Schwann cell deletion of Tumor Susceptibility Gene 101 ( Tsg101 ) in mice results in severe peripheral neuropathy
Derek Silvius, Edward Hurley, Yannick Poitelon, Kay-Uwe Wagner, M. Laura Feltri, Teresa M. Gunn

TL;DR
Deleting a specific gene in nerve cells causes severe nerve damage in mice, highlighting its role in healthy nerve function.
Contribution
This study shows that TSG101 is essential for myelination in the peripheral nervous system.
Findings
Deleting Tsg101 in Schwann cells causes severe peripheral neuropathy in mice.
TSG101 is required for normal myelination in the peripheral nervous system.
The gene plays a key role in endosomal trafficking in Schwann cells.
Abstract
Myelinating Schwann cells are particularly susceptible to defects in endosomal trafficking. TSG101 is a component of the endosomal trafficking machinery that mediates the sorting of ubiquitinated receptors into multivesicular bodies. We previously demonstrated that deleting Tsg101 from mouse oligodendrocytes in the central nervous system causes rapid onset de/dys-myelination and vacuolation of white matter, suggesting an important role for TSG101-dependent trafficking in myelination. Here, we show that TSG101 is also required for normal myelination in the peripheral nervous system.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
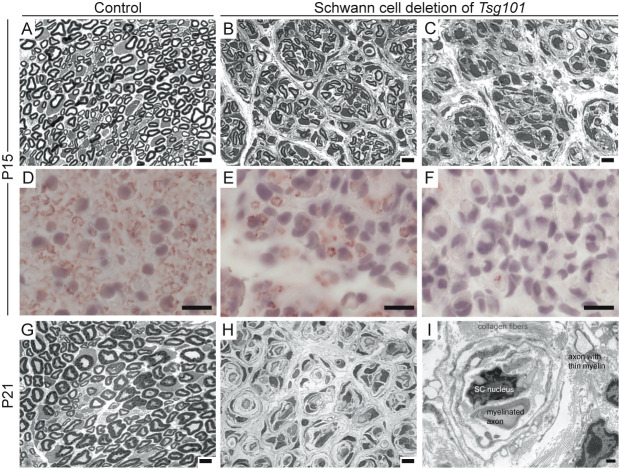 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeuroblastoma Research and Treatments
Description
The endosomal pathway traffics receptor proteins and lipids into early endosomes and directs them either to recycling endosomes for trafficking back to the cell membrane or sorts them into intraluminal vesicles (ILVs) within multivesicular bodies (MVBs), which fuse with lysosomes to degrade their contents or with the plasma membrane to secrete ILVs as exosomes (reviewed in (Scott et al., 2014) ). The timing of these events is important to cellular processes since receptors can continue to activate downstream signaling pathways while they remain on early endosomes. Several forms of demyelinating CMT are caused by mutations in genes encoding proteins involved in membrane dynamics and endosomal trafficking, including N-myc downstream regulated gene 1 ( NDRG1), myotubularin related protein 2 ( MTMR2), SET binding factor 2 (SBF2/MTMR13),SH3 domain and tetratricopeptide repeats 2 ( *SH3TC2), dynamin 2 (DNM2), FIG4 phosphoinositide 5-phosphatase * ( FIG 4), and LPS-induced TN factor ( LITAF/SIMPLE ) (reviewed in (Markworth et al., 2021) ). For example, mutations in SH3TC2 that cause CMT type 4C cause impaired recycling of membrane components necessary for Schwann cell function (Roberts et al., 2010; Stendel et al., 2010) . Furthermore, mouse models with Schwann cell-specific deletion of Fig4 , HGF-regulated tyrosine kinase substrate ( Hgs/Hrs), or phosphatidylinositol 3-kinase catalytic subunit type 3 ( Pik3c3) show mild peripheral hypo/de/dys-myelination associated with altered ERBB2/3 signaling (Logan et al., 2017; McLean et al., 2022; Vaccari et al., 2015) . In Schwann cells, neuregulin 1 (NRG1) signaling through erb-b2 receptor tyrosine kinases 2 and 3 (ERBB2/B3) mediates myelination, and endosomal trafficking of NRG1-bound ERBB2/B3 regulates receptor down-regulation and recycling (Newbern and Birchmeier, 2010; Salzer, 2015) . Disrupted endosomal sorting can result in sustained activation of downstream pathways, including the ERK1/2 signaling cascade, to negatively impact Schwann cell myelin integrity (Newbern and Birchmeier, 2010; Salzer, 2015) . Thus, there is support for the idea that endosomal trafficking defects can cause Schwann cell dysfunction and demyelination, reinforcing the relevance of this pathway to myelination.
