Two new species of the genus Malaxa Melichar, 1914 from China (Hemiptera, Fulgoromorpha, Delphacidae, Tropidocephalini)

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
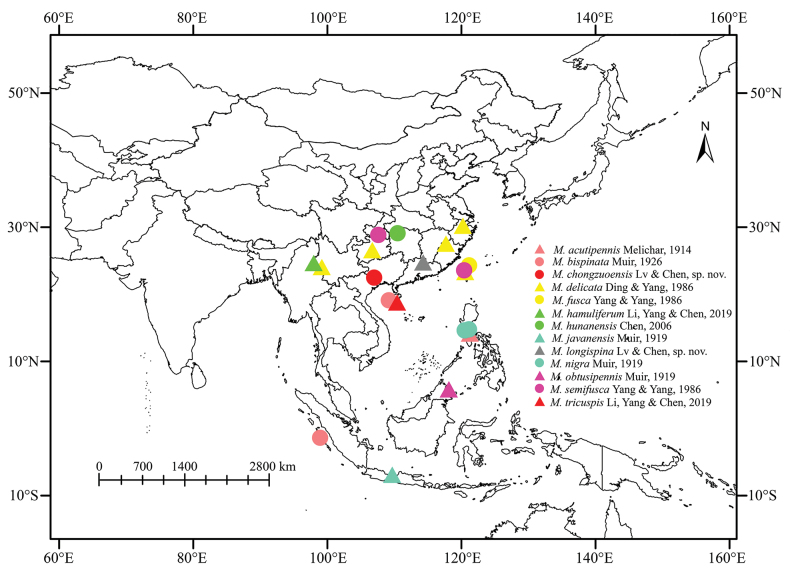 Figure 1
Figure 1 Figures 2–5
Figures 2–5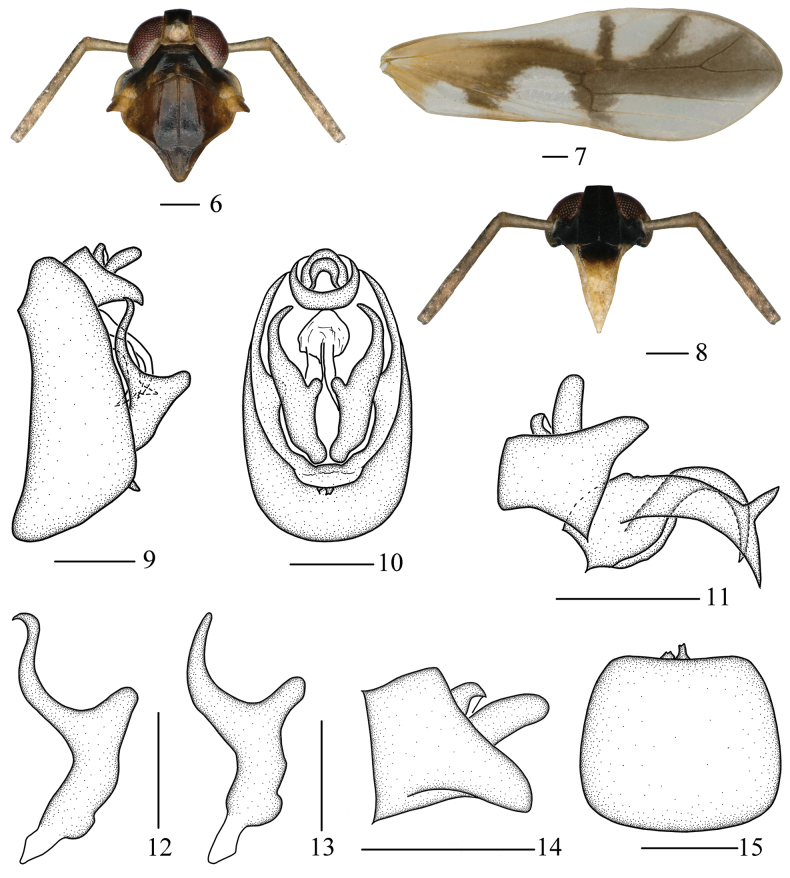 Figures 6–15
Figures 6–15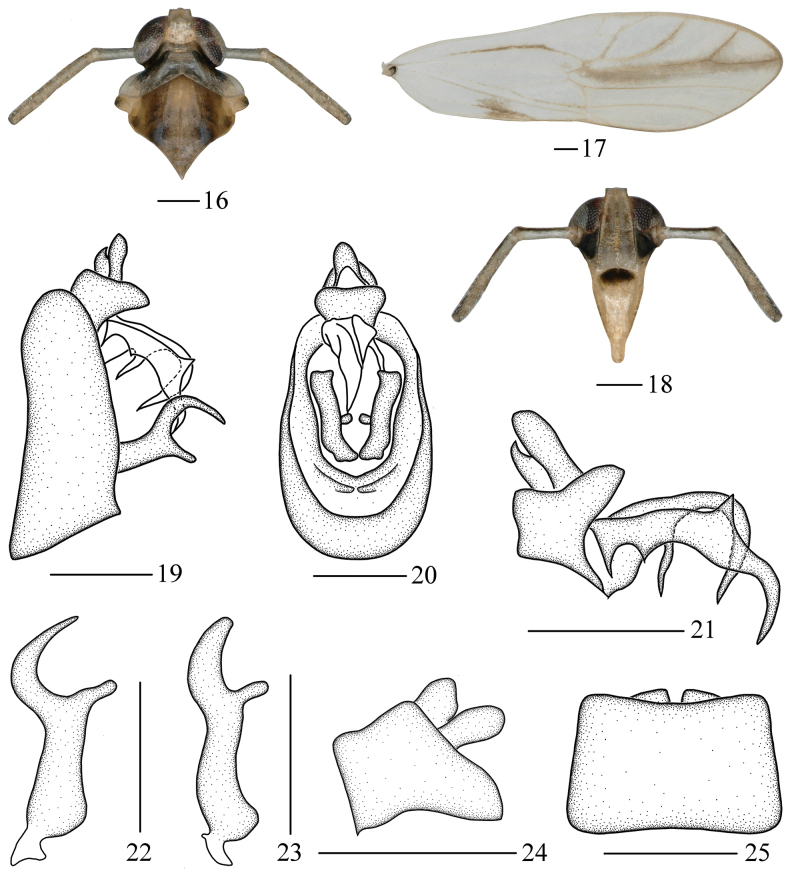 Figures 16–25
Figures 16–25 Figure 26
Figure 26 Figure 27
Figure 27| 1 | Postclypeus yellow; forewings with apical veins CuA and MP3 diverging apically, posterior half of apical forewings dark brown ( |
|
| – | Postclypeus with basal half blackish-brown; forewings with apical veins CuA and MP3 fused, first and second apical cells hyaline |
|
| 2 | Anal segment without process; aedeagus with phallobase |
|
| – | Anal segment with a long process; aedeagus without phallobase |
|
| 3 | Genae all dark brown; forewings with basal half most yellow, apical half most dark brown |
|
| – | Genae dark brown in part; forewings mostly hyaline |
|
| 4 | Forewings (Fig. |
|
| – | Forewings with basal 1/2 bearing a hyaline V-shaped marking, fifth apical cell dark brown; dorsal margin of phallobase without a slender horned process ( |
|
| 5 | Vertex with apical half and basal half of frons dark brown; middle part of apical half of forewings without grayish-brown markings; pygofer with three medioventral processes very distinct ( |
|
| – | Vertex and frons (Figs |
|
| 6 | Genae dark brown; in posterior view, process of anal segment situated in middle of ventral margin ( |
|
| – | Genae mostly dark brown but apical with small part yellow; in posterior view, process of anal segment situated on left side of ventral margin |
|
| 7 | Gonostyles with apex not forked; aedeagus with three processes ( |
|
| – | Gonostyles with apex forked; aedeagus with two processes |
|
| 8 | Area between lateral carinae of pronotum dark brown; two branches of outer apical angle of gonostyles subequal; aedeagus with a small spine situated near basal third, directed caudally ( |
|
