|
|
|
|
| NH 12863 | – |
|
|
|
|
| Miettinen 14774 | Finland |
| – |
|
|
| WD 2207 | Japan |
|
|
|
|
| CLZhao 19094 | China |
|
|
|
|
| CLZhao 7118 | China |
|
|
|
|
| JT 04 | UK |
|
|
|
|
| Dai 13621 | China |
|
|
|
|
| FO 25132 | Germany |
|
|
|
|
| TUFC 12920 | Japan |
|
|
|
|
| Dai 13336 | China |
|
|
|
|
| BCP5551 | New Zealand |
|
|
|
|
| SQUCC 15289 | Oman |
|
|
|
|
| AFTOL-ID 452 | Canada |
|
|
|
|
| Dai 9261 | China |
|
|
|
|
| CLZhao 35693* | China |
|
| Present study |
|
| CLZhao 36343 | China |
|
| Present study |
|
| CLZhao 37176 | China |
|
| Present study |
|
| CLZhao 39238 | China |
| – | Present study |
|
| CLZhao 34654 | China |
| – |
|
|
| CLZhao 34662 | China |
| – |
|
|
| CBS321.66 | African |
|
|
|
|
| CLZhao 21515 | China |
| – | Present study |
|
| G07_P24A | Switzerland |
| – |
|
|
| CBS:125849 | Estonia |
|
|
|
|
| NH13291 | USA |
|
|
|
|
| MG252 | China |
|
|
|
|
| MG242 | Tailand |
|
|
|
|
| MG49 | Finland |
|
|
|
|
| CLZhao 30380* | China |
|
| Present study |
|
| He 4195 | China |
|
|
|
|
| WEI 20-166 | China |
|
|
|
|
| CBS:106.56 | France |
|
|
|
|
| KUC20121109-32 | Korea |
|
|
|
|
| MG159 | France |
|
|
|
|
| MG162 | France |
|
|
|
|
| Dai 12580 | Africa |
|
|
|
|
| Dai 10867 | China |
|
|
|
|
| Dai 12004 | – |
|
|
|
|
| Dai 12010 | – | – |
|
|
|
| CBS:419.70 | Netherland |
|
|
|
|
| CBS:433.85 | USA |
|
|
|
|
| He4055b | Thailand |
| – |
|
|
| HHB 7194 | USA |
|
|
|
|
| He 20120916-1 | China |
|
|
|
|
| TAAM 104431 | Uzbekistan |
|
|
|
|
| Dai 6391 | China |
| – |
|
|
| CLZhao 34516* | China |
|
| Present study |
|
| CLZhao 34793 | China |
| – | Present study |
|
| He2969 | China |
|
|
|
|
| TAAM 101077 | Russia |
|
|
|
|
| CLZhao 25143 | China |
| – | Present study |
|
| He3469 | China |
|
|
|
|
| LR 49350 | Namibia |
|
|
|
|
| He4584 | China |
|
|
|
|
| He4763 | China |
|
|
|
|
| LR 49412 | UK |
|
|
|
|
| LE 303261 | Russia |
|
|
|
|
| He2189 | USA |
|
|
|
|
| SP467242 | Brazil |
| – |
|
|
| LR 23258 | – |
| – |
|
|
| USJ 54609 | Costa Rica |
|
| Weiß et al. 2001 |
|
| OM 17349 | USA |
|
|
|
|
| CLZhao 29368 | China |
| – | Present study |
|
| CLZhao 31647 | China |
|
| Present study |
|
| He4196 | China |
|
|
|
|
| CLZhao 35797 | China |
| – | Present study |
|
| He3483 | China |
|
|
|
|
| CLZhao 836 | China |
|
|
|
|
| CLZhao 842 | China |
|
|
|
|
| CLZhao 31317 | China |
|
|
|
|
| CLZhao 31350 | China |
|
|
|
|
| Miettinen 14314 | Indonesia |
|
|
|
|
| Miettinen 16431 | Malaysia |
|
|
|
|
| MG169 | France |
|
|
|
|
| MG72 | – |
|
|
|
|
| VS 3921 | Russia |
|
|
|
|
| MW 355 | Germany |
|
| Weiß et al. 2001 |
|
| MW 313 | Germany |
|
| Weiß et al. 2001 |
|
| Dai 20833 | China |
|
|
|
|
| Wu 270 | China |
|
|
|
|
| OM 19136 | Finland |
|
|
|
|
| Norway |
|
|
| |
|
| FCUG2731 | Russia |
|
|
|
|
| Dai 12080 | – |
|
|
|
|
| LR15502 | – |
|
|
|
|
| NZFS:4549 | New Zealand |
| – |
|
|
| PDD:91626 | New Zealand |
| – |
|
|
| CLZhao 30018 | China |
|
| Present study |
|
| CLZhao 30049* | China |
|
| Present study |
|
| H 22988 | Switzerland |
|
|
|
|
| NH 6990 | Canada |
|
|
|
|
| NH 282 | Sweden |
|
|
|
|
| Dai 20962 | China |
|
|
|
|
| LWZ 20180512-25 | Australia |
|
|
|
|
| LE 38182 | Sweden |
|
|
|
|
| He 4915 | China |
|
|
|
|
| Yuan 1600 | China |
|
|
|
|
| Dai 17193 | China |
|
|
|
|
| CLZhao 17007 | China |
|
|
|
|
| Thailand |
|
|
| |
|
| AFTOL-ID 682 | USA |
|
|
|
|
| CBS:182.49 | Netherlands |
|
|
|
|
| CBS:299.82 | – |
|
|
|
|
| Palice 2509 | Ecuador |
|
|
|
|
| Palice 4369 | Ecuador |
|
|
|
|
| TENN 059307 | Austria |
| – |
|
|
| 7267 | Sweden |
| – |
|
|
| FCUG 1166 | Sweden |
|
|
|
|
| JL128-98 | – |
|
|
|
|
| V.Spirin4244 | Sweden |
|
|
|
|
| HK9808 | Sweden |
|
|
|
|
| CBS:172.36 | Canada |
|
|
|
|
| Duhem 3613 | France |
|
|
|
|
| He2921 | China |
|
|
|
|
| Wu0807-41 | Japan | – |
|
|
|
| CBS:864.84 | France |
|
|
|
|
| CBS:398.50 | France |
|
|
|
|
| Yuan 5691 | China |
|
|
|
|
| WD 548 | Japan |
|
|
|
|
| UC 2022981 | USA |
| – | Knijn & Ferretti 2018 |
|
| WEI 17-662 | China |
|
|
|
|
| CLZhao 4133 | China |
|
|
|
|
| CLZhao 9098 | China |
|
|
|
|
| CLZhao 30592* | China |
|
| Present study |
|
| AFTOL-ID 1248 | – |
|
|
|
|
| CBS:34534 | – |
|
|
|
|
| Burdsall 8824 | USA |
|
|
|
|
| MG70 | Ethiopia |
|
|
|
|
| FCUG 2535 | Argentina |
|
|
|
|
| 569/BB08.066 | Switzerland |
|
|
|
|
| 545/BB08.061 | Switzerland |
|
|
|
|
| TN 2722 | Canada |
|
|
|
|
| VS 7674 | Russia |
|
|
|
|
| Dai 24608 | China |
|
|
|
|
| JXH 643 | China |
|
|
|
|
| 236 | Netherlands |
|
|
|
|
| SPG6799 | Argentina |
|
|
|
|
| SPG6874 | Argentina |
|
|
|
|
| CBS:108532 | Russia |
|
|
|
|
| CBS:529.50 | Canada |
|
|
|
|
| Dai 18546 | Malaysia |
|
|
|
|
| Dai 18547 | Malaysia |
|
|
|
|
| Brazil |
|
|
| |
|
| SP467241 | Brazil |
|
|
|
|
| LE 303446 | Russia |
|
|
|
|
| CLZhao 8171 | China |
|
|
|
|
| CBS:704.81 | France |
|
|
|
|
| AFTOL-ID 1247 | – |
|
|
|
|
| MG136 | Turkmenistan |
|
|
|
|
