Machine Learning Predicts Non-Preferred and Preferred Vertebrate Hosts of Tsetse Flies (Glossina spp.) Based on Skin Volatile Emission Profiles
Olabimpe Y. Orubuloye, David P. Tchouassi, Abdullahi A. Yusuf, Christian W. W. Pirk, Daniel K. Masiga, Edward Kariuki, Baldwyn Torto

TL;DR
This study uses machine learning to predict which animals tsetse flies prefer to bite based on the chemicals they emit from their skin.
Contribution
The study introduces a machine learning approach to identify chemical signals that help tsetse flies distinguish between preferred and non-preferred hosts.
Findings
96 volatile organic compounds were detected in skin emissions, varying by vertebrate host.
Three repellent ketones plus background odors may help tsetse flies discriminate hosts.
Random forest analysis showed weak correlation between ketones and host preference alone.
Abstract
Tsetse fly vectors of African trypanosomosis preferentially feed on certain vertebrates largely determined by olfactory cues they emit. Previously, we established that three skin-derived ketones including 6-methyl-5-hepten-2-one, acetophenone and geranyl acetone accounted for avoidance of zebra by tsetse flies. Here, we tested the hypothesis that these three ketones serve as biomarkers for tsetse flies to distinguish between non-preferred- and preferred-vertebrate hosts. We used coupled gas chromatography/mass spectrometry to analyze and compare the skin volatile emissions of two non-preferred- (waterbuck and zebra) and four preferred- (buffalo, donkey, horse, warthog) vertebrate hosts in two wildlife parks in Kenya. We detected a total of 96 volatile organic compounds (VOCs) in the skin emissions composed mainly of aldehydes, ketones, alcohols, phenols and alkanes, which varied with…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
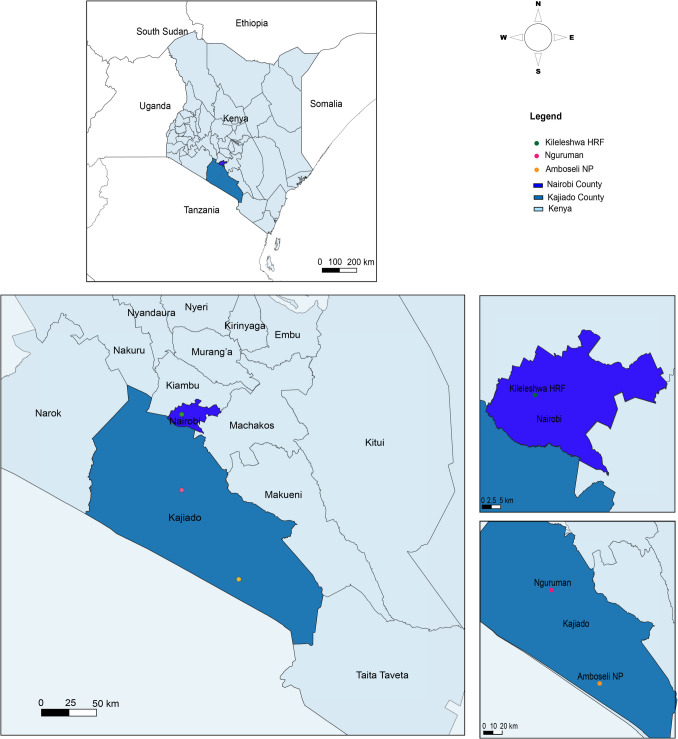 Figure 1
Figure 1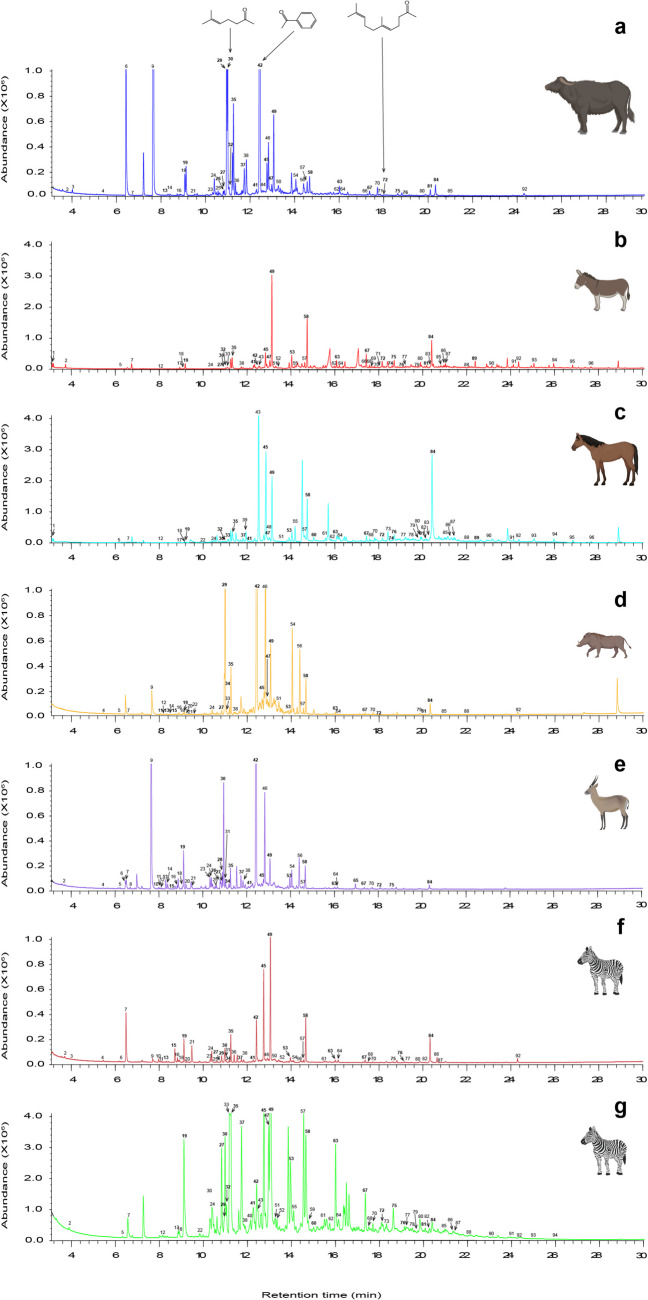 Figure 2
Figure 2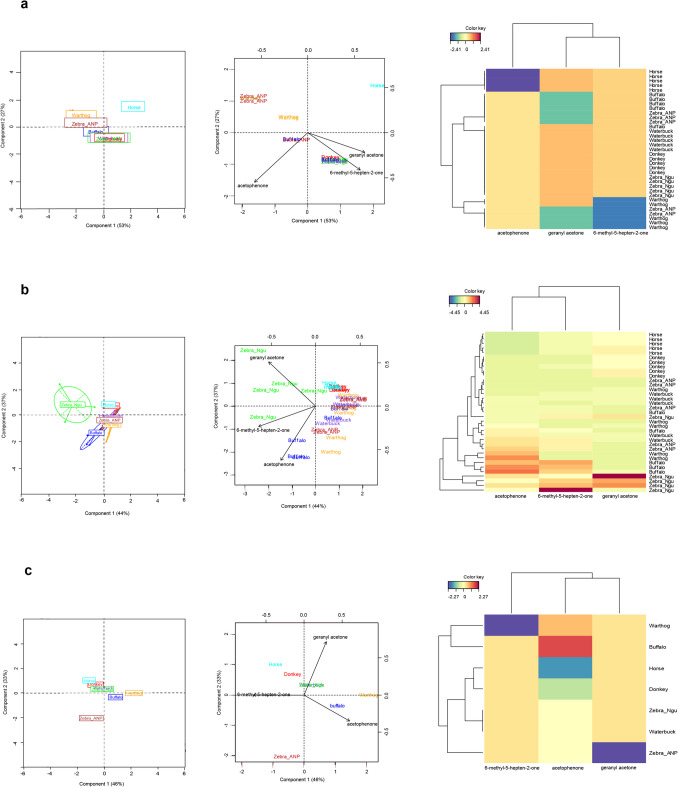 Figure 3
Figure 3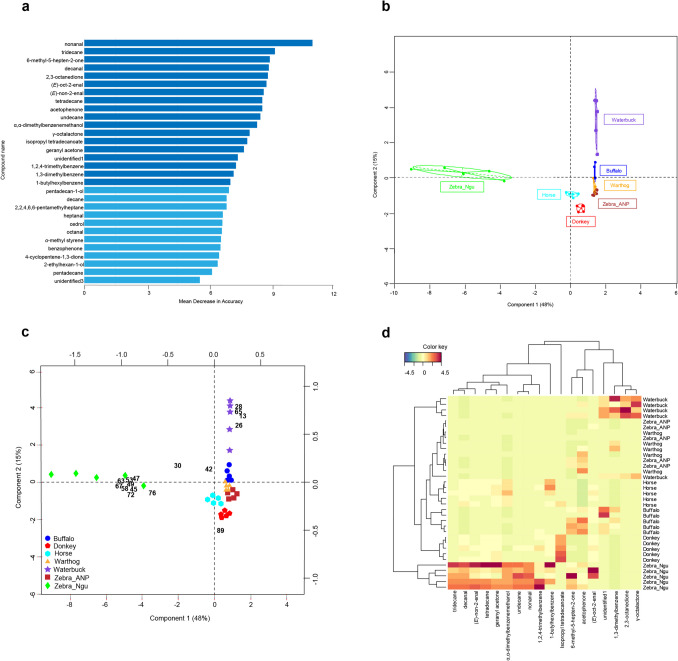 Figure 4
Figure 4- —European Union’s Integrated Biological Control Applied Research Programme – tsetse repellent component (EUIBCARP tsetse)
- —German Academic Exchange Service (DAAD)
- —https://doi.org/10.13039/501100001343University of Pretoria
- —UK’s Department for International Development (DFID)
- —https://doi.org/10.13039/501100000974Australian Centre for International Agricultural Research
- —Swedish International Development Cooperation Agency (Sida)
- —Swiss Agency for Development and Cooperation (SDC)
- —Norwegian Agency for Development Cooperation
- —German Federal Ministry for Economic Cooperation and Development
- —Government of the Republic of Kenya
- —University of Pretoria
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect and Pesticide Research · Mosquito-borne diseases and control · Insect and Arachnid Ecology and Behavior
Introduction
The Afro-endemic tsetse flies Glossina spp. (Diptera: Glossinidae) are obligate blood feeding insects and vectors of trypanosome pathogens causing African trypanosomosis, a devastating but neglected tropical disease. African trypanosomosis affects both humans and animals, particularly livestock, with 60 million and 50 million humans and cattle, respectively, at risk of infection in 37 sub-Saharan African countries (FAO 2024). While human African trypanosomosis is a declining public health concern (FAO and WHO 2022; Franco et al. 2024), animal African trypanosomosis still poses a constraint to sustainable agricultural and livestock production accounting for about three million cattle deaths and losses valued at USD 4.75 billion annually (Abro et al. 2023; FAO 2024; Muriithi et al. 2021; Shaw et al. 2017). There are 31 extant species and sub-species of tsetse flies within the Glossina species complex classified into three groups, namely: savannah or morsitans (subgenus Glossina s.s.), riverine or palpalis (subgenus Nemorhina) and forest or fusca (subgenus Austenina) group (Orubuloye et al. 2024; Vreysen et al. 2013), which differ in their habitat requirements, host preferences, epidemiological and economic importance.