| – | Area between lateral carinae of pronotum mostly yellow; two branches of outer apical angle of gonostyles unequal; aedeagus with a small tooth situated near middle, directed right ( |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhytoplasmas and Hemiptera pathogens · Insect-Plant Interactions and Control · Plant Virus Research Studies
Introduction
Melichar (1914) established the delphacid planthopper genus Malaxa with the type species M.acutipennis Melichar, 1914 from the Philippines and placed this genus in the tribe Tropidocephalini of the subfamily Delphacinae (Hemiptera: Fulgoromorpha: Delphacidae) (Muir 1915, 1916). Until now, 11 species have been recorded in the genus (Bartlett and Kennedy 2018; Li et al. 2019; Bourgoin 2024). In turn, Muir (1919, 1926) described four new species, M.javanensis Muir, 1919 and M.bispinata Muir, 1926 from Indonesia, M.nigra Muir, 1919 from Philippines, and M.obtusipennis Muir, 1919 from Malaysia. Ding et al. (1986) and Yang and Yang (1986) added three new species from China, M.delicata Ding & Yang, 1986, M.fusca Yang & Yang, 1986 and M.semifusca Yang & Yang, 1986. Chen et al. (2006) and Hou et al. (2013) both reviewed the Chinese species of Malaxa, and added a new species, M.hunanensis Chen, 2006. Bartlett and Kennedy (2018) reviewed the New World species attributed to the genus, redescribed the type species M.acutipennis Melichar, 1914, and transferred M.occidentalis Muir, 1926 and M.gracilis Fennah, 1945 to Lamaxa Bartlett & Kennedy, 2018, and M.microstyla Muir, 1930 to Xalama Bartlett & Kennedy, 2018. Li et al. (2019) described two new species, M.hamuliferum Li, Yang & Chen, 2019 and M.tricuspis Li, Yang & Chen, 2019 from China. This genus is known to occur in the Oriental region, with seven species in China, two species in the Philippines, two species in Indonesia, and one species in Malaysia (Melichar 1914; Muir 1919, 1926; Ding et al. 1986; Yang and Yang 1986; Chen et al. 2006; Ding 2006; Hou et al. 2013; Li et al. 2019; Bourgoin 2024).