| KUC20131022-26 | Korea |
|
|
|
|
| MG97 | Canada |
|
|
|
|
| MG167 | France |
|
|
|
|
| MG171 | France |
|
|
|
|
| MG69 | France |
|
|
|
|
| CBS:472.82 | USA |
|
|
|
|
| GUY13-110 | Guiana |
|
|
|
|
| He2231 | USA |
|
|
|
|
| FP-106735 | USA | – |
|
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Lichen and fungal ecology · Plant Pathogens and Fungal Diseases
Introduction
The kingdom of fungi is one of the most diverse groups of living organisms on earth; its members occur across a broad range of ecosystems, including extreme environments, with an estimated number of species in the range of 2–3 million (Bhunjun et al. 2022, 2024; Wijayawardene et al. 2022, 2024; Dong et al. 2024a; Hyde et al. 2024a, b). Based on molecular phylogenetic evidence, numerous new taxa have been discovered in the past ten years (Wang and Cai 2023; Yuan et al. 2023; Zhao et al. 2023; Deng et al. 2024b; Dong et al. 2024b; He et al. 2024; Qin et al. 2024).
The genus Conferticium Hallenb. (Stereaceae, Russulales) was erected in 1980 and typified by C.insidiosum (Bourdot & Galzin) Hallenb., which is characterized by the resupinate basidiomes with membranaceous to ceraceous, smooth to tuberculate hymenophore, a monomitic simple-septate hyphae, and the numerous cylindrical, sinuous gloeocystidia (Bernicchia and Gorjón 2010). Based on the MycoBank database (http://www.mycobank.org, accessed on 27 January 2025) and the Index Fungorum (http://www.indexfungorum.org, accessed on 27 January 2025), six specific and infraspecific names were registered in Conferticium, and it is a small genus only including five widely recognized species up to now.
The genus Dendrocorticiopsis Sheng H. Wu et al. (Punctulariaceae, Corticiales) was introduced by Sheng H. Wu, with the description of one species, D.orientalis Sheng H. Wu et al. (Wei et al. 2022). It is conventionally defined by having strictly resupinate basidiomata, an ivory hymenophore, a monomitic hyphal system with clamped hyphae, encrusted cystidia, dendrohyphidia, and ellipsoid to ovoid basidiospores (Wei et al. 2022). Based on the MycoBank database (http://www.mycobank.org, accessed on 27 January 2025) and the Index Fungorum (http://www.indexfungorum.org, accessed on 27 January 2025), Dendrocorticiopsis is a monotypic genus.
The genus Eichleriella Bres. was introduced in 1903, typified by E.incarnata Bres., and it is a species-rich genus that belongs to Auriculariaceae (Auriculariales). The genus is characterized by annual or short-living perennial, leathery to ceraceous basidiomata with smooth, pale-colored hymenophore (in some species covered by spines), a monomitic to dimitic hyphal system with clamped genitive hyphae, cystidia often present, longitudinally septate basidia with 2- or 4-celled, and colorless, cylindrical to narrowly cylindrical basidiospores (Malysheva and Spirin 2017; Li et al. 2023; Deng et al. 2024a). Based on the MycoBank database (http://www.mycobank.org, accessed on 27 January 2025) and the Index Fungorum (http://www.indexfungorum.org, accessed on 27 January 2025), the genus Eichleriella has 32 specific and registered names, with 22 species accepted worldwide (Malysheva and Spirin 2017, Liu et al. 2019, Li et al. 2023; Deng et al. 2024a).
The genus Gloeohypochnicium (Parmasto) Hjortstam (Russulales), typified by G.analogum (Bourdot & Galzin) Hjortstam (Bernicchia and Gorjón 2010), is characterized by the resupinate basidiomes with smooth to tuberculate hymenophore, a monomitic hyphal system with clamps on generative hyphae, the numerous cylindrical, sinuous gloeocystidia, and globose to ellipsoid, thick-walled, warted basidiospores (Bernicchia and Gorjón 2010). Based on the MycoBank database (http://www.mycobank.org, accessed on 27 January 2025) and the Index Fungorum (http://www.indexfungorum.org, accessed on 27 January 2025), Gloeohypochnicium has registered two specific and infraspecific names, and it is a small genus only including two widely recognized species so far (Bernicchia and Gorjón 2010; He et al. 2024).
Punctularia Patouillard (Punctulariaceae, Corticiales) was typified with P.tuberculosa (Pat.) Pat. & Lagerh. (current name P.atropurpurascens (Berk. & Broome) Petch, which is characterized by resupinate to effused-reflexed basidiomata, gelatinous when fresh, rigid upon drying, tuberculate or radial ridges hymenophore, a monomitic hyphal system with clamped generative hyphae, yellowish to brown dendrohyphidia, and thin-walled, smooth, ellipsoid, acyanophilous basidiospores (Bernicchia and Gorjón 2010). Based on the MycoBank database (http://www.mycobank.org, accessed on 27 January 2025) and the Index Fungorum (http://www.indexfungorum.org, accessed on 27 January 2025), Punctularia has registered six specific and infraspecific names, and three species have been recognized worldwide up to now (Bernicchia and Gorjón 2010; Guan et al. 2021; He et al. 2024).