Like other blood feeding insects, such as mosquitoes (Takken 1991) and sandflies (Tchouassi et al. 2024), tsetse flies also locate their hosts using a combination of olfactory and visual cues. Visual cues are important at close range for landing decisions (Gibson and Torr 1999; Vreysen et al. 2013), but tsetse flies principally employ olfactory cues to locate and discriminate vertebrate hosts for a blood meal (Gikonyo et al. 2000, 2003; Olaide et al. 2019; Takken and Knols 2010). Host-derived olfactory cues comprise kairomones (attractants) and allomones (repellents), mainly volatile organic compounds (VOCs) suited for long range communication. For example, the VOCs 3-n-propylphenol and 4-methylphenol (p-cresol) emanating from host urine, and 1-octen-3-ol and acetone from host breath are important kairomones for tsetse host location (Masiga et al. 2014; Rayaisse et al. 2010; Vale and Torr 2004). A four-component blend of these attractive chemicals combined with visual traps or targets form the basis of the bait technology, an effective and highly successful semiochemical-based control tool for savannah tsetse flies (Masiga et al. 2014).
Selective feeding on certain vertebrates by tsetse flies is well described in the literature (Auty et al. 2016; Channumsin et al. 2021; Clausen et al. 1998; Ebhodaghe et al. 2021; Gashururu et al. 2023; Makhulu et al. 2021; Moloo et al. 1993; Muturi et al. 2011) and driven mainly by semiochemicals which are now being exploited for their control (Bett et al. 2015; Gikonyo et al. 2003; Orubuloye et al. 2024; Saini et al. 2017). Generally, vertebrates like buffalo, cattle, warthog, elephant, giraffe are more frequently fed on by savannah tsetse flies compared to others such as waterbuck, zebra, wildebeest, impala and Thompson’s gazelle. For instance, in the Masai Mara National Reserve Kenya (Auty et al. 2016) and Serengeti National Park Tanzania (Makhulu et al. 2021) both in eastern Africa, the savannah tsetse flies G. pallidipes and G. swynnertoni preferentially fed on vertebrates such as African buffalo, warthog and elephant compared to the more abundant wildebeest and zebra. Similar feeding preferences have been reported for tsetse flies in areas of Kenya, Tanzania and Uganda in East Africa (Channumsin et al. 2021; Ebhodaghe et al. 2021; Muturi et al. 2011), Zambia in southern Africa (Gaithuma et al. 2020), and Rwanda, Central Africa (Gashururu et al. 2023). However, these feeding preferences could vary depending on the tsetse fly group. For instance, savannah tsetse flies are more selective than their riverine counterparts which are opportunistic feeders (Oloo et al. 2014; Torr and Vale 2015; Vale et al. 2014). Additionally, hunger status, environmental conditions (e.g. host availability), and species-specific preferences (Clausen et al. 1998; Gikonyo et al. 2000; Muturi et al. 2011; Orubuloye et al. 2024) may all contribute to the feeding preferences of tsetse flies.
Repellent chemicals emitted in the skin odors of non-preferred vertebrates but absent in the preferred ones, or found at sub-optimal amounts are important semiochemical drivers of tsetse fly feeding preferences (Gikonyo et al. 2002, 2003; Olaide et al. 2019). For example, a four-component tsetse repellent blend comprised of geranyl acetone, guaiacol, pentanoic acid and δ-octalactone and identified from the waterbuck protects livestock host (cattle) from tsetse bites and trypanosomosis infection (Bett et al. 2015; Saini et al. 2017). In another study, zebra skin odors, comprised of a blend of three ketones, 6-methyl-5-hepten-2-one, acetophenone, and geranyl acetone repelled two tsetse fly species Glossina pallidipes and G. fuscipes fuscipes (Olaide et al. 2019). Of the seven skin-derived repellents comprised of four classes of chemicals (ketone, phenol carboxylic acid and lactone), keto-compounds dominate (geranyl acetone, 6-methyl-5-hepten-2-one and acetophenone) with geranyl acetone being common to the skin-odors of waterbuck and zebra.
Hence, in this study, we tested the hypothesis that the zebra-derived ketones geranyl acetone, 6-methyl-5-hepten-2-one and acetophenone discriminate tsetse fly non-preferred and preferred hosts in two wildlife natural habitats in Kenya: Nguruman and Amboseli National Park. To achieve this, we screened skin volatile emission profiles and assessed the levels and ratios of the three ketones from selected tsetse fly non-preferred (waterbuck and zebra) and preferred (buffalo, donkey, horse, warthog) hosts. Data was further analyzed using machine learning algorithms to identify discriminatory skin odors among the vertebrates.