Tumor susceptibility gene 101 (TSG101) encodes a component of the E ndosomal S orting C omplex R equired for T ransport-1 (ESCRT-I), which helps mediate the sorting of ubiquitinated receptors onto ILVs of MVBs. The ESCRT-0 protein, HGF-regulated tyrosine kinase substrate, HGS (formerly referred to as HRS) mediates the initial recruitment of ESCRT-I to endosomes (via interaction with TSG101), but in multiple mammalian cell lines, siRNA depletion of TSG101 or HGS had significantly different effects on endosomal and MVB morphology (Bache et al., 2003; Lu et al., 2003; Raiborg et al., 2008; Razi and Futter, 2006) . Specifically, knockdown of TSG101 inhibited epidermal growth factor degradation and MVB formation and caused tubulation of the vacuolar domains of early endosomes, while depletion of HGS had only a modest effect on EGF degradation, did not induce tubulation of early endosomes, and resulted in the production of enlarged MVBs containing few ILVs but that could still fuse with the lysosome (Bishop et al., 2002; Doyotte et al., 2005; Razi and Futter, 2006) . These data suggest that HGS and TSG101 have distinct roles in the endosomal trafficking pathway, with TSG101 being required for the formation of stable vacuolar domains within the early endosome that subsequently develop into MVBs and HGS being more important in the formation and/or accumulation of ILVs within MVBs. Mice lacking HGS in Schwann cells developed mild motor and sensory defects, a reduced number of myelinated axons and thinner myelin sheaths in the sciatic nerve, as well as aberrantly folded myelin sheaths (McLean et al., 2022) .
TSG101 and HGS also both interact and partially colocalize with LITAF (Lee et al., 2011) , which is expressed in Schwann cells and mutations in it cause dominant demyelinating peripheral neuropathy, CMT1C (Bennett et al., 2004) . Although one study showed that CMT1C-associated LITAF mutations did not effect on its subcellular localization or association with TSG101 (Shirk et al., 2005) , another study showed a dominant negative effect on EGFR degradation and lysosomal trafficking of EGF, associated with reduced membrane association of HGS and TSG101 (Lee et al., 2011) . In the latter study, expression of CMT1C-associated LITAF mutants in Schwann cells caused prolonged activation of ERK1/2 signaling, presumably downstream of NRG1-ERBB2/3 signaling.
Deleting Tsg101 from oligodendroglia in the central nervous system resulted in severe, rapid-onset myelination defects and vacuolation (Walker et al., 2016) , suggesting an important role for TSG101-dependent trafficking in signaling pathways that regulate myelination. To test whether TSG101 is also required for normal myelination in the peripheral nervous system, we investigated the consequences of deleting Tsg101 in Schwann cells. We predicted this would cause a more severe peripheral neuropathy than deleting Hgs, given the stronger effect of Tsg101 depletion on endosomal/MVB phenotypes in cultured cells.
Tsg101 conditional knockout mice ( * Tsg101 ^tm1KuW^ * , referred to here as * Tsg101 ^fl^ * ) were mated to P0-Cre transgenic mice, which express cre recombinase specifically in Schwann cells starting on embryonic day 13.5. Cre-positive * Tsg101 ^fl/+^ * offspring were backcrossed to * Tsg101 ^fl/fl^ * animals. All pups that were homozygous for the Tsg101 conditional allele and carried the P0-Cre transgene (referred to herein as Tsg101 -Schwann cell null, or * Tsg101 ^SC-null^ * animals) were smaller than their littermates, developed a tremor by 12 days of age, had abnormal posture of their fore- and hind-limbs (arthrogryposis), and failed to thrive. They died throughout the postnatal period, with very few surviving to 3 weeks of age. We recorded 89 affected animals out of 427 pups born from 15 different breeder pairs, for a frequency of ~21% affected. This is significantly different from the 25% expected (χ ^2^ = 4.04, p = 0.044) and is likely due to the loss of some affected animals prior to them being observed and recorded.