Molecular phylogenetic approaches have revolutionized the fungal taxonomy of Basidiomycota in the last decades, and these advances have greatly enhanced our knowledge of species diversity in Basidiomycota (Lücking et al. 2021; He et al. 2022; Wang et al. 2023; Dong et al. 2024a). The family Auriculariaceae is the largest and best-supported clade in the order Auriculariales and consists of a large group of wood-decaying fungi with varied basidiomes (Dong et al. 2024b; He et al. 2024). Corticioid and stereoid taxa are numerous in Auriculariaceae and are typically classified into three main genera: Eichleriella Bres., Exidiopsis (Bref.) A. Møller, and Heterochaete Pat. (Malysheva and Spirin 2017; Li et al. 2023; Deng et al. 2024a). DNA sequence-based classification and identification of the genus Eichleriella (Auriculariaceae) have reported that six species have been described from China recently (Li et al. 2023; Deng et al. 2024a). Corticiales K.H. Larss. is a small order of corticioid fungi with four families *viz.*Corticiaceae Herter, Dendrominiaceae Ghobad-Nejhad, Punctulariaceae Donk, and Vuilleminiaceae Maire ex Lotsy (Wei et al. 2022; He et al. 2024). Most of the previous studies of Punctulariaceae focused on European species (Bernicchia and Gorjón 2010; Gorjón and Bernicchia 2017). In China, the research on this family mainly focuses on the genera Dendrocorticiopsis and Punctularia; some new taxa have been proposed based on a combination of morphological features and molecular data (Guan et al. 2021; Wei et al. 2022; He et al. 2024). Members of the Russulales Kreisel ex P.M. Kirk, P.F. Cannon & J.C. David exhibit diverse basidiome structures, which can range from agaricoid to discoid, clavarioid, polyporoid, corticoid, and even gasteroid, and the multigene phylogenetic analyses clarify the evolutionary relationships of some small genera in the latest study (Yuan et al. 2021; He et al. 2024). According to recent research in molecular systematics, the genus Conferticium (Stereaceae, Russulales) has reported one new species, C.fissuratum Xin Yang & C.L. Zhao from Yunnan Province (Bernicchia and Gorjón 2010; Shen et al. 2024). The taxonomic status of the genus Gloeohypochnicium is unclear, and it was only placed in the order Russulales, and it is a mystery genus; no new taxa in the genus have been described for nearly a decade (He et al. 2022, 2024). However, the phylogeny of the genus Gloeohypochnicium is ambiguous due to a lack of molecular evidence and morphological data.
In this paper, we presented the morphological characteristics and multigene molecular analyses with ITS and nLSU DNA markers to support the taxonomy and phylogenetic position of five new species.
Materials and methods
Sample collection and herbarium specimen preparation
Fresh basidiomata growing on angiosperm branches were collected from Dehong, Diqing, Tengchong, and Xishuangbanna of Yunnan Province, P.R. China. The samples were photographed in situ, and fresh macroscopic details were recorded. Photographs were recorded using a Jianeng 80D camera (Tokyo, Japan). Specimens were dried in an electric food dehydrator at 45 °C (Hu et al. 2022), then the specimens were sealed in an envelope and zip-lock plastic bags and labeled (Zhao et al. 2023). The dried specimens were deposited in the herbarium of the Southwest Forestry University (SWFC), Kunming, Yunnan Province, China.
Morphology
The macromorphological descriptions were based on field notes and photos captured in the field and lab. The color terminology follows Petersen (1996). The micromorphological data were obtained from the dried specimens observed under a Nikon Eclipse E100 light microscope following Zhao and Wu (2017). The following abbreviations are used: KOH = 5% potassium hydroxide water solution, CB– = acyanophilous, IKI+ = amyloid, IKI– = both inamyloid and indextrinoid, L = mean spore length (arithmetic average for all spores), W = mean spore width (arithmetic average for all spores), Q = variation in the L/W ratios between the specimens studied, and n = a/b (number of spores (a) measured from given number (b) of specimens).
DNA extraction, PCR, and sequencing
The CTAB rapid plant genome extraction kit-DN14 (Aidlab Biotechnologies Co., Ltd., Beijing, China) was used to obtain genomic DNA from the dried specimens according to the manufacturer’s instructions. The nuclear ribosomal internal transcribed spacer (ITS) region was amplified with the primer pair ITS5/ITS4 (White et al. 1990). The nuclear large subunit (nLSU) region with the primer pair LR0R/LR7 (Vilgalys and Hester 1990). The PCR procedure for ITS was as follows: initial denaturation at 95 °C for 3 min, followed by 35 cycles at 94 °C for 40 s, 58 °C for 45 s, and 72 °C for 1 min, and a final extension of 72 °C for 10 min. The PCR procedure for nLSU was as follows: initial denaturation at 94 °C for 1 min, followed by 35 cycles at 94 °C for 30 s, 48 °C for 1 min, and 72 °C for 1.5 min, and a final extension of 72 °C for 10 min. The PCR products were purified and sequenced at Kunming Tsingke Biological Technology Limited Company (Yunnan Province, P.R. China). All newly generated sequences were deposited in GenBank (Table 1).
Molecular phylogeny
The sequences were aligned in MAFFT version 7 using the G-INS-i strategy (Katoh et al. 2019). The alignment was adjusted manually using AliView version 1.27 (Larsson 2014). (1) Hyphodermacremeoalbum (Höhn. & Litsch.) Jülich was assigned as an outgroup to root trees in the ITS+nLSU analysis (Fig. 1) (Dong et al. 2024a); (2) Varariafissurata Y.L. Deng & C.L. Zhao were assigned as an outgroup to root trees following the ITS+nLSU analysis (Fig. 2) (Deng et al. 2024b); (3) Gloeophyllumabietinum (Bull.) P. Karst. was assigned as an outgroup to root trees following the ITS+nLSU analysis (Fig. 3) (He et al. 2014); (4) Sistotremabrinkmannii (Bres.) J. Erikss. was assigned as an outgroup to root trees following the ITS+nLSU analysis (Fig. 4) (Alvarenga and Gibertoni 2021); (5) Adustochaetepunctata J.H. Dong & C.L. Zhao were assigned as an outgroup to root trees following the ITS+nLSU analysis (Fig. 5) (Dong et al. 2024b).
Maximum parsimony strict consensus tree illustrating the phylogeny of Conferticium and Gloeohypochnicium and related genera in the order Russulales, based on ITS+nLSU sequences; branches are labeled with maximum likelihood bootstrap value ≥ 70%, parsimony bootstrap value ≥ 50%, and Bayesian posterior probabilities ≥ 0.95.
Maximum parsimony strict consensus tree illustrating the phylogeny of Conferticium and related genus in the family Stereaceae, based on ITS+nLSU sequences; branches are labeled with maximum likelihood bootstrap value ≥ 70%, parsimony bootstrap value ≥ 50%, and Bayesian posterior probabilities ≥ 0.95.