Methods
Study Area and Vertebrate Species
This study was conducted between April 2017 and May 2019, in Nguruman and Amboseli National Park both of which are in Kajiado County, Kenya (Fig. 1). Amboseli National Park (S 2° 38′ 29″ E 37° 14′ 53″) is a 392 km^2^ protected area and highly diverse ecosystem famous for its large herds of elephants and other key wildlife species like buffalo, zebra, wildebeest, giraffe, a variety of antelopes and predators, and many bird species (KWS 2024; UNESCO 2024). Nguruman (S 1° 54′ 26.1″ E 36° 46′ 56.5) also supports diverse wildlife species which commonly interact with livestock (especially cattle, goat, and sheep) due to the proximity of Maasai pastoralists. Both areas harbor the savannah tsetse fly species G. pallidipes; although Nguruman provides more favourable breeding habitat for this species and the forest species G. longipennis (Olaide et al. 2019; Ouma et al. 2006). The study areas were selected based on accessibility of the target vertebrates, and the presence of savannah tsetse flies which are selective blood feeders.Fig. 1. Map of Kenya showing the vertebrate skin odor collection areas. The maps were designed using the open-source software QGIS 3.20.3 “Odense” (https://www.qgis.org/download/). The coordinates of Nguruman, Amboseli National Park, and Kileleshwa horse riding farm (HRF) were obtained in the field using a GPS gadget (garmin etrex 20, https://buy.garmin.com/en-US/US/p/518046). The spatial data (Kenya, World) were downloaded from GADM (https://gadm.org/data.html), also an open source. Skin odors of zebra (Equus quagga) and donkey (Equus africanus asinus) were collected from Nguruman while waterbuck (Kobus ellipsiprymnus defassa), buffalo (Syncerus caffer), warthog (Phacochoerus africanus), and zebra (Equus quagga) were sampled from Amboseli, and horse (Equus ferus caballus) from Kileleshwa HRF in Nairobi, based on accessibility of the vertebrates in each location. For each vertebrate species, skin odor samples were collected from five adult individuals
Vertebrate species sampled included zebra (Equus quagga), waterbuck (Kobus ellipsiprymnus defassa), buffalo (Syncerus caffer), donkey (Equus africanus asinus), horse (Equus ferus caballus), and warthog (Phacochoerus africanus) were used in this study. Zebra and donkey were sampled in Nguruman, while waterbuck, buffalo, warthog and zebra were sampled in Amboseli, based on their accessibility in each location. Horse was not available in both localities and was sampled in Kileleshwa horse riding farm (S 1° 17′ 2.5″ E 36° 46′ 56.5″) in Nairobi, Kenya (Fig. 1). Donkey and horse ranged from 4–7 years old, however, ages of the wildlife species (zebra, waterbuck, buffalo, warthog) were unknown. Except for horses, individuals of each vertebrate species (five replicates) were sampled from different groups within the parks to capture possible diversity in skin volatile chemical profiles.
Skin Odor Collection and Trapping
Skin odor samples were collected from five adult individuals of each vertebrate species. Zebra, waterbuck and buffalo were darted in the wild before skin odour collection, as described in Olaide et al. (2019). Immobilization drug composed of opiate etorphine hydrochloride (Captivon® 98, Wildlife Pharmaceuticals Ltd, South Africa) and azaperone (Kyron Laboratories, South Africa) (Olaide et al. 2019; Walzer 2007). The vertebrates were located using a vehicle and immobilization drug safely delivered in a 3 ml dart with a plain 50 mm needle shot using a Dan-Inject CO_2_ Injection rifle (Dan-Inject APS, Denmark). After skin odor collection, anaesthesia was reversed using intravenous injection of opioid antagonist diprenorphine (Activon®, Wildlife Pharmaceuticals Ltd, South Africa) at a dose of 0.045 mg/kg body weight (Olaide et al. 2019; Walzer 2007). All individual vertebrates recovered, with no complications observed after antagonist administration. Warthogs were captured using a nylon capture net, and were restrained manually by trained capture rangers prior to collection of skin odors.
Skin odor collection and trapping protocol is as described in Olaide et al. (2019) and Tchouassi et al. (2013). Briefly, Soxhlet-extracted cotton materials (23 cm × 23 cm, Lux Premium, Bidhannagar, West Bengal, India) were rubbed on each vertebrate’s skin around the belly and upper parts of the front legs which are major focus areas for tsetse feeding, for 10—12 min, using latex-gloved hands. For each replicate, headspace odors (volatile organic compounds, VOCs) were immediately collected onsite from the cotton materials trapped onto two pre-packed adsorbent filters (Carbopak B 3.5″ with 30 mg ± 5 mg, Sigma Scientific, Gainesville, Florida, US). Odor trapping from cotton materials onto adsorbent filters was carried out for 12 h using a portable field pump (USDA/CMAVE, Gainesville, Florida, USA), fitted with a push–pull headspace volatile trapping system supplying charcoal-filtered air at a flow rate of 348 ml/min. Tightly sealed (0.075 mm P.T.F.E. thread seal tape MAAT, UK) Carbopak B adsorbents with trapped headspace volatiles were wrapped in aluminum foil and transported to the laboratories at icipe, Nairobi in a cool box underlaid with carbon dioxide CO_2_ (dry ice pellets, Carbacid Investment Limited, Nairobi, Kenya). Trapped skin odors were eluted from each adsorbent filter under laboratory conditions with 200 μl gas chromatography (GC) grade dichloromethane into 2 ml glass vials, concentrated to 100 μl under a gentle stream of charcoal filtered nitrogen gas and kept at −80 °C for GC/MS analysis.
Chemical Analysis
Skin odor samples were analyzed on a HP 7890 A series gas chromatograph (GC) tandem HP 5975 C mass spectrometer (MS) (Agilent Technologies, Wilmington, USA) fitted with an autosampler, a split-splitless injection port (200 ˚C), an Agilent HP-5MS non-polar capillary column (5% phenyl and 95% methylpolysiloxane, 30 m length × 250 μm i.d. × 0.25 μm film thickness), an Agilent technologies 5975C EIMS (70 eV) triple axis MS, and an Agilent ChemStation data system. The aliquot (1 μl) skin odor extracts were analyzed in splitless mode (purge flow to split vent 3 mL/min at 0.8 min) with temperature programming of the column oven (35 ˚C for 5 min then increase at 10 ˚C/min to 280 ˚C and held at this temperature for 10.5 min) and helium as the carrier gas (1.2 ml/min flow rate, 8.8271 psi head pressure). The GC injector and MS transfer line were kept at 270 ˚C and 280 ˚C, respectively, while the MS ion source and quadrupole were respectively kept at 230 ˚C and 150 ˚C. Mass spectra were acquired with a solvent delay of 3 min, with mass ranging from 38—550 Daltons (Da) and a threshold of 70 Da at a scan time of 0.73 scans/sec. The volatile organic compounds were tentatively identified using their retention times, electron ionization mass spectra and Kovats retention indices (RIs) which were compared with library GC/MS data (NIST11, Adams2 and Chemecol), published mass spectra and RIs from online NIST library database. Additionally, the identities of the previously identified repellent ketones 6-methyl-5-hepten-2-one, acetophenone, and geranyl acetone (Olaide et al. 2019) were confirmed with commercially purchased standards. The RIs of the VOCs were obtained using commercial standards of straight chain alkanes (C_7_-C_30_, 49,451-U, Supelco, Bellefonte, Pennsylvania, USA) run on the GC/MS using the same conditions described earlier for the skin odor samples. Retention indices (non-isothermal) were calculated using the formula: \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$${I}_{x}=100(n+({t}_{x}-{t}_{n})/({t}_{n+1}-{t}_{n})$$\end{document} , according to Van den Dool and Kratz (1963) for temperature-progammed GC where \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$${I}_{x}$$\end{document} is the Kovats retention index of VOC \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$X$$\end{document} with retention time \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$${t}_{x}$$\end{document} , \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$${t}_{n}$$\end{document} and \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$${t}_{n+1}$$\end{document} respectively are the retention times of the reference n-alkanes eluting immediately before and after the VOC \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$X$$\end{document} , and \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$n$$\end{document} is the number of carbon atoms in the n-alkane eluting before VOC \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$X$$\end{document} . Using the absolute peak areas (abundance), the amount (μg/μl) of the repellent ketones were quantified in the skin odor samples of the vertebrates. To achieve this, external quantification was done using commercial samples of acetophenone for the benzenoid ketone and geranyl acetone for the aliphatic ketones prepared in five known concentrations (50 ng/µL to 250 ng/µL) within the expected range in the crude skin odors (Olaide et al. 2019). The peak areas of the commercial standards obtained after GC–MS analysis were used to generate calibration curves and linear equations (R^2^ = 0.986, y = 6E + 06x − 2E + 07 for acetophenone; and R^2^ = 0.988, y = 1E + 06x − 2E + 07 for geranyl acetone and 6-methyl-5-hepten-2-one) from which the naturally occurring-amounts in the skin odors were estimated. All peaks detected in the control (blank cotton material) were considered as contaminants and therefore discarded in the volatile analysis.