Here we describe and illustrate two new species of Malaxa discovered in southern China, and provide a checklist of species, a distribution map, and an updated key to Chinese species.
Material and methods
The external morphology terminologies follow Bourgoin (1987) for male genitalia and Bourgoin et al. (2015) for wing venation. The zoogeographic regionalization scheme follows Holt et al. (2013). Specimens were collected by sweeping. Dry male specimens were used for the descriptions and illustrations. Body length was measured from the apex of the vertex to the tip of the forewings. All measurements are in millimeters (mm). Color pictures for adult habitus were taken using the KEYENCE VHX-6000 system. External morphology and drawings were done with a Leica MZ 12.5 stereomicroscope. The photographs and illustrations were scanned with a CanoScan LiDE 200 and imported into Adobe Photoshop v. 6.0 for labeling and plate composition. The genital segments of the examined specimens were macerated in 10% NaOH, and then transferred to glycerol in small plastic tubes pinned together with the specimens for examination. The distribution map was generated with ArcGIS v. 10.7.
The type specimens examined are deposited in the Institute of Entomology, Guizhou University, Guiyang, Guizhou Province, China (IEGU).
Taxonomy
Malaxa
Taxon classificationAnimaliaHemipteraDelphacidae
Melichar, 1914
C0792920-7DFF-5D89-B47E-5781E963BDF7
Malaxa Melichar, 1914: 275; Muir 1926: 7; Metcalf 1943: 103; Fennah 1945: 429; Yang and Yang 1986: 56; Ding et al. 1986: 418, 1999: 443; Chen et al. 2006: 160; Ding 2006: 150; Bartlett 2009: 387; Hou et al. 2013: 866; Bartlett and Kennedy 2018: 514; Li et al. 2019: 44.
Type species.
Malaxaacutipennis Melichar, 1914, original designation.
Diagnosis. For the diagnosis of Malaxa see Hou et al. (2013: 866) and Li et al. (2019: 44).
Host plants.
Bamboo (Poales: Poaceae: Bambusoideae).
Distribution.
China, Indonesia, Malaysia, Philippines (Fig. 1).
Geographic distributions of species of Malaxa Melichar, 1914.