Maximum parsimony strict consensus tree illustrating the phylogeny of Dendrocorticiopsis and Punctularia and related genera in the order Corticiales, based on ITS+nLSU sequences; branches are labeled with maximum likelihood bootstrap value ≥ 70%, parsimony bootstrap value ≥ 50%, and Bayesian posterior probabilities ≥ 0.95.
Maximum parsimony strict consensus tree illustrating the phylogeny of Eichleriella and related genera in the family Auriculariaceae, based on ITS+nLSU sequences; branches are labeled with maximum likelihood bootstrap value ≥ 70%, parsimony bootstrap value ≥ 50%, and Bayesian posterior probabilities ≥ 0.95.
Maximum parsimony strict consensus tree illustrating the phylogeny of Eichleriella and related species in the genus Eichleriella, based on ITS+nLSU sequences; branches are labeled with maximum likelihood bootstrap value ≥ 70%, parsimony bootstrap value ≥ 50%, and Bayesian posterior probabilities ≥ 0.95.
Maximum parsimony (MP), maximum likelihood (ML), and Bayesian inference (BI) analyses were applied to the combined datasets following a previous study (Wu et al. 2022; Dong et al. 2024a), and the tree construction procedure was performed in PAUP* version 4.0b10 (Swofford 2002). All characters were equally weighted, and gaps were treated as missing data. Trees were inferred using the heuristic search option with TBR branch swapping and 1,000 random sequence additions. Max trees were set to 5000, branches of zero length were collapsed, and all parsimonious trees were saved. Clade robustness was assessed using bootstrap (BT) analysis with 1000 replicates (Felsenstein 1985). Descriptive tree statistics, tree length (TL), consistency index (CI), retention index (RI), rescaled consistency index (RC), and homoplasy index (HI) were calculated for each maximum parsimonious tree generated. The multiple sequence alignment was also analyzed using maximum likelihood (ML) in RAxML-HPC2 (Miller et al. 2012). Branch support (BS) for ML analysis was determined by 1000 bootstrap replicates.
MrModeltest 2.3 (Nylander 2004) was used to determine the best-fit evolution model for each dataset for Bayesian inference (BI), which was performed using MrBayes 3.2.7a with a general time reversible model of DNA substitution and a gamma distribution rate variation across sites (Ronquist et al. 2012). Four Markov chains were run twice from a random starting tree for 0.4 million generations of the datasets (Fig. 1), 0.4 million generations of the datasets (Fig. 2), 1.6 million generations of the datasets (Fig. 3), 0.8 million generations of the datasets (Fig. 4), and for 0.6 million generations of the datasets (Fig. 5), and the tree was sampled every 1000 generations. The first one-fourth of all generations were discarded as burn-in. The majority rule consensus tree of all remaining trees was calculated. Branches were considered as significantly supported if they received a maximum likelihood bootstrap value (BS) ≥70%, maximum parsimony bootstrap value (BT) ≥50%, or Bayesian posterior probabilities (BPP) ≥0.95.
Results
The phylogeny of Conferticium
The datasets based on ITS+nLSU (Fig. 1) comprise sequences from 44 fungal specimens representing 39 species. The datasets had an aligned length of 2,270 characters, of which 1,183 characters are constant, 293 are variable and parsimony-uninformative, and 794 are parsimony-informative. Maximum parsimony analysis yielded 1 equally parsimonious tree (TL = 4447, CI = 0.4160, H = 0.5840, RI = 0.5275, RC = 0.2194). Bayesian analysis and ML analysis resulted in a similar topology as MP analysis with an average standard deviation of split frequencies of 0.008678 (BI), and the effective sample size (ESS) across the two runs is double the average ESS (avg ESS) = 144.5. The phylogenetic tree (Fig. 1) inferred from ITS+nLSU sequences revealed that the new species Conferticiumalbocremeum grouped into the genus Conferticium and clustered into the family Stereaceae (Russulales), in which it was retrieved as a sister to C.heimii (Boidin) Sheng H. Wu.
The datasets based on ITS+nLSU (Fig. 2) comprise sequences from 31 fungal specimens representing 23 species. The datasets had an aligned length of 2,136 characters, of which 1,456 characters are constant, 298 are variable and parsimony-uninformative, and 382 are parsimony-informative. Maximum parsimony analysis yielded 1 equally parsimonious tree (TL = 1491, CI = 0.6237, HI = 0.3763, RI = 0.5681, RC = 0.3544). Bayesian analysis and ML analysis resulted in a similar topology as MP analysis with an average standard deviation of split frequencies of 0.013157 (BI), and the effective sample size (ESS) across the two runs is double the average ESS (avg ESS) = 265.5. The phylogenetic tree (Fig. 2) inferred from ITS+nLSU sequences revealed that Conferticiumalbocremeum grouped into the genus Conferticium and clustered into the family Stereaceae, in which it was closely related to C.heimii.
The phylogeny of Dendrocorticiopsis
The datasets based on ITS+nLSU (Fig. 3) comprise sequences from 44 fungal specimens representing 37 species. The datasets had an aligned length of 2,181 characters, of which 1,313 characters are constant, 232 are variable and parsimony-uninformative, and 736 are parsimony-informative. Maximum parsimony analysis yielded 1 equally parsimonious tree (TL = 3423, CI = 0.4368, HI = 0.5632, RI = 0.5695, RC = 0.2488). Bayesian analysis and ML analysis resulted in a similar topology as MP analysis with an average standard deviation of split frequencies of 0.013154 (BI), and the effective sample size (ESS) across the two runs is double the average ESS (avg ESS) = 658. The phylogenetic tree (Fig. 1) inferred from ITS+nLSU sequences revealed that the new species Dendrocorticiopsisluteoalba grouped into the genus Dendrocorticiopsis and clustered into the family Punctulariaceae (Corticiales), in which it was retrieved as a sister to D.orientalis Sheng H. Wu et al.
The phylogeny of Eichleriella
The datasets based on ITS+nLSU (Fig. 4) comprise sequences from 45 fungal specimens representing 43 species. The datasets had an aligned length of 2,104 characters, of which 1,516 characters are constant, 254 are variable and parsimony-uninformative, and 334 are parsimony-informative. Maximum parsimony analysis yielded 1 equally parsimonious tree (TL = 1783 CI = 0.4773, HI = 0.5227, RI = 0.5633, RC = 0.2688). Bayesian analysis and ML analysis resulted in a similar topology as MP analysis with an average standard deviation of split frequencies of 0.0024569 (BI), and the effective sample size (ESS) across the two runs is double the average ESS (avg ESS) = 361.5. The phylogenetic tree (Fig. 4) inferred from ITS+nLSU sequences revealed that Eichleriellabiluoxueshanensis grouped into the genus Eichleriella and clustered into the family Auriculariaceae, in which it was retrieved as a sister to E.sinensis (Teng) S.H. He & Nakasone.