Chemicals
The commercial chemicals used in this study were sourced as follows: 6-methyl-5-hepten-2-one (99%, M48805, Sigma-Aldrich), acetophenone (99%, A10701, Sigma-Aldrich), geranyl acetone (96%, 328,677, Sigma-Aldrich), dichloromethane solvent (≥ 99.9%, analytical grade, 270,997, Sigma-Aldrich) and C_7_-C_30_ straight chain alkanes (49,451-U, Supelco).
Statistical Analyses
All data analyses of the skin-derived volatile organic compounds (VOCs) of zebra, donkey, horse, waterbuck, buffalo and warthog were performed using R statistical software version 4.4.1, in the R Studio graphical user interface (R Core Team 2024). The dataset (abundance of the VOCs across replicates of the different vertebrates) was first tested for normality using Shapiro–Wilk’s test, and homogeneity of variance using Bartlett’s test. Since the dataset was found to be not normally distributed and with non-homogenous variance, it was analyzed using the non-parametric tests Kruskal–Wallis test to compare abundance of the VOCs across the different vertebrates. When a significant difference was observed (P < 0.05), a Dunn’s post hoc test with Bonferroni adjustment was applied for means separation using the “dunn.test” package (Dinno 2024). The VOCs that were detected in only two groups (vertebrate species) were analyzed using the Mann–Whitney U test. All analyses were carried out at 5% probability level, i.e. alpha < 0.05. The machine learning algorithm random forest (RF) analysis (Breiman 2001) was employed to select the most discriminating VOCs among the vertebrates, based on the mean decrease in accuracy (MDA) obtained using the function “importance” in the “randomForest” package. The higher the MDA value, the higher its importance to discrimination (Liaw and Wiener 2002; Ranganathan and Borges 2010). Based on the abundance of top most discriminating VOCs, Sparse Partial Least Square Discriminant Analysis (sPLS-DA) was performed to visualize similarity of the vertebrate skin VOC profiles using the “mixOmics” and “ade4” packages (Dray and Dufour 2007; Hervé et al.2018; Lê Cao et al. 2011; Thioulouse et al 2018) as in previous studies (Adams et al. 2023; Ayelo et al. 2022). The function “tune.splsda” in the “mixOmics” package was used to select the optimum number of components and variables (volatiles) in the sPLS-DA model for enhanced classification accuracy. Further, biplot of the sPLS-DA was performed using the function “biplot” in the “mixOmics” package (Rohart et al. 2017), to illustrate variations in the VOC profiles of the different vertebrates and to highlight the correlation between the top discriminating VOCs and the different vertebrates. A clustering heatmap was carried out using the function “cim” in the “mixOmics” package (Rohart et al. 2017), to illustrate the variations in emission of the most discriminating VOCs across skin odors of replicate vertebrates. The out-of-bag error of the RF analysis was used to estimate the classification accuracy (1-OOB error) (Liaw and Wiener 2002; Ranganathan and Borges 2010). Similarly, the quality of the sPLS-DA model fit and prediction accuracy was validated using the function “perf”, the quality parameters R2X and R2Y (explaining the variation in the most discriminating VOCs and vertebrate groups, respectively), and the “leave-one-group-out” cross-validation method in the “mixOmics” package (Rohart et al. 2017). The sPLS-DA plot, biplot and clustering heatmap were also used to visualize and highlight the variations in the occurrence, abundance and natural ratios of previously identified repellent compounds across the different vertebrates.
Ethics Statement
The Kenyan Wildlife Service KWS approved the sampling of skin odors of buffalo, warthog, waterbuck, and zebra (permit number: KWS/BRM/5001). Internal ethical clearance for use of donkey and horse was sought from icipe’s institutional biosafety committee. Further, oral consent from the horse and donkey farmers was sought to collect skin odor samples from their animals for use in our study.
Results
Analysis of Volatile Organic Compounds In Vertebrate Skin Odors
A total of 96 volatile organic compounds (VOCs) were detected in the headspace skin emission profiles of buffalo, donkey, horse, warthog, waterbuck, and zebra (Fig. 2, Table 1). These compounds belong to 11 chemical classes namely: alkanes (20), alkenes (9), benzenoids (17), alcohols (10), phenols (2), furans (3), ketones (11), aldehydes (15), benzoquinone (1), esters (4), and a lactone (1), which vary among the vertebrate species. Alkanes ranged between (1.84–25.45%), alkenes (0.21–6.16%), benzenoids (1.17–9.67%), alcohols (6.57–29.94%), phenols (0–10.73%), furans (0–1.0%), ketones (4.81–59.14%), aldehydes (12.22–45.19%), benzoquinone (0–0.78%), esters (0.44–3.30%), and lactone (0–0.68%). The least and most abundant VOCs in the different classes were: alkanes (dodecane-15.38%, tetracosane-57.48%); alkenes ((E)-oct-2-ene-0.17%, dodec-1-ene-32.92%); benzenoids (toluene-0.25%, α-methylstyrene-23.89%); alcohols (hexanol-0.21%, α,α-dimethylbenzenemethanol-45.13%); phenols (p-cresol-44.44%, o-guaiacol-55.56%); furans (2,4-dimethylfuran-25.24%, 2-pentylfuran-47.92%); ketones (acetophenone- 45.14%, 2,3-octanedione-0.22%); aldehydes (nonanal- 39.48%, (E)-hex-2-enal-0.1%); and esters (hex-3-enyl acetate-4.03%, isopropyl hexadecanoate-59.48%).Fig. 2. Representative total ion chromatograms (TIC) of the skin volatile emission profiles of different vertebrates. (a) buffalo, (b) donkey, (c) horse, (d) warthog, (e) waterbuck, (g) and (h) zebra skin odors collected in Amboseli and Nguruman, respectively. Numbers correspond to the list presented in Table 1. The most discriminating VOCs distinguishing the vertebrates (“mean decrease in accuracy” in random forest analysis) are in bold. Chemical structures of the repellent ketones previously identified from zebra skin odor, that is 6-methyl-5-hepten-2-one (30), acetophenone (42) and geranyl acetone (72), are shown. Vertebrate illustrations are not to scale, obtained via BioRender.com and Flaticon.comTable 1Volatile Organic Compounds Detected In The Skin Emission Profiles Of Preferred And Non-Preferred Vertebrate Hosts Of Tsetse Flies (n = 5)Median Abundance (Peak Area) (× 10^6^) ± SEMdn^g^Peak no.