Checklist and distributions of species of Malaxa Melichar, 1914
M.acutipennis Melichar, 1914; Philippines (Luzón)
M.bispinata Muir, 1926; Indonesia (Mentawai), China (Hainan)
M.chongzuoensis Lv & Chen, sp. nov.; China (Guangxi)
M.delicata Ding & Yang, 1986; China (Fujian, Guizhou, Taiwan, Yunnan, Zhejiang)
M.fusca Yang & Yang, 1986; China (Taiwan)
M.hamuliferum Li, Yang & Chen, 2019; China (Yunnan)
M.hunanensis Chen, 2006; China (Hunan)
M.javanensis Muir, 1919; Indonesia (Java)
M.longispina Lv & Chen, sp. nov.; China (Guangdong)
M.nigra Muir, 1919; Philippines (Luzón)
M.obtusipennis Muir, 1919; Malaysia (Borneo)
M.semifusca Yang & Yang, 1986; China (Guizhou, Taiwan)
M.tricuspis Li, Yang & Chen, 2019; China (Hainan)
Key to Chinese species of Malaxa Melichar, 1914
(modified from Li et al. 2019)
**: **
Malaxa
chongzuoensis
Taxon classificationAnimaliaHemipteraDelphacidae
Lv & Chen sp. nov.
498C4CA5-B9D9-5C0C-A6D0-55AF901A5E41
https://zoobank.org/BB6FCADA-56F5-4854-94F3-16AA5FA5F82E
Type material.
Holotype: China • ♂; Guangxi Zhuang Autonomous Region, Chongzuo City, Longzhou County, Zhubu Township, Nonggang Village; 22°39'N, 106°57'E; sweeping, 16 August 2024; Sha-Sha Lv and Xiang-Sheng Chen leg.; IEGU. Paratypes: China • 1♂, 4♀♀; same collection data as for holotype; IEGU.
Diagnosis.
The salient features of the new species include: vertex (Figs 2, 6) with apical half brownish-black; frons and genae (Figs 3, 8) black; mesonotum (Figs 2, 6) blackish-brown at middle, rest tawny to reddish-brown; forewings (Fig. 7) with a reversed hyaline V-shaped marking; pygofer (Fig. 15) in ventral view medioventral processes asymmetrical; outer process of gonostyles (Fig. 12) snakelike in lateral view; dorsal margin of phallobase (Fig. 11) with a slender horned process at apical 1/3.
Description.
Measurements. Total length: male 3.5–3.7 mm (N = 2), female 4.1–4.5 mm (N = 4).
Coloration. General color pale yellowish-brown (Figs 2, 3). Vertex (Figs 2, 6) with apical half brownish-black, basal half yellowish-brown. Frons (Fig. 8) and genae (Figs 3, 8) black. Clypeus (Fig. 8) with basal half brownish-black. Eyes (Figs 6, 8) reddish-brown. Pronotum (Figs 2, 6) brown to black except lateral sides yellow. Mesonotum (Figs 2, 6) blackish-brown at middle, rest tawny to reddish-brown. Outer part of tegulae (Figs 2, 6) black brown, inner part yellowish-white. Forewings (Fig. 7) greyish-white, hyaline, veins gray to light yellowish-brown, basal 1/4 light yellowish-brown except areas around bifurcation of Pcu and A1, basal 1/4 to middle part with an arched dark brown stripe, forming a reverse hyaline V-shaped marking, along ScP, ir, RP and area between MP_1_ and MP_2_ dark brown.
Head and thorax. Vertex (Fig. 6) slightly longer than wide at base (1.07: 1), width at apex narrower than at base (0.83: 1), submedian carinae uniting slightly beyond middle, apex produced in front of eyes, apical margin straight, greatest length of basal compartment shorter than wide at base of vertex (0.67: 1). Frons (Fig. 8) longer in middle line than wide at widest portion (about 1.28: 1), widest at apex, median carina simple. Postclypeus (Fig. 8) wide at base as wide as frons at apex. Antennae (Figs 6, 8) very long, cylindrical, surpassing apex of clypeus, scape longer than wide, shorter than pedicel (0.46: 1). Pronotum (Fig. 6) with lateral carinae not attaining hind margin, longer than vertex in midline (0.75: 1). Mesonotum (Fig. 6) with lateral carinae not attaining hind margin, longer than 1.67 times pronotum and vertex combined. Forewings (Fig. 7) slender, longer than maximal width (3.02: 1).