Histological analysis of the sciatic nerves of * Tsg101 ^SC-null^ * animals at postnatal days 15 and 21 revealed striking de- and dys-myelination ( Fig. 1 ). Toluidine blue-stained semi-thin cross sections showed reduced myelin, enlargement of the interstitial space, and presence of onion bulb structures in the sciatic nerves of * Tsg101 ^SC-null^ * mice by postnatal day 15 (P15, Fig. 1A-C ). The sciatic nerves of some * Tsg101 ^SC-null^ * animals had fewer, thinner myelin sheaths, while others showed an almost complete absence of myelin, as detected by toluidine blue staining on semithin sections ( Fig. 1A-C ) and immunohistochemistry for myelin basic protein (MBP) on paraffin sections ( Fig. 1D-F ). The myelination defects associated with loss of TSG101 were progressive, with the sciatic nerve of a P21 * Tsg101 ^SC-null^ * animal showing severe hypomyelination and presence of onion bulb formations, (Fig. G-H). Ultrathin electron micrograph (EM) analysis of P21 sciatic nerve cross-sections revealed axons with thin myelin sheaths and multiple cell layers adjacent to nucleus of Schwann cells. These “onion bulbs” comprise concentric layers of Schwann cell processes and connective tissue (collagen) arranged around axons and are consistent with multiple rounds of de- and remyelination (Iwata et al., 1998; Tracy et al., 2019; Webster et al., 1967) . This suggests that TSG101 is not essential for myelin production, but it is required for the maintenance of stable myelin sheaths. Schwann cell nuclei were still present by P21 and did not appear pyknotic ( Fig. 1F ), suggesting that loss of TSG101 did not disrupt myelination by triggering Schwann cell apoptosis.
As predicted, based on the differences observed in cellular phenotypes and EGF/EGFR degradation when TSG101 or HGS was depleted from mammalian cells by siRNA, the phenotype of * Tsg101 ^SC-null^ * mice was more severe than that observed in * Hgs ^SC-null^ * mice. Onion bulb formations are characteristic features in CMT1A and other demyelinating CMTs. Since TSG101 and ESCRT proteins are critical for endosomal sorting, their dysfunction could impair myelin production and turnover, exacerbating the cycles of demyelination and remyelination that contribute to onion bulb pathology. In the future, proteomic studies may shed light on the specific signaling pathways disrupted in * Tsg101 ^SC-null^ * that contribute to the severe peripheral neuropathy phenotype. The variable expressivity of myelination defects and survival of * Tsg101 ^SC-null^ * mice likely reflects their mixed genetic background (129S1/Sv x C57BL/6) and an effect of modifier genes, although inter-animal differences in P0-cre expression and Tsg101 deletion cannot be ruled out. Identifying the genes and pathways that influence disease severity in these mice could reveal druggable targets to treat some forms of CMT or other peripheral neuropathies.
Methods
Animals
All studies were approved by the McLaughlin Research Institute’s Institutional Animal Care and Use Committee and adhered to the Association for Assessment and Accreditation of Laboratory Animal Care guidelines. Mice homozygous for a Tsg101 conditional knockout allele (129X1/SvJ- * Tsg101 ^tm1Kuw^ * , referred to herein as * Tsg101 ^fl^ * ; RRID: MMRRC_037407-MU), in which exon 1 is flanked by a floxed neo cassette inserted approximately 3 kb upstream and a single loxP site inserted 230 bp into intron 1 (Wagner et al., 2003) , were mated to mice hemizygous for Tg(Mpz-cre)26Mes/J (obtained from the Jackson Laboratory and referred to herein as P0-cre; RRID: IMSR_JAX:017928). P0-cre transgenic mice express cre recombinase in Schwann cells, under the control of the myelin protein zero (P 0 , Mpz ) promoter, by embryonic day 14 (Feltri et al., 1999) . * Tsg101 ^fl/+^ ; P0-cre+ * F1 offspring were backcrossed to * Tsg101 ^fl/fl^ * mice and * Tsg101 ^fl/fl^ ; P0-cre+ * and control ( * Tsg101 ^fl/+^ ; P0-cre+ * and * Tsg101 ^fl/fl^ ; P0-cre *
neg ) pups were obtained from intercrossing N1 or N1F1 Tsg101fl/+; P0-cre+ x * Tsg101 ^fl/fl^ ; P0-cre-neg * sibs.
Histology
Sciatic nerve segments were sampled from approximately the same position for each animal. Contralateral nerves were either fixed in 4% buffered paraformaldehyde, for immunohistochemistry (IHC), or 2% buffered glutaraldehyde with postfixation in 1% osmium tetroxide (for semi- and ultra-thin sections).
For IHC, specimens from five * Tsg101 ^SC-null^ * and four control animals were embedded in paraffin following standard protocols and sectioned at 5 microns. Following deparaffinization and rehydration, sections were permeabilized in a 0.2% Triton-X100 solution in PBS, then subjected to antigen retrieval in 10mM sodium citrate (pH 6.0, 100C, 10 minutes). Slides were blocked in 10% serum, then incubated with an antibody against myelin basic protein (MBP; Covance Cat# SMI-99, RRID:AB_2314772) at 1:1000, followed by horseradish peroxidase conjugated anti-mouse secondary antibody (BD Pharmingen horseradish peroxidase-conjugated anti-mouse Ig, cat# 554002, RRID: AB_395198) at 1:100 and chromogenic visualization using NovaRed substrate (Vector Labs, CA). Slides were counterstained with hematoxylin prior to coverslipping, and then examined on a Zeiss AxioImagerM1 light microscope.