The datasets based on ITS+nLSU (Fig. 5) comprise sequences from 31 fungal specimens representing 24 species. The datasets had an aligned length of 1,905 characters, of which 1,687 characters are constant, 101 are variable and parsimony-uninformative, and 117 are parsimony-informative. Maximum parsimony analysis yielded 1 equally parsimonious tree (TL = 402, CI = 0.6542, HI = 0.3458, RI = 0.7495, RC = 0.4094). Bayesian analysis and ML analysis resulted in a similar topology as MP analysis with an average standard deviation of split frequencies of 0.009368 (BI), and the effective sample size (ESS) across the two runs is double the average ESS (avg ESS) = 264.5. The phylogenetic tree (Fig. 5) inferred from ITS+nLSU sequences revealed that Eichleriellabiluoxueshanensis grouped into the genus Eichleriella, in which it was grouped with the clade comprising E.sinensis (Teng) S.H. He & Nakasone.
The phylogeny of Gloeohypochnicium
The datasets based on ITS+nLSU (Fig. 1) comprise sequences from 44 fungal specimens representing 39 species. The datasets had an aligned length of 2,270 characters, of which 1,183 characters are constant, 293 are variable and parsimony-uninformative, and 794 are parsimony-informative. Maximum parsimony analysis yielded 1 equally parsimonious tree (TL = 4447, CI = 0.4160, HI = 0.5840, RI = 0.5275, RC = 0.2194). Bayesian analysis and ML analysis resulted in a similar topology as MP analysis with an average standard deviation of split frequencies of 0.008678 (BI), and the effective sample size (ESS) across the two runs is double the average ESS (avg ESS) = 144.5. The phylogenetic tree (Fig. 1) inferred from ITS+nLSU sequences revealed that Gloeohypochniciumyunnanense grouped into the genus Gloeohypochnicium and clustered into the order Russulales, in which it was closely related to G.analogum (Bourdot & Galzin) Hjortstam.
The phylogeny of Punctularia
The datasets based on ITS+nLSU (Fig. 3) comprise sequences from 44 fungal specimens representing 37 species. The datasets had an aligned length of 2,181 characters, of which 1,313 characters are constant, 232 are variable and parsimony-uninformative, and 736 are parsimony-informative. Maximum parsimony analysis yielded 1 equally parsimonious tree (TL = 3423, CI = 0.4368, HI = 0.5632, RI = 0.5695, RC = 0.2488). Bayesian analysis and ML analysis resulted in a similar topology as MP analysis with an average standard deviation of split frequencies of 0.013154 (BI), and the effective sample size (ESS) across the two runs is double the average ESS (avg ESS) = 658. The phylogenetic tree (Fig. 1) inferred from ITS+nLSU sequences revealed that Punctularianigrodontea grouped into the genus Punctularia and clustered into the family Punctulariaceae (Corticiales), in which it was grouped with the clade comprising P.atropurpurascens (Berk. & Broome) Petch, P.bambusicola C.L. Zhao and P.strigosozonata (Schwein.) P.H.B. Talbot.
Taxonomy
Conferticium
albocremeum
Taxon classificationFungiRussulalesStereaceae
L. Wang & C.L. Zhao sp. nov.
4A61C852-B8C1-514E-B3C8-39DA9CEBFA49
856958
Typification.
China. Yunnan Province • Xishuangbanna, Wild Elephant Valley, GPS coordinates: 22°10′N, 100°51′E, altitude: 900 m asl., on the fallen angiosperm branch, leg. C.L. Zhao, 25 January 2024 CLZhao 35693 (SWFC!).
Etymology.
Albocremeum (Lat.) refers to the new species having white to cream hymenophore.
Basidiomata.
Annual, resupinate, closely adnate, membranaceous, without odor or taste when fresh, up to 10 cm long, 2 cm wide, and 700 μm thick. Hymenophore smooth, white (60) to cream (4A2/3) when fresh, cream (4A2/3) upon drying. Sterile margin narrow, cream (4A2/3), up to 1 mm.
Basidiomata of Conferticiumalbocremeum (holotype CLZhao 35693). Scale bars: 1 cm (A); 1 mm (B).
Hyphal system.
Monomitic; generative hyphae with simple-septa, colorless, thin-walled, smooth, rarely branched, interwoven, IKI+, CB–, 2–3 µm in diameter; tissues unchanged in KOH.
Microscopic structures of Conferticiumalbocremeum (holotype CLZhao 35693): basidiospores (A); basidia and basidioles (B); gloeocystidia (C); a section of the hymenium (D). Scale bars: 10 µm (A–D).
Hymenium.
Gloeocystidia subclavate, flexuous, colorless, mostly constricted in the middle, thin-walled, smooth, 33–47.5 × 5–8 µm. Basidia subcylindrical to subclavate, slightly flexuous, with a basal simple septum and four sterigmata, 22–36 × 4.5–7 µm; basidioles numerous, in shape similar to basidia.
Sections of hymenium of Conferticiumalbocremeum (holotype CLZhao 35693): basidiospores (A); basidia (B); basidioles (C); gloeocystidia (D); a section of the generative hyphae (E). Scale bars: 10 µm (A–E); 10 × 100.
Spores.
Basidiospores ellipsoid with a distinct apiculus, colorless, thin-walled, finely verrucose but appearing smooth by light microscope, IKI+, CB–, 9–11 × 5–7 µm, L = 9.82 µm, W = 5.84 µm, Q = 1.36–1.68 (n = 90/3).
Additional specimens examined (paratypes).
China. Yunnan Province • Dehong, Mang City, Mengga Town, Tongbiguan Provincial Nature Reserve, GPS coordinates: 24°46′N, 97°34′E, altitude: 1300 m asl., on the fallen angiosperm branch, leg. C.L. Zhao, 29 June 2024, CLZhao 36343 • Dehong, Yingjiang County, Tongbiguan Provincial Nature Reserve, GPS coordinates: 25°50′N, 97°36′E, altitude: 1000 m asl., on the fallen angiosperm branch, leg. C.L. Zhao, 2 July 2024, CLZhao 37176 • Tengchong, Tuantian Town, Gaoligongshan National Nature Reserve, GPS coordinates: 25°27′N, 98°46′E, altitude: 2500 m asl., on the fallen angiosperm branch, leg. C.L. Zhao, 7 July 2024, CLZhao 39283 (SWFC!).
Dendrocorticiopsis
luteoalba
Taxon classificationFungiCorticialesPunctulariaceae
L. Wang & C.L. Zhao sp. nov.
3D3B5305-96D0-5FC0-AEDB-262A3FA24824
856959
Typification.