Male genitalia. Pygofer in lateral view (Fig. 9) ventral margin longer than dorsal margin, in posterior view (Fig. 10) with opening longer than wide, in ventral view (Fig. 15) medioventral processes asymmetrical, concave medially, left process shorter. Gonostyles (Figs 12, 13) long, in lateral view apical half bifurcated into two processes, outer process slender and curved, snakelike, tapering to apex, inner process short and thick, rounded at apex; in posterior view C-shaped. Male genitalia (Fig. 11) with phallobase, aedeagus tubular, vaulted ventrally, tapering to apex; phallobase wide, curved ventrally, tapering to apex, dorsal margin with a slender horned process at apical 1/3. Anal segment (Figs 9, 14) small, ring-like.
Malaxachongzuoensis Lv & Chen, sp. nov., male 2 habitus, dorsal view 3 habitus, lateral view 4, 5Malaxalongispina Lv & Chen, sp. nov., male 4 habitus, dorsal view 5 habitus, lateral view. Scale bars: 0.5 mm (2–5).
Malaxachongzuoensis Lv & Chen, sp. nov., male 6 head and thorax, dorsal view 7 forewing 8 frons, ventral view 9 male genitalia, lateral view 10 male genitalia, posterior view 11 anal segment and aedeagus, lateral view 12 gonostyle, lateral view 13 gonostyle, posterior view 14 anal segment, lateral view 15 pygofer, ventral view. Scale bars: 0.2 mm (6–15).
Host plant.
Indocalamustessellatus (Munro) P. C. Keng (Poales: Poaceae: Bambusoideae) (Fig. 26).
Distribution.
China (Guangxi) (Fig. 1).
Etymology.
The new species is named after the city in which it was collected (Chongzuo) with the Latin adjectival suffix ‘-ensis’ meaning ‘from’.
Remarks.
This species (Figs 6–15) is similar to M.hamuliferum Li, Yang & Chen, 2019 (Li et al. 2019: figs 5–14), but differs from the latter in: (1) forewings with basal 1/2 bearing a reversed hyaline V-shaped marking (forewings with basal 1/2 bearing a hyaline V-shaped marking in M.hamuliferum); (2) dorsal margin of phallobase with a long horned process (dorsal margin of phallobase without a long horned process in M.hamuliferum); and (3) aedeagus shorter than phallobase (aedeagus longer than phallobase in M.hamuliferum).
Malaxa
longispina
Taxon classificationAnimaliaHemipteraDelphacidae
Lv & Chen sp. nov.
EB7B8BB8-4C98-5F87-97A6-21FA6C929CCD
https://zoobank.org/5CB9A6AB-56C4-4064-A999-3086A7DC9918
Type materials.
Holotype: China • ♂: Guangdong Province, Shixing County, Luoba Town, Dashui Village; 24°46'N, 114°17'E; sweeping, 18 June 2023; Sha-Sha Lv leg.; IEGU. Paratypes: China • 1♂, 3♀♀; Guangdong Province, Shixing County, Luoba Town, Dashui Village; 24°46'N, 114°17'E; sweeping, 18 June 2023; Sha-Sha Lv, Feng-E Li and Yong-Jin Sui leg.; IEGU.
Diagnosis.
The salient features of the new species include: vertex (Figs 4, 16) and frons (Fig. 18) light grayish-yellow; genae (Figs 5, 18) with basal 3/4 black markings; pronotum (Figs 4, 16) with outer sides of lateral carinae with black markings; forewings (Fig. 17) around ScP+R, ir, RP and MP_1_ with light greyish-brown marking; medioventral process of pygofer (Fig. 25) nearly rectangular in ventral view; gonostyles (Fig. 22) with apical half forming a C-shaped in lateral view; aedeagus (Fig. 21) narrows sharply near the middle, with a slender process.
Description.
Measurements. Total length: male 3.3–3.4 mm (N = 2), female 4.1–4.3 mm (N = 3).
Coloration. General color greyish-white to yellowish-brown (Figs 4, 5). Vertex (Figs 4, 16) and frons (Fig. 18) light grayish-yellow, frons with two indistinct light brown stripes. Genae (Figs 5, 18) with basal 3/4 black markings. Clypeus (Fig. 18) black at base, and rest light yellowish-brown. Eyes (Figs 16, 18) gray to reddish-black. Pronotum (Figs 4, 16) light grayish-brown, outer sides of lateral carinae with black markings. Mesonotum (Figs 4, 16) yellowish-brown, outer sides of lateral carinae with brown to black markings. Apex of tegulae (Figs 4, 16) black brown, rest light yellowish-brown. Forewings (Fig. 17) greyish-white, hyaline, veins gray to brown, areas after bifurcation of Pcu and A1 with yellowish-brown spot, around ScP+R, ir, RP and MP_1_ with light greyish-brown marking.
Head and thorax. Vertex (Fig. 16) shorter submedially than wide at base (0.96: 1), width at apex narrower than at base (0.62: 1), submedian carinae uniting slightly beyond middle, apex produced in front of eyes, apical margin straight, greatest length of basal compartment shorter than wide at base of vertex (0.59: 1). Frons (Fig. 18) longer in middle line than wide at widest portion (about 2.17: 1), widest at apex, median carina simple. Postclypeus (Fig. 18) wide at base as wide as frons at apex. Antennae (Figs 16, 18) very long, cylindrical, surpassing apex of clypeus, scape longer than wide, shorter than pedicel (0.55: 1). Pronotum (Fig. 16) with lateral carinae not attaining hind margin, longer than vertex in midline (0.71: 1). Mesonotum (Fig. 16) with lateral carinae not attaining hind margin, longer than 1.61 times pronotum and vertex combined. Forewings (Fig. 17) slender, longer than maximal width (3.63: 1).
Malaxalongispina Lv & Chen, sp. nov., male 16 head and thorax, dorsal view 17 forewing 18 frons, ventral view 19 male genitalia, lateral view 20 male genitalia, posterior view 21 anal segment and aedeagus, lateral view 22 gonostyle, lateral view 23 gonostyle, posterior view 24 anal segment, lateral view 25 pygofer, ventral view. Scale bars: 0.2 mm (16–25).
Male genitalia. Pygofer in lateral view (Fig. 19) ventral margin longer than dorsal margin, ventral angles slightly produced, in posterior view (Fig. 20) with opening longer than wide, in ventral view (Fig. 25) medioventral processes nearly rectangular, concave medially. Gonostyles (Figs 22, 23) moderately long, in lateral view apical half bifurcated into two processes, forming a C-shaped, outer process long and bent, tapering to apex, inner process short, relatively rounded at apex; in posterior view curved, basal angles produced, outer process thick. Male genitalia (Fig. 21) with phallobase, aedeagus tubular, vaulted ventrally, basal part broad, narrows sharply near the middle, with a slender spinous process; phallobase long, curved ventrally, tapering to apex, basal part with two spiniform processes, dorsal margin with an angular process medially. Anal segment (Figs 19, 24) small, ring-like.
Adult of Malaxachongzuoensis Lv & Chen, sp. nov. resting on the leaf of Indocalamustessellatus (Munro) P. C. Keng (Poales: Poaceae: Bambusoideae). Photographed by Xiang-Sheng Chen.
Host plant.
Indocalamus sp. (Poales: Poaceae: Bambusoideae).
Distribution.
China (Guangdong) (Fig. 1).
Etymology.
The species name is a combination of the Latin word “long-” and “spina” (with the connecting letter “i”, i.e., “long spine”), referring to the ventral margin of the aedeagus with a long spinous process medially. The name is intended to be feminine.
Remarks.
This species (Figs 16–25) is similar to M.bispinata Muir, 1926 (Hou et al. 2013: figs 1–14), but differs from the latter in: (1) vertex and frons grayish-white (vertex with apical half and basal half of frons dark brown in M.bispinata); (2) middle part of apical half of forewings with grayish-brown markings (middle part of apical half of forewings without grayish-brown markings in M.bispinata); and (3) pygofer with two medioventral processes not distinct (pygofer with three medioventral processes very distinct in M.bispinata).
Discussion
Most members of the Tropidocephalini tribe with documented plant associations primarily feed on bamboo (Poales: Poaceae: Bambusoideae), as reported by Wilson et al. (1994), where this association accounts for 78% of the records. The remaining species are associated with various grasses (e.g., Wilson et al. 1994; Chen 2003; Chen and Tsai 2009; Qin and Zhang 2010). Specifically, species of Malaxa from China that have reported plant associations also feed on bamboo. Specimens have been collected from the leaves of several bamboo genera, including Bambusa, Indocalamus, Fargesia, and Phyllostachys (Yang and Yang 1986; Chen et al. 2006; Hou et al. 2013). We have also collected these two species of Malaxa from bamboo. These species may pose potential threats as pests in bamboo forests.