Routine semithin and ultrathin (electron micrograph [EM]) analyses were performed as described (Quattrini et al., 1996) on sciatic nerves from * Tsg101 ^SC-null^ * animals (n=2 at P15, n=1 at P21) and controls (n=1-2 at each age). Briefly, sciatic nerves were fixed in 2% buffered glutaraldehyde, then postfixed in 1% osmium tetroxide. After alcohol dehydration, nerves were submerged in propylene oxide, and then in a 1:1 mixture of Epon-propylene oxide. Nerves were embedded in 100% Epon, and resin was allowed to polymerize. Semithin transverse sections were sliced 0.5-μm-thick using Leica UC7, stained with 2% toluidine blue, and then examined by light microscopy with Leica DM6000B. EM transverse sections were sliced 700–900 Å-thick using Leica UC7, stained with uranyl acetate and lead citrate, and then examined with an electron microscope (model FEI BioTwin). Analyzed sections were sliced from the distal end of embedded sciatic nerve. Images acquired from semithins and EMs were nonoverlapping and comprehensive.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bache Kristi G. Brech Andreas Mehlum Anja Stenmark Harald 200384 Hrs regulates multivesicular body formation via ESCRT recruitment to endosomes The Journal of Cell Biology 16231540-814043544210.1083/jcb.20030213112900395 PMC 2172703 · doi ↗ · pubmed ↗
- 2Bennett Craig L. Shirk Andrew J. Huynh Huy M. Street Valerie A. Nelis Eva Van Maldergem Lionel De Jonghe Peter Jordanova Albena Guergueltcheva Velina Tournev Ivailo Van den Bergh Peter Seeman Pavel Mazanec Radim Prochazka Tomas Kremensky Ivo Haberlova Jana Weiss Michael D. Timmerman Vincent Bird Thomas D. Chance Phillip F. 2004426 SIMPLE mutation in demyelinating neuropathy and distribution in sciatic nerve Annals of Neurology 5550364-513471372010.1002/ana.2009415122712 · doi ↗ · pubmed ↗
- 3Bishop Naomi Horman Alistair Woodman Philip 2002326 Mammalian class E vps proteins recognize ubiquitin and act in the removal of endosomal protein–ubiquitin conjugates The Journal of Cell Biology 15711540-81409110210.1083/jcb.20011208011916981 PMC 2173266 · doi ↗ · pubmed ↗
- 4Doyotte Aurelie Russell Matthew R. G. Hopkins Colin R. Woodman Philip G. 2005715 Depletion of TSG 101 forms a mammalian `Class E' compartment: a multicisternal early endosome with multiple sorting defects Journal of Cell Science 118141477-91373003301710.1242/jcs.0242116014378 · doi ↗ · pubmed ↗
- 5Feltri ML D'Antonio M Previtali S Fasolini M Messing A Wrabetz L 1999914 P 0-Cre transgenic mice for inactivation of adhesion molecules in Schwann cells.Ann N Y Acad Sci 8830077-892311612310586237 · pubmed ↗
- 6Iwata Atsushi Kunimoto Masanari Inoue Kiyoharu 199841 Schwann cell proliferation as the cause of peripheral neuropathy in neurofibromatosis-2Journal of the Neurological Sciences 15620022-510X 20120410.1016/s 0022-510x(98)00032-x 9588858 · doi ↗ · pubmed ↗
- 7Lee SM Olzmann JA Chin LS Li L 201196 Mutations associated with Charcot-Marie-Tooth disease cause SIMPLE protein mislocalization and degradation by the proteasome and aggresome-autophagy pathways.J Cell Sci 124Pt 190021-95333319333110.1242/jcs.08711421896645 PMC 3178453 · doi ↗ · pubmed ↗
- 8Logan Anne M. Mammel Anna E. Robinson Danielle C. Chin Andrea L. Condon Alec F. Robinson Fred L. 2017615 Schwann cell‐specific deletion of the endosomal PI 3‐kinase Vps 34 leads to delayed radial sorting of axons, arrested myelination, and abnormal Erb B 2‐Erb B 3 tyrosine kinase signaling Glia 6590894-14911452147010.1002/glia.2317328617998 PMC 5818149 · doi ↗ · pubmed ↗