China. Yunnan Province • Dehong, Yingjiang County, Tongbiguan Provincial Nature Reserve, GPS coordinates: 25°50′N, 97°36′E, altitude: 1000 m asl., on the fallen angiosperm branch, leg. C.L. Zhao, 19 July 2023, CLZhao 30380 (SWFC!).
Etymology.
Luteoalba (Lat.) refers to the new species having a white to buff hymenophore.
Basidiomata.
Annual, resupinate, closely adnate, membranaceous, without odor or taste when fresh, up to 6 cm long, 2 cm wide, and 300 μm thick. Hymenophore smooth, white (60) when fresh, white (60) to buff (4A4) upon drying. Sterile margin narrow, white (60) to buff (4A4), up to 1 mm.
Basidiomata of Dendrocorticiopsisluteoalba (holotype CLZhao 30380). Scale bars: 1 cm (A); 1 mm (B).
Hyphal system.
Monomitic; generative hyphae with clamp connections, colorless, thin- to slightly thick-walled, smooth, branched, interwoven, usually with crystal masses, IKI–, CB–, 2.5–4 µm in diameter; tissues unchanged in KOH.
Microscopic structures of Dendrocorticiopsisluteoalba (holotype CLZhao 30380): basidiospores (A); basidia and basidioles (B); dendrohyphidia (C); cystidia (D); gloeocystidia (E); a section of the hymenium (F). Scale bars: 10 µm (A–F).
Hymenium.
Cystidia are of two types: (1) capitate, apically with resinous materials, gradually dissolving in KOH, colorless, thin-walled, smooth, 8.5–14 × 6.5–8.5 µm; (2) gloeocystidia, clavate to subulate, slightly flexuous, colorless, thin-walled, smooth, 68.5–90 × 8–10 µm. Dendrohyphidia numerous, thick-walled toward base, colorless, 16–19 × 2.5–3.5 µm. Basidia subclavate to clavate, flexuous, with a basal clamp connection and four sterigmata, 15.5–28.5 × 4–5.5 µm; basidioles numerous, in shape similar to basidia.
Sections of hymenium of Dendrocorticiopsisluteoalba (holotype CLZhao 30380): basidiospores (A); basidia and basidioles (B); dendrohyphidia (C); gloeocystidia (D); cystidia (E); a section of generative hyphae (F). Scale bars: 10 µm (A–F); 10 × 100.
Spores.
Basidiospores ellipsoid, colorless, thin-walled, smooth, IKI–, CB–, (6.5–)7–8 × (4–)4.5–5.5 µm, L = 7.29 µm, W = 4.97µm, Q = 1.47 (n = 30/1).
Eichleriella
biluoxueshanensis
Taxon classificationFungiAuricularialesAuriculariaceae
L. Wang & C.L. Zhao sp. nov.
BF299467-08A4-55BE-B9A4-D984347DF251
856960
Typification.
China. Yunnan Province • Diqing, Weixi County, Weideng, Songpo, GPS coordinates: 27°05′N, 99°13′E, altitude: 1400 m asl., on the fallen angiosperm branch, leg. C.L., 13 November 2023, CLZhao 34516 (SWFC!).
Etymology.
Biluoxueshanensis (Lat.) refers to the locality (Biluoxueshan) of the holotype specimen.
Basidiomata.
Annual, resupinate, closely adnate, coriaceous, without odor or taste when fresh, up to 8 cm long, 2 cm wide, and 700 μm thick. Hymenophore grandinioid, cream (4A2/3) to buff (4A4) when fresh, buff (4A4) to cinnamon-buff (4/5B4) upon drying. Sterile margin narrow, white (60) to cream (4A2/3), up to 1 mm.
Basidiomata of Eichleriellabiluoxueshanensis (holotype CLZhao 34516). Scale bars: 1 cm (A); 1 mm (B).
Hyphal system.
Dimitic; generative hyphae simple-septate, colorless, thin-walled, smooth, rarely branched, interwoven, 3–4 µm in diameter; skeletal hyphae distinctly thick-walled, smooth, unbranched, interwoven, IKI–, CB–, 3–4 µm in diameter; tissues unchanged in KOH.
Microscopic structures of Eichleriellabiluoxueshanensis (holotype CLZhao 34516): basidiospores (A); basidia and basidioles (B); cystidia (C); hyphidia (D); a section of the hymenium (E). Scale bars: 10 µm (A–E).
Hymenium.
Cystidia subclavate, flexuous, colorless, thin-walled, smooth, 40–45.5 × 6–9.5 µm. Basidia narrowly ovoid to ellipsoid, longitudinally septate, four-celled, 21–29 × 11.5–15 µm; basidioles numerous, in shape similar to basidia but smaller. Hyphidia arising from generative hyphae, nodulose, branched, colorless, thin-walled, 58–72.5 × 2.5–4 µm in diameter.
Sections of hymenium of Eichleriellabiluoxueshanensis (holotype CLZhao 34516): basidiospores (A); basidia (B); basidioles (C); cystidia (D); hyphidia (E); a section of the hymenium (F). Scale bars: 10 µm (A–F); 10 × 100.
Spores.
Basidiospores allantoid, colorless, thin-walled, smooth, usually with one or more oil drops, IKI–, CB–, (12–)13.5–17.5(–18) × (6.5–)7–9(–9.5) µm, L = 15.57 µm, W = 7.99 µm, Q = 1.95–2.06 (n = 60/2).
Additional specimen examined (paratype).
China. Yunnan Province • Diqing, Weixi County, Zhonglu, Lagaluo, GPS coordinates: 27°9′N, 99°8′E, altitude: 1710 m asl., on the fallen angiosperm branch, leg. C.L. Zhao, 10 October 2023, CLZhao 34793 (SWFC!).
Gloeohypochnicium
yunnanense
Taxon classificationFungiRussulalesGloeohypochnicium
L. Wang & C.L. Zhao sp. nov.
18083A14-BC5B-5C0A-AE73-0C1449626429
856961
Typification.
China. Yunnan Province • Dehong, Yingjiang County, Tongbiguan Provincial Nature Reserve, GPS coordinates: 25°50′N, 97°36′E, altitude: 1000 m asl., on the dead bamboo, leg. C.L. Zhao, 18 July 2023, CLZhao 30049 (SWFC!).
Etymology.
Yunnanense (Lat.) refers to the locality “Yunnan Province” of the holotype specimen.
Basidiomata.
Annual, resupinate, closely adnate, coriaceous, without odor or taste when fresh, up to 5 cm long, 3 cm wide, and 600 μm thick. Hymenophore smooth, slightly buff (4A4) when fresh, buff (4A4) to cream (4A2/3) upon drying. Sterile margin narrow, white (60) to buff (4A4), up to 1 mm.
Basidiomata of Gloeohypochniciumyunnanense (holotype CLZhao 30049). Scale bars: 1 cm (A); 1 mm (B).
Hyphal system.
Monomitic; generative hyphae with clamp connections, colorless, thick-walled, smooth, branched, interwoven, 1.5–4 µm in diameter; IKI–, CB–, tissues unchanged in KOH.
Microscopic structures of Gloeohypochniciumyunnanense (holotype CLZhao 30049): basidiospores (A); basidia and basidioles (B); cystidia (C); a section of the hymenium (D). Scale bars: 10 µm (A–D).
Hymenium.
Cystidia subcylindrical to subulate, flexuous, thin-walled, encrusted with whitish to yellowish crystals, 75–115.5 × 11.5–15 µm. Basidia subcylindrical to clavate, slightly flexuous, a basal clamp connection and four sterigmata, 55.5–70 × 9.5–11 µm; basidioles numerous, in shape similar to basidia but smaller.
Sections of hymenium of Gloeohypochniciumyunnanense (holotype CLZhao 30049): basidiospores (A); basidia (B); basidioles (C); gloeocystidia (D). Scale bars: 20 µm (A–D); 10 × 100.
Spores.
Basidiospores globose, colorless, thick-walled, warted, IKI–, CB–, (9.5–)10–12 × (9.5–)10–11.5 µm, L = 10.91 µm, W = 10.45 µm, Q = 1.04–1.05 (n = 60/2).
Additional specimen examined (paratype).
China. Yunnan Province • Dehong, Yingjiang County, Tongbiguan Provincial Nature Reserve, GPS coordinates: 27°52′N, 97°38′E, altitude: 1000 m asl., on dead bamboo, leg. C.L. Zhao, 18 July 2023, CLZhao 30018 (SWFC!).
Punctularia
nigrodontea
Taxon classificationFungiCorticialesPunctulariaceae
L. Wang & C.L. Zhao sp. nov.
374F9664-0CFA-54A3-9106-FB44261E2DDA
856962
Typification.
China. Yunnan Province • Dehong, Yingjiang County, Tongbiguan Provincial Nature Reserve, GPS coordinates: 25°50′N, 97°36′E, altitude: 1000 m asl., on the angiosperm trunk, leg. C.L. Zhao, 20 July 2023, CLZhao 30592 (SWFC!).
Etymology.
Nigrodontea (Lat.) refers to the new species having black basidiomata.
Basidiomata.
Annual, resupinate to effused-reflexed, adnate but easily separable, gelatinous, without odor or taste when fresh, up to 7 cm long, 3 cm wide, and 600 μm thick. Pileal surface smooth, rigid, fuscous (5/6F5) when fresh, fuscous (5/6F5) to black (51) upon drying; pileal back cushion-shaped grandinioid, rigid, black (51) when fresh, black (51) upon drying. Sterile margin narrow, black (51), up to 1 mm.
Basidiomata of Punctularianigrodontea (holotype CLZhao 30592). Scale bars: 1 cm (A, B); 1 mm (C, D).
Hyphal system.
Monomitic; generative hyphae clamp connections, colorless, thin to thick-walled, smooth, rarely branched, interwoven, 3–7 µm in diameter; IKI–, CB–, tissues unchanged in KOH.
Microscopic structures of Punctularianigrodontea (holotype CLZhao 30592): basidiospores (A); basidia and basidioles (B); a section of the hymenium (C). Scale bars: 10 µm (A–C).
Hymenium.
Cystidia absent. Basidia clavate, flexuous, with a basal clamp connection and four sterigmata, 16–25 × 3–4.5 µm; basidioles numerous, in shape similar to basidia but smaller.
Sections of hymenium of Punctularianigrodontea (holotype CLZhao 30592): basidiospores (A); basidia and basidioles (B); a section of thin-walled generative hyphae (C); a section of thick-walled generative hyphae (D). Scale bars: 10 µm (A–D); 10 × 100.
Spores.
Basidiospores ellipsoid, colorless, thin-walled, smooth, IKI–, CB–, 8.5–10(–10.5) × (4.5–)5–6(–6.5) µm, L = 9.24 µm, W = 5.47 µm, Q = 1.69 (n = 30/1).
Discussion
For fungal groups that are difficult to identify based on their morphological features, it is believed (and in most cases also proved) that the application of DNA sequences is able to delimit/recognize species much more easily and unequivocally (Wu et al. 2020; Bhunjun et al. 2024; He et al. 2022, 2024; Hyde et al. 2024a, b; Zhao et al. 2024; Zhou et al. 2024). Over time, understanding different aspects of fungi (i.e., taxonomy, diversity, species number) has improved rapidly by incorporating molecular and bioinformatics tools with traditional approaches (Cui et al. 2019; Wang et al. 2021; Hyde et al. 2023; Zhou et al. 2023; Dong et al. 2024a; Wang et al. 2024a; Wijayawardene et al. 2024).
Phylogenetically, the phylogenetic tree (Figs 1, 2) inferred from ITS+nLSU sequences revealed that the new species Conferticiumalbocremeum was nested into the family Stereaceae within the order Russulales, in which it was retrieved as a sister to C.heimii. However, C.heimii is distinguished from C.albocremeum by having a grayish orange hymenial surface and smaller basidiospores (5.3–6.2 × 3.5–4.2 µm; Wu 1996). Morphologically, C.fissuratum and C.ravum are similar to C.albocremeum by having finely verrucose, ellipsoid, and thin-walled basidiospores (Bernicchia and Gorjón 2010; Shen et al. 2024). However, C.fissuratum differs in its tuberculate, rough, white to cream, and cracking hymenial surface and longer subcylindrical cystidia (37–54.5 × 4–8 µm; Shen et al. 2024), and C.ravum differs in its smooth, brownish orange to grayish orange hymenial surface and bigger gloeocystidia (30–60 × 6–15 µm; Bernicchia and Gorjón 2010).
Phylogenetically, the multiple genes with ITS+nLSU analysis (Fig. 3) showed that the new species Dendrocorticiopsisluteoalba was nested into the family Punctulariaceae within the order Corticiales, and it is closely related with D.orientalis. Morphologically, D.luteoalba is similar to D.orientalis by having clavate apically with resinous cystidia and clavate to subclavate basidia (Wei et al. 2022). However, D.orientalis is delimited from D.luteoalba by its finely cracked, grayish ivory hymenial surface and smaller, ellipsoid to ovoid basidiospores (5–7 × 3.2–5.2 µm; Wei et al. 2022).
The phylogenetic tree (Figs 4, 5) inferred from ITS+nLSU sequences revealed that Eichleriellabiluoxueshanensis grouped into the genus Eichleriella and clustered into the family Auriculariaceae, in which it was grouped with the clade comprising E.sinensis. However, E.sinensis is distinguished from E.biluoxueshanensis by having narrower basidiospores (10.5–16 × 5.5–7 µm; Li et al. 2023). Morphologically, E.xinpingensis C.L. Zhao and E.yunnanensis Y.L. Deng & C.L. Zhao are similar to E.biluoxueshanensis by all having subcylindrical to allantoid, thin-walled, and smooth basidiospores (Liu et al. 2019; Deng et al. 2024a). However, E.xinpingensis differs in its soft, leathery to ceraceous, flesh-pink to clay-pink, and covered by blunt-pointed spines hymenial surface and shorter basidia (15–28 × 5–9 µm; Liu et al. 2019), and E.yunnanensis differs in its cream to flesh-pink hymenial surface and smaller basidiospores (7.5–11.5 × 3.5–5 µm; Deng et al. 2024a).
The phylogenetic tree (Fig. 1) inferred from ITS+nLSU sequences revealed that Gloeohypochniciumyunnanense grouped into the genus Gloeohypochnicium and clustered into the order Russulales, in which it was closely related to G.analogum. Morphologically, G.yunnanense is similar to G.analogum in that it has subglobose, thick-walled, and warted basidiospores (Bernicchia and Gorjón 2010). However, G.analogum is delimited from G.yunnanense by its coriaceous, cream to ochraceous hymenial surface with a fibrillose margin and smaller gloeocystidia (40–60 × 6–10 µm; Bernicchia and Gorjón 2010).
Based on the ITS+nLSU sequence data (Fig. 3), Punctularianigrodontea was nested into the family Punctulariaceae within the order Corticiales and grouped with the clade comprising P.atropurpurascens, P.bambusicola, and P.strigosozonata. Morphologically, P.nigrodontea is similar to P.atropurpurascens, P.bambusicola, and P.strigosozonata by having smooth, thin-walled, and ellipsoid basidiospores (Bernicchia and Gorjón 2010; Guan et al. 2021). However, P.atropurpurascens is delimited from P.nigrodontea by having the effuse-reflexed and reddish-brown to dark purplish-brown or bluish hymenial surface and larger basidia (40–65 × 5–6 µm; Guan et al. 2021). Punctulariabambusicola differs in its resupinate, tuberculate with rose tints, pink to purple hymenial surface, and smaller basidiospores (6.5–8.5 × 3.5–5 µm; Guan et al. 2021). Punctulariastrigosozonata differs in its resupinate to effuse-reflexed basidiomata with a brown, velutinous margin and longer basidia (40–60 × 4–5 µm; Bernicchia and Gorjón 2010).
Fungi are an ancient, diverse, and heterogeneous group of organisms; they can be found in a wide range of habitats, and play key roles in ecosystems as decomposers, mutualists, and pathogens (Dai et al. 2015, 2021; Cui et al. 2023; Wei 2021; Bhunjun et al. 2022). The Yunnan Province is rich in woody plant species, providing excellent substrates for wood-inhabiting fungi (Dong et al. 2023, 2024a; Wang and Cai 2023; Deng et al. 2024b; Wang et al. 2024b; Zhu et al. 2024). Our study is helpful to further understand the species diversity of wood-inhabiting fungal groups in Yunnan and explore their evolutionary relationships.
Supplementary Material
XML Treatment for Conferticium albocremeum
XML Treatment for Dendrocorticiopsis luteoalba
XML Treatment for Eichleriella biluoxueshanensis
XML Treatment for Gloeohypochnicium yunnanense
XML Treatment for Punctularia nigrodontea
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alvarenga RLM Gibertoni TB (2021) Crystallodon Alvarenga gen. nov., a new genus of the Auriculariales from the Neotropics. Cryptogamie.Mycologie 42(2): 17–24. 10.5252/cryptogamie-mycologie 2021 v 42a 2 · doi ↗
- 2Alvarenga RLM Spirin V Malysheva V Gibertoni TB Larsson KH (2019) Two new genera and six other novelties in Heterochaete sensu lato (Auriculariales, Basidiomycota).Botany 97(8): 439–451. 10.1139/cjb-2019-0046 · doi ↗
- 3Bernicchia A Gorjón SP (2010) Fungi Europaei 12: Corticiaceae s.l.. Edizioni Candusso, Italia.
- 4Bhunjun CS Niskanen T Suwannarach N Wannathes N Chen YJ Mc Kenzie EH Maharachchikumbura SS Buyck B Zhao CL Fan YG Zhang JY Dissanayake AJ Marasinghe DS Jayawardena RS Kumla J Padamsee M Chen Y-Y Liimatainen K Ammirati JF Phukhamsakda C Liu J-K Phonrob W RandrianjohanyÉHongsanan S Cheewangkoon R Bundhun D Khuna S Yu W-J Deng L-S Lu Y-Z Hyde KD Lumyong S (2022) The numbers of fungi: Are the most speciose genera truly diverse? Fungal Diversity 114(1): 387–462. 10.1007/s 13225-022-00501-4 · doi ↗
- 5Bhunjun CS Chen YJ Phukhamsakda C Boekhout T Groenewald JZ Mc Kenzie EHC Francisco EC Frisvad JC Groenewald M Hurdeal VG Luangsa-ard J Perrone G Visagie CM Bai FYBłaszkowski J Braun Ude Souza F Ade Queiroz MB Dutta AK Gonkhom D Goto BT Guarnaccia V Hagen F Houbraken J Lachance MA Li JJ Luo KY Magurno F Mongkolsamrit S Robert V Roy N Tibpromma S Wanasinghe DN Wang DQ Wei DP Zhao CL Aiphuk W Ajayi-Oyetunde O Arantes TD Araujo JC Begerow D Bakhshi M Barbosa RN Behrens FH Bensch K Bezerra JDP Bilański P Bradley CA Bubner B Burgess TI Buyck BČadežN Cai L Calaça FJS Campbell LJ Chaverri P Chen YY Chethana KWT Co · doi ↗ · pubmed ↗
- 6Cui BK Li HJ Ji X Zhou JL Song J Si J Yang ZL Dai YC (2019) Species diversity, taxonomy and phylogeny of Polyporaceae (Basidiomycota) in China.Fungal Diversity 97(1): 137–392. 10.1007/s 13225-019-00427-4 · doi ↗
- 7Cui BK Pan XH Pan F Sun YF Xing JH Dai YC (2023) Species diversity and resources of Ganoderma in China.Junwu Xuebao 42(1): 170–178.
- 8Dai YC Cui BK Si J He SH Hyde KD Yuan HS Liu XY Zhou LW (2015) Dynamics of the worldwide number of fungi with emphasis on fungal diversity in China.Mycological Progress 14(8): 62. 10.1007/s 11557-015-1084-5 · doi ↗