Based on geographic distribution (Fig. 1), all species of the genus Malaxa are found in the Oriental Realm (Holt et al. 2013), with a particularly rich diversity in China, where nine species are currently recorded. However, the genus is primarily distributed in Central China, South China, and Southwest China, with most species known only from their type localities. We believe that the actual distributions of many species remain unclear, as our findings are predominantly based on type localities, indicating a need for more extensive and in-depth investigations.
Forewings of Chinese Malaxa species AM.bispinata Muir, 1926 BM.chongzuoensis Lv & Chen, sp. nov. CM.delicata Ding & Yang, 1986 DM.fusca Yang & Yang, 1986 EM.hamuliferum Li, Yang & Chen, 2019 FM.hunanensis Chen, 2006 GM.longispina Lv & Chen, sp. nov. HM.semifusca Yang & Yang, 1986 IM.tricuspis Li, Yang & Chen, 2019.
Bartlett and Kennedy (2018) redescribed the type species M.acutipennis Melichar, 1914. They observed several differences between M.acutipennis and the Chinese species in the genus Malaxa, the most salient difference is that M.acutipennis has an apically pointed forewing, while the forewings of all other Malaxa species are rounded. Here, we illustrate the forewings of all Chinese species of this genus, as shown in Fig. 27. We find that these forewings are also slightly different when extending towards the apex; some are relatively slender and apex slightly pointed, but the differences are not as pronounced as in the type species. Therefore, the study of this genus still requires more species, and it is highly necessary to expand the collection area for further collection.
Supplementary Material
XML Treatment for Malaxa
XML Treatment for Malaxa chongzuoensis
XML Treatment for Malaxa longispina
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bartlett CR (2009) A new genus of New World Tropidocephalini (Hemiptera: Delphacidae: Delphacinae), with the description of two new species.Entomological News 120(4): 387–396. 10.3157/021.120.0407 · doi ↗
- 2Bartlett CR Kennedy AC (2018) A review of New World Malaxa (Hemiptera: Fulgoroidea: Delphacidae).Zootaxa 4441(3): 511–528. 10.11646/zootaxa.4441.3.530313998 · doi ↗ · pubmed ↗
- 3Bourgoin T (1987) A new interpretation of the homologies of the Hemiptera male genitalia, illustrated by the Tettigometridae (Hemiptera, Fulgoromorpha). Proceedings 6th Auchenorrhyncha Meeting, Turin, Italy, 7–11 September, 113–120.
- 4Bourgoin T (2024) FLOW (Fulgoromorpha Lists On the Web): A knowledge and a taxonomy database dedicated to planthoppers (Insecta, Hemiptera, Fulgoromorpha, Fulgoroidea). Version 8, updated 10 November 2024. https://flow.hemiptera-databases.org/flow/ [Accessed on: 1 December 2024]
- 5Bourgoin T Wang RR Asche M Hoch H Soulier-Perkins A Stroiński A Yap S Szwedo J (2015) From micropterism to hyperpterism recognition strategy and standardized homology-driven terminology.Zoomorphology 134(1): 63–77. 10.1007/s 00435-014-0243-625705075 PMC 4326643 · doi ↗ · pubmed ↗
- 6Chen XS (2003) Key to genera of the tribe Tropidocephalini (Hemiptera: Fulgoroidea: Delphacidae) from the People’s Republic of China, with description of a new genus.Canadian Entomologist 135(6): 811–821. 10.4039/n 02-097 · doi ↗
- 7Chen XS Tsai JH (2009) Two new genera of Tropidocephalini (Hemiptera: Fulgoroidea: Delphacidae) from Hainan Province, China.Florida Entomologist 92(2): 261–268. 10.1653/024.092.0210 · doi ↗
- 8Chen XS Li XF Liang AP Yang L (2006) A review of the bamboo delphacid genus Malaxa (Hemiptera: Fulgoroidea: Delphacidae) from China.Annales Zoologici 56(1): 159–166.