^a^t_R_^b^ (min)I^c^ObsI^d^Lit_Compound^e^MF^b^Chemical class^f^BuffaloDonkeyHorseWarthogWaterbuckZebra_ANPZebra_NguP value^h^13.12-686^A^pentan-2-oneC_5_H_10_Oketonend1.53 ± 0.171.10 ± 0.55ndndndnd0.5323.86700702^A^pentanalC_5_H_10_Oaldehyde0.00 ± 0.34^a^0.28 ± 0.13^ab^ndnd0.22 ± 0.23^ab^0.20 ± 0.11^b^1.79 ± 0.80^a^0.02334.02706695^B^2,4-dimethylfuranC_6_H_8_Ofuran0.00 ± 0.15ndndndnd0.18 ± 0.11nd145.46760762^A^tolueneC_7_H_8_benzenoid0.00 ± 0.05^a^ndnd0.30 ± 0.07^b^0.16 ± 0.06^ab^0.14 ± 0.05^ab^nd0.01956.27790789.6^B^oct-1-eneC_8_H_16_alkenend0.21 ± 0.110.00 ± 0.100.00 ± 0.400.23 ± 0.17nd0.96 ± 0.650.36966.44797782^C^4-methyl-3-penten-2-oneC_6_H_10_Oketone26.06 ± 13.61ndndnd1.00 ± 1.6620.53 ± 12.13nd0.11776.56801800^D^hexanalC_6_H_12_Oaldehyde0.00 ± 0.24^b^0.98 ± 0.18^ab^0.58 ± 0.31^ab^0.75 ± 0.40^ab^4.83 ± 3.53^a^0.00 ± 1.95^ab^6.70 ± 5.61^a^0.01486.9815797.9^B^(E)-oct-2-eneC_8_H_16_alkenendndndnd0.16 ± 0.06ndnd97.66844841.3^B^4-hydroxy-4-methylpentan-2-oneC_6_H_12_O_2_ketone26.71 ± 12.89^a^ndnd1.37 ± 5.72^b^10.46 ± 10.50^ab^4.89 ± 3.05^b^nd0.021107.97856854^E^(E)-hex-2-enalC_6_H_10_Oaldehydendndndnd0.52 ± 0.460.00 ± 0.19nd0.332118.04859855^F^2-furanmethanolC_5_H_6_O_2_furanndndnd0.19 ± 0.08^ab^0.34 ± 0.08^a^0.00 ± 0.06^b^nd0.018128.13863868.3^B^ethylbenzeneC_8_H_10_benzenoidnd0.00 ± 0.090.00 ± 0.140.20 ± 0.120.18 ± 0.02nd1.48 ± 1.280.285138.35871877.8^B^1,3-dimethylbenzene^^C_8_H_10_benzenoid0.38 ± 0.25^ab^ndnd0.24 ± 0.37^ab^1.37 ± 0.70^a^0.12 ± 0.05^b^nd0.013148.41874871^G^hexanolC_6_H_14_Oalcoholndndnd0.26 ± 0.100.44 ± 0.15ndnd0.105158.74887884^F^4-cyclopentene-1,3-dione^^C_5_H_4_O_2_ketonendndnd0.31 ± 0.090.72 ± 0.230.43 ± 0.47nd0.151168.84891890^D^styreneC_8_H_8_benzenoid0.14 ± 0.10ndnd0.25 ± 0.080.36 ± 0.080.22 ± 0.17nd0.449178.92894893^E^non-1-eneC_9_H_18_alkenend1.06 ± 0.320.00 ± 0.26ndndnd0.00 ± 3.730.13189.1901900^E^nonaneC_9_H_2_0alkane1.11 ± 0.61^b^0.58 ± 0.24^ab^1.01 ± 0.17^ab^0.27 ± 0.25^ab^0.26 ± 0.13^a^0.50 ± 0.15^ab^0.00 ± 1.03^ab^0.048199.16904903^A^heptanal^^C_7_H_14_Oaldehyde4.21 ± 2.26^ab^3.16 ± 6.21^ab^1.68 ± 0.51^b^1.36 ± 0.38^b^5.01 ± 1.46^ab^3.13 ± 0.88^ab^94.03 ± 53.39^a^** < 0.001**209.23907909^H^2-butoxyethanolC_6_H_14_O_2_alcoholndndnd0.00 ± 0.920.94 ± 0.330.39 ± 0.28nd0.785219.5920888^I^1,4-benzoquinoneC_6_H_4_O_2_benzoquinone0.00 ± 0.12ndnd0.00 ± 0.700.51 ± 0.040.33 ± 0.61nd0.12229.85937933.3^B^2,6-dimethyloctaneC_10_H_22_alkanendnd0.76 ± 0.320.00 ± 0.15ndnd4.96 ± 1.420.0612310.34960956^D^(E)-hept-2-enalC_7_H_12_Oaldehyde0.79 ± 0.67ndndnd1.53 ± 0.300.34 ± 0.49nd0.5392410.4963962^A^benzaldehydeC_7_H_6_OAldehyde^#^3.05 ± 1.10^ab^0.80 ± 0.40^b^3.07 ± 0.52^a^1.36 ± 0.41^ab^1.48 ± 0.20^ab^1.71 ± 0.32^ab^36.26 ± 19.55^a^0.0162510.63974969^D^heptanolC_7_H_16_Oalcohol0.41 ± 0.27ndndnd0.36 ± 0.180.30 ± 0.19nd0.8042610.8982unidentified1^^-0.68 ± 0.57ndndnd0.71 ± 0.24ndnd12710.83983987.9^B^α-methyl styrene^^C_9_H_10_benzenoid0.80 ± 0.33^ab^0.00 ± 0.17^b^nd1.00 ± 0.19^ab^0.97 ± 0.31^b^1.17 ± 0.27^ab^65.19 ± 32.48^a^0.0022810.9987987^ J^2,3-octanedione^^C_8_H_14_O_2_ketonendndndnd1.23 ± 0.53ndnd2910.99991997^ K^2,2,4,6,6-pentamethylheptane^^C_12_H_26_alkane4.76 ± 4.84^b^ndnd14.05 ± 7.60^ab^nd2.44 ± 4.00^b^86.27 ± 12.55^a^0.0043010.99991987^E^6-methyl-5-hepten-2-one^^C_8_H_14_Oketone23.32 ± 6.84^a^0.44 ± 0.35^b^2.89 ± 0.72^ab^nd3.96 ± 2.65^ab^5.74 ± 4.07^ab^24.68 ± 13.39^a^0.0033111.04993993^E^2-pentylfuranC_9_H_14_Ofuranndndndnd0.91 ± 0.330.43 ± 0.20nd0.2093211.079951000^B^1,2,4-trimethylbenzene^^C_9_H_12_benzenoid1.07 ± 0.280.91 ± 1.530.00 ± 0.66ndndnd4.17 ± 8.650.0973311.08995974.2^B^mesityleneC_9_H_12_benzenoidnd0.47 ± 0.663.54 ± 1.390.58 ± 0.63ndnd10.71 ± 8.580.0823411.1910011000^L^decane^^C_10_H_22_alkanendndnd0.68 ± 0.600.62 ± 0.190.00 ± 0.25nd0.0923511.2610041004^G^octanal^^C_8_H_16_Oaldehyde11.30 ± 3.36^ab^8.38 ± 1.09^ab^11.64 ± 2.37^ab^4.28 ± 1.06^b^1.63 ± 0.34^b^3.71 ± 1.57^b^177.80 ± 69.14^a^** < 0.001**3611.3510091004^ M^hex-3-enyl acetateC_8_H_14_O_2_ester0.56 ± 0.42ndndndnd0.53 ± 0.27nd13711.7410311030^N^2-ethylhexan-1-ol^^C_8_H_18_Oalcohol2.39 ± 1.13nd0.38 ± 1.40nd0.90 ± 0.401.46 ± 0.3452.12 ± 33.990.1253811.910401033^D^benzyl alcoholC_7_H_8_Oalcohol^#^6.77 ± 7.911.43 ± 0.56nd1.05 ± 0.460.85 ± 0.262.41 ± 0.830.00 ± 3.970.1573911.951043unidentified2-ndnd10.90 ± 3.04ndndndnd4012.1410541058.2^B^1-methyl-3-propylbenzeneC_10_H_14_benzenoidndndndndndnd7.67 ± 10.604112.2810611058.2^B^(E)-oct-2-enal^^C_8_H_14_Oaldehyde3.75 ± 0.703.52 ± 0.592.42 ± 1.58nd1.17 ± 0.221.39 ± 0.510.00 ± 11.360.1784212.4410701067^E^acetophenone^^C_8_H_8_Oketone^#^109.93 ± 32.69^b^2.79 ± 0.63^a^nd46.97 ± 34.56^ab^22.66 ± 16.85^ab^26.32 ± 23.85^ab^47.44 ± 12.79^ab^0.0224312.510741070^D^octan-1-olC_8_H_18_Oalcoholnd2.29 ± 0.82^a^93.86 ± 20.11^b^ndndnd0.00 ± 10.40^a^0.0164412.5510771086^A^p-cresolC_7_H_8_Ophenol1.55 ± 14.37ndndndndndnd4512.7610881089.6^B^α,α-dimethylbenzenemethanol^^C_9_H_12_Oalcohol^#^5.03 ± 1.47^b^13.96 ± 3.60^ab^69.54 ± 13.12^ab^6.15 ± 3.31^b^3.40 ± 3.61^b^2.45 ± 4.56^b^120.04 ± 7.78^a^** < 0.001**4612.8310921090^G^o-guaiacolC_7_H_8_O_2_phenol7.99 ± 1.83ndnd5.43 ± 7.204.56 ± 3.232.21 ± 1.72nd0.5084712.9811011100^B^undecane^^C_11_H_24_alkane2.07 ± 1.04^b^7.86 ± 2.35^ab^0.00 ± 1.37^b^2.04 ± 0.91^b^ndnd107.42 ± 12.28^a^0.0054813.0111031100^B^methylbenzoateC_8_H_8_O_2_ester^#^ndnd4.84 ± 4.07ndndndnd4913.0711061103^D^nonanal^^C_9_H_18_Oaldehyde13.49 ± 2.73^bc^53.88 ± 3.^26ab^44.76 ± 2.72^ab^10.64 ± 2.09^bc^3.58 ± 0.77^c^17.34 ± 5.35^abc^447.65 ± 35.38^a^** < 0.001**5013.2311161259^O^4-methylundecaneC_12_H_26_alkane1.87 ± 0.69ndndndnd0.94 ± 1.18nd0.835113.4211281031^A^p-cymeneC_10_H_14_benzenoid^^nd0.00 ± 0.860.00 ± 0.441.52 ± 0.81ndnd11.43 ± 6.960.3235213.4211281026^P^o-cymeneC_10_H_14_benzenoid^^nd1.50 ± 0.51ndndnd0.90 ± 1.0524.27 ± 12.860.1245313.9711631163^G^(E)-non-2-enal^^C_9_H_16_Oaldehydend6.27 ± 1.32^abc^9.32 ± 1.02^ab^0.84 ± 0.35^bc^1.14 ± 0.27^bc^0.00 ± 0.44^c^97.18 ± 30.32^a^** < 0.0015414.0611691167^Q^o-hydroxyacetophenoneC_8_H_8_O_2_ketone3.78 ± 0.41ndnd2.79 ± 2.611.42 ± 0.701.32 ± 0.82nd0.1475514.1111721193^E^dodec-1-eneC_12_H_24_alkenend0.00 ± 0.58^b^0.00 ± 0.90^b^ndndnd23.22 ± 10.78^a^0.0325614.411901181^D^p-methylacetophenoneC_9_H_10_Oketone^#^2.56 ± 0.73ndnd2.84 ± 2.041.68 ± 1.211.02 ± 0.79nd0.3955714.5511991200^E^dodecaneC_12_H_26_alkane1.44 ± 0.51^ab^5.02 ± 1.67^ab^8.12 ± 2.14^ab^0.53 ± 0.54^b^0.43 ± 0.17^b^0.81 ± 0.17^b^115.38 ± 14.91^a^0.0035814.6612071207^E^decanal^^C_10_H_20_Oaldehyde3.61 ± 0.44^bc^19.21 ± 6.69^abc^28.23 ± 2.85^ab^6.22 ± 0.74^abc^2.14 ± 0.52^c^2.46 ± 1.22^bc^108.67 ± 14.43^a^ < 0.0015914.7812151213.6^L^2,6-dimethylundecaneC_13_H_28_alkanendndndndndnd7.89 ± 8.076015.041233unidentified3^^-ndnd8.03 ± 1.76ndndnd0.00 ± 3.540.1316115.5112661264^E^(E)-dec-2-enalC_10_H_18_Oaldehydendnd5.38 ± 7.36ndnd0.49 ± 0.240.00 ± 4.790.1436215.912921293^E^tridec-1-eneC_13_H_26_alkene0.55 ± 0.482.47 ± 0.810.00 ± 1.05ndndnd0.00 ± 2.430.2846316.0213011300^G^tridecane^^C_13_H_28_alkane0.97 ± 0.34^abc^6.66 ± 0.71^ab^6.51 ± 1.39^ab^0.54 ± 0.62^bc^0.25 ± 0.05^c^0.74 ± 0.12^bc^87.51 ± 18.66^a^ < 0.0016416.1613111308.8^B^undecanalC_11_H_22_Oaldehyde0.00 ± 0.47^b^3.35 ± 0.99^ab^12.36 ± 5.73^a^0.00 ± 0.21^b^0.52 ± 0.09^ab^0.62 ± 0.22^ab^29.42 ± 11.82^ab^0.0246516.9613701260^D^γ-octalactone^^C_8_H_14_O_2_lactonendndndnd0.84 ± 0.19ndnd6617.2313901393^E^tetradec-1-eneC_14_H_28_alkene0.24 ± 0.092.12 ± 0.83ndndndndnd0.3896717.3614001400^N^tetradecane^^C_14_H_30_alkane1.01 ± 0.71^abc^9.69 ± 0.74^ab^6.52 ± 1.87^ab^0.58 ± 0.23^bc^0.31 ± 0.05^c^0.46 ± 0.28^bc^50.26 ± 19.54^a^ < 0.0016817.5414141412^G^dodecanalC_12_H_24_Oaldehydend2.72 ± 0.810.00 ± 10.66ndnd0.00 ± 0.070.00 ± 8.260.5926917.5914181511^D^tridecanalC_13_H_26_Oaldehydend0.00 ± 1.083.38 ± 1.18ndndndnd0.3737017.7414301419^E^α-cedreneC_15_H_24_alkene^^1.10 ± 0.32^ab^4.14 ± 0.98^a^3.08 ± 1.33^a^0.37 ± 0.87^ab^0.17 ± 0.07^b^0.23 ± 0.19^ab^15.82 ± 6.62^a^**0.003**7117.8814411424^G^β-cedreneC_15_H_24_alkene^^0.00 ± 0.070.00 ± 0.62ndndndndnd0.7247218.1214591453^D^geranyl acetone^^C_13_H_22_Oketone^^0.00 ± 0.05^c^4.60 ± 0.49^ab^5.55 ± 0.66^ab^0.00 ± 0.31^bc^0.33 ± 0.07^abc^nd17.74 ± 8.08^a^** < 0.001**7318.4214831575^R^tridecan-1-olC_13_H_28_Oalcoholndnd6.23 ± 6.75ndndnd0.00 ± 1.710.1197418.6114981772^S^pentadecan-1-ol^*^C_15_H_32_Oalcoholnd3.65 ± 0.30^a^0.00 ± 0.73^b^ndndndnd**0.009**7518.6515011500^L^pentadecane^*^C_15_H_32_alkane0.33 ± 0.14^c^7.42 ± 0.52^b^7.03 ± 2.65^b^nd0.21 ± 0.04^c^0.28 ± 0.12^c^42.93 ± 17.70^a^**0.018**7619.1715441526^ T^1-butylhexylbenzene^*^C_16_H_26_benzenoid0.22 ± 0.40^c^2.03 ± 0.71^b^7.10 ± 3.04^a^ndnd0.23 ± 0.21^c^7.11 ± 5.92^a^**0.025**7719.2915541534^ T^1-propylheptylbenzeneC_16_H_26_benzenoidnd0.00 ± 0.752.23 ± 1.54ndnd0.00 ± 0.040.00 ± 0.730.3757819.5415751553^ T^1-ethyloctylbenzeneC_16_H_26_benzenoidndnd2.60 ± 1.22ndndnd0.00 ± 1.520.4517919.8416001587^U^hexadec-1-eneC_16_H_32_alkenend0.00 ± 0.670.00 ± 0.500.00 ± 0.23ndnd0.00 ± 0.740.9638019.8516011600^E^hexadecaneC_16_H_34_alkane0.00 ± 0.40^b^4.58 ± 0.45^ab^2.65 ± 1.30^ab^ndnd0.00 ± 0.19^b^37.75 ± 19.09^a^**0.005**8120.1416271607.9^L^cedrol^*^C_15_H_26_OAlcohol^^0.97 ± 0.363.88 ± 0.640.00 ± 1.130.00 ± 0.47ndnd13.06 ± 6.160.0728220.216321626^ T^1-butylheptylbenzeneC_17_H_28_benzenoidndnd0.00 ± 0.35ndnd0.00 ± 0.055.15 ± 5.240.1658320.3416441620^ T^1-pentylhexylbenzeneC_17_H_28_benzenoidnd0.00 ± 0.401.84 ± 0.82ndndndnd0.0768420.3716471625^G^benzophenone^^C_13_H_10_Oketone0.61 ± 0.62^b^13.42 ± 2.01^ab^58.19 ± 16.13^a^1.03 ± 0.55^b^1.06 ± 0.57^b^0.37 ± 1.09^b^2.78 ± 7.97^ab^0.0038521.0117041700^L^heptadecaneC_17_H_36_alkane0.00 ± 0.053.07 ± 0.942.23 ± 1.250.00 ± 0.19ndnd3.80 ± 3.270.1948621.3817381719^ T^1-pentylheptylbenzeneC_18_H_30_benzenoidnd2.37 ± 0.242.15 ± 1.03ndnd0.00 ± 0.063.36 ± 4.670.0508721.4417431723^ T^1-butyloctylbenzeneC_18_H_30_benzenoidnd0.00 ± 0.602.30 ± 1.49ndnd0.00 ± 0.050.00 ± 2.260.1948822.0818021800^E^octadecaneC_18_H_38_alkanend2.05 ± 0.270.77 ± 0.880.00 ± 0.13ndnd0.00 ± 3.830.0598922.418331831^ V^isopropyl tetradecanoate^^C_17_H_34_O_2_esternd3.85 ± 0.56^a^0.00 ± 0.40^b^ndndndnd0.01**9023.1519061900^E^nonadecaneC_19_H_40_alkanend3.02 ± 0.342.48 ± 2.23ndndnd0.00 ± 11.350.1279124.0820012000^E^eicosaneC_20_H_42_alkanend3.01 ± 0.372.18 ± 0.86ndndnd0.00 ± 1.790.4559224.3720321999^D^isopropyl hexadecanoateC_19_H_38_O_2_ester0.16 ± 0.154.31 ± 0.582.78 ± 1.271.12 ± 0.39nd0.38 ± 0.388.83 ± 5.170.0719325.0821082100^Q^heneicosaneC_21_H_44_alkanend3.85 ± 1.084.20 ± 1.32ndndnd0.00 ± 2.210.3639425.9722072200^Q^docosaneC_22_H_46_alkanend3.19 ± 0.302.74 ± 1.82ndndnd0.00 ± 2.600.3059526.8423072300^Q^tricosaneC_23_H_48_alkanend2.23 ± 0.244.06 ± 1.79ndndndnd0.1439627.6724082400^Q^tetracosaneC_24_H_50_alkanend1.47 ± 0.690.00 ± 1.16ndndndnd0.656^a^ Peak number correlates with numbering on the total ion chromatograms in Fig. 2^b^ t_R (min) = retention time in minutes; MF = molecular formular^c^ IObs = retention index observed in this study calculated based on C_7_-C_30_ n-alkanes of HP-5MS capillary column (30 m/0.25 mm/0.25 μm, He, 35 ˚C @ 5 min, 10 ˚C/min, 280 ˚C @ 10.5 min; T_end_: 280 ˚C)^d^ ILit = retention index obtained from literature. References: ^A^ Bonaiti et al. 2005; ^B^ Xu et al. 2003; ^C^ Yamaguchi and Sibamoto-1981; ^D^ Pino et al. 2005; ^E^ Flamini et al. 2006; ^F^ Deport et al. 2006; ^G^ Asuming et al. 2005; ^H^ Cajka et al. 2007; ^I^ Zenkevich 2005; ^J^ Turchini et al. 2004; ^K^ Insausti et al. 2005; ^L^ Zeng et al. 2007; ^M^ Campeol et al. 2003; ^N^ Sartin et al. 2001; ^O^ Kotowska et al. 2012; ^P^ Vagionas et al. 2007; ^Q^ Radulovic et al. 2009; ^R^ Skaltsa et al. 2001; ^S^ Saroglou et al. 2007; ^T^ Peng et al. 1992; ^U^ Mimica-Dukic et al. 2003; ^V^Lalel et al. 2003^e^ Compounds were identified based on their retention times, electron ionization mass spectra and retention indices compared with library GC/MS data (NIST11, Adams2 and Chemecol) and published mass spectra and retention indices s from online NIST library database. ^*^ indicates most discriminating VOCs distinguishing the vertebrates based on the “mean decrease in accuracy” in random forest analysis; compounds in bold are the primary focus repellent ketones in this study previously identified from zebra skin odor and for which identities were confirmed using authentic standards^f^ Chemical class: compounds are also aromatic (^#^) or terpenoids (^$^)^g^ Median Abundance (× 10^6^) ± standard error of the median (SEMdn): volatiles were collected from five biological replicates for each vertebrate species. Zebra_ANP and Zebra_Ngu = Zebra skin odor collected in Amboseli NP and Nguruman, respectively. Means with different superscript letters are significantly different (Kruskal-Wallis test, or Mann-Whitney U test where applicable, followed by Dunn’s post hoc test; P < 0.05). nd = not detected^h^ P – value = probability value of the non-parametric Kruskal–Wallis and Mann–Whitney U tests comparing abundance of the 96 VOCs across the different vertebrates. Significant values (i.e. P < 0.05) are in bold
Proposed Repellent Biomarkers for Less Preferred Vertebrate Hosts
The proposed three ketone repellent biomarkers 6-methyl-5-hepten-2-one, acetophenone and geranyl acetone previously identified from zebra skin odor for tsetse flies were generally present in all the vertebrates tested but were emitted at different levels (Kruskal–Wallis followed by Dunn’s test; P < 0.05; Table 1). Remarkably, all three ketones were detected in the non-preferred hosts waterbuck and zebra which was generally not the case for the preferred hosts buffalo, donkey, horse, and warthog. However, exceptions were recorded in the non-preferred zebra skin odor from Amboseli, where geranyl acetone was not detected. Further quantification of these compounds revealed their natural ratios in skin volatile emissions of the different vertebrate hosts (Table 2). Notably, where present, the ratios of acetophenone relative to the other two ketones were different for the non-preferred waterbuck and zebra compared to buffalo, donkey, horse, and warthog which are preferred hosts of tsetse flies. Table 2. Ratios Of The Zebra-Derived Tsetse Repellent Ketones Across Different Vertebrate Skin Odors (n = 5)Compoundbuffalodonkeyhorsewaterbuckwarthogzebra_ANP^a^zebra_Ngu^a^6-methyl-5-hepten-6-one1111nd11acetophenone41nd2322geranyl acetone1^b^1111nd1^a^ zebra_ANP and zebra_Ngu are skin odors collected from zebra in Amboseli NP and Nguruman, respectivelynd = not detected^b^ detected in the skin emissions of only one individual buffalo
Based on the occurrence and abundance of the three ketones in the skin volatile emission profiles, the vertebrates were grouped together, except for horse (Fig. 3a) and zebra (Nguruman) (Fig. 3b), respectively. By contrast, using the ratios of these ketones the vertebrates were grouped into four: one composed of buffalo; a second group had only warthog; and a third group comprised zebra (Nguruman), waterbuck, donkey and horse (Fig. 3c). The latter group clustered closely together; and a fourth group had only zebra (Amboseli) which grouped separately from the others (Fig. 3c). The first two dimensions of the sPLS-DA accounted for 79% of the total variation with dimension 1 explaining 46% and dimension 2 accounting for 33% of the total variation. In addition, the clustering heatmap (horizontal direction) classified the vertebrates into two main groups, that is, warthog and buffalo, and others (Fig. 3c).Fig. 3. Variations in zebra skin-derived tsetse repellent ketones in non-preferred and preferred vertebrate hosts. Sparse partial least square discriminant analysis (sPLS-DA) grouping of the vertebrates (left), a sPLS-DA biplot classification of vertebrates and correlation of volatiles (center), and Heatmap clustering showing the abundance (in decreasing color intensity) across vertebrate skin odors (right) using (a) occurrence, (b) abundance and (c) natural ratios of occurrence of the repellent ketones 6-methyl-5-hepten-2-one, acetophenone and geranyl acetone found in the skin volatile emission profiles of different vertebrates. Zeb_ANP and Zebra_Ngu = zebra skin odor collected from Amboseli and Nguruman, respectively. Skin odors were collected from five (5) individuals for each vertebrate species
Contribution of Background Skin Odors to Tsetse Fly Discrimination of Non-Preferred Vertebrates
Thirty (30) VOCs were found to be the best to distinguish between the different vertebrates based on the “mean decrease in accuracy” (Random Forest analysis, MDA) (Figs. 3a, 4). The abundance of which varied significantly across the vertebrates (Kruskal–Wallis test followed by Dunn’s post hoc test, or Mann–Whitney U test where applicable; P < 0.05) (Table 1). Using the top most discriminating compounds (MDA > 7), the vertebrates were grouped into four clusters comprising of: (i) zebra (Nguruman), (ii) waterbuck, (iii) buffalo, warthog, zebra (Amboseli), and horse, and (iv) donkey (Fig. 4b). While groups (i) and (ii) clustered completely separately, groups (iii) to (iv) clustered closely together. The quality parameters of the random forest (that is, 1—OOB error (out-of-bag error) of 97.14%) and sPLS-DA (i.e., R2X 0.81 and R2Y 0.82) models show their strong predictive power and accurate classification of the different vertebrates based on their specific skin-derived VOCs.Fig. 4. Discriminating volatile organic compounds (VOCs) and their correlation with different vertebrates. (a) Thirty (30) most discriminating VOCs in skin volatile emission profiles of buffalo, donkey, horse, warthog, waterbuck and zebra listed in decreasing importance based on the mean decrease in accuracy in the random forest analysis; (b) A sparse partial least square discriminant analysis (sPLS-DA) plot showing the distribution of VOCs across the vertebrate skin emissions using the top most discriminating volatiles (MDA > 7) (R2X = 0.81, R2Y = 0.82); (c) A sPLS-DA biplot showing the correlation of the top most discriminating VOCs with different vertebrates (R2X = 0.81, R2Y = 0.82). Numbers in bold within the plot are the discriminating VOCs corresponding to the numbering presented in Table 1 and total ion chromatograms in Fig. 2; (d) Heatmap clustering showing the abundance (in decreasing color intensity) of the most discriminant VOCs across replicate skin odors of the vertebrates. zebra_ANP = zebra skin odor collected from Amboseli; zebra_Ngu = zebra skin odor collected from Nguruman
We found a correlation between the top discriminating VOCs and the vertebrates, most of which were associated with zebra (Nguruman) and waterbuck (Fig. 4c). The first two (2) dimensions of the sPLS-DA explained 63% of the total variation, with dimensions 1 and 2 contributing 48% and 15%, respectively. Dimension 1 was associated mainly with 1-butylhexylbenzene, 1,2,4-trimethylbenzene, geranyl acetone, tetradecane, (Z)-non-2-enal, α,α-dimethylbenzenemethanol, tridecane, undecane, decanal and nonanal correlated with zebra (Nguruman) (Fig. 4c). However, dimension 2 was primarily associated with undecane, geranyl acetone, 1-butylhexylbenzene and 6-methyl-5-hepten-2-one correlated with zebra (Nguruman), acetophenone correlated with buffalo, isopropyl tetradecanoate correlated with donkey, and 1,3-dimethylbenzene, 2,3-octanedione, γ-octalactone correlated with waterbuck (Fig. 4c). Clustering heatmap classified the vertebrates into two main categories; one group composed of zebra (Nguruman), and a second group made up of the remaining vertebrates which were subsequently grouped separately (Fig. 4d).
Discussion
We found that the three repellent ketones 6-methyl-5-hepten-2-one, acetophenone and geranyl acetone previously identified in zebra skin odor were generally present in all the vertebrates tested, including buffalo, donkey, horse, warthog, waterbuck, and zebra. However, their ratios within the vertebrates varied specifically with acetophenone which appeared to distinguish non-preferred (zebra and waterbuck) and preferred (donkey, horse, buffalo, warthog) hosts. Additionally, our results suggest that background odors in each vertebrate’s skin volatile emission profiles could contribute to their overall discrimination by tsetse flies. Surprisingly, zebra skin odors collected in Nguruman and Amboseli National Park differed in their volatile emission profiles which could be associated with the sites. These results suggest that the three repellent ketones present in specific ratios combined with background vertebrate skin volatiles may serve as biomarkers for tsetse flies to distinguish between non-preferred- and preferred vertebrate hosts in their natural habitats.
In the current study, we found that while 6-methyl-5-hepten-2-one and acetophenone were common in zebra skin odors regardless of geographic location, geranyl acetone was detected only in Nguruman and not Amboseli suggesting it could be associated with genetic, physiological, pathological, and environmental factors (De Moraes et al. 2018; Emami et al. 2017; Stanczyk et al. 2018), which would require additional research.
Our analysis identified acetophenone as a potential discriminatory biomarker for tsetse flies to distinguish non-preferred and preferred hosts. Apart from occurrence (i.e. presence or absence) and concentration (i.e. abundance) in the skin volatile emissions, ratios in which individual compounds are found in skin odors could determine behavioral activity on arthropod vectors (Beyaert et al. 2010; Cha et al. 2011; Guerenstein and Lazzari 2009). Similarly, interactions of these compounds could dictate their effectiveness, as previously observed for phenols in cattle urine (Bursell et al. 1988; Vale et al. 1988). In turn, the ratios of behaviorally active repellents and their interactions could dictate the repellency of vertebrates which is consistent with our findings. Although this would require additional behavioral experiments to establish preferences of tsetse flies towards blends simulating the natural ratio of occurrence in the non-preferred and preferred vertebrates observed in this study. The role individual chemicals play in the repellency and their interaction with tsetse olfactory architecture vary (Diallo et al. 2020; Orubuloye et al. 2024). Previous research highlighted the importance of geranyl acetone as a superior spatial repellent compared to acetophenone (Olaide et al. 2021), and with strong antifeedant activity correlating with its effect on mRNA transcripts of the antennal olfactory receptors (ORs) of tsetse flies (Diallo et al. 2020). The higher vapor pressure of acetophenone than geranyl acetone could explain differences in their spatial repellency (Olaide et al. 2021). However, the antifeedant activity of acetophenone and their effects on mRNA transcripts of tsetse fly antennal ORs relative to geranyl acetone is currently unknown, and investigating this may unravel its importance in tsetse fly ecology.
Machine learning algorithms, such as random forest and sPLS-DA are increasingly applied in chemical ecology to identify discriminating VOCs involved in insect communication (Adams et al. 2023; Ayelo et al. 2022; Baleba et al. 2019; Hervé et al. 2018; Miano et al. 2022; Ranganathan and Borges 2010). This enhances the ability to handle complex data sets like the skin volatile emission profiles of vertebrates, identify patterns, and predict ecologically relevant components in the complex skin odor matrix. Interestingly, 6-methyl-5-hepten-2-one, acetophenone and geranyl acetone were among the most discriminating compounds of the 96 skin-derived VOCs detected in our study. Confirmation of the behavioral effects of other discriminatory compounds are required including assessment of physiological activity via electrophysiology.
We highlight three key limitations in our study. First, the sample size of individual vertebrate species used for odor collection was small (range) attributed to constraints in trapping wildlife and the use of vertebrates for research (Paul et al. 2016; Slijkerman et al. 2015; Verderio et al. 2023). Secondly, skin volatile emissions of vertebrates can vary by age and sex even for specific species as demonstrated in cattle (Torr et al. 2006, 2007), possibly linked to differing physiological states of the vertebrates and ecological roles of VOCs (Alberts 1992; Apps et al. 2015). Lastly, other potential confounders like host infection status, can contribute to the observed variation in the volatiles of the hosts examined (De Moraes et al. 2018; Emami et al. 2017; Stanczyk et al. 2018). However, this study provides valuable insights into the skin volatile emission profiles of vertebrate hosts and potential adaptive significance relevant to tsetse fly ecology.
We conclude that in nature, tsetse fly vertebrate hosts skin-derived volatiles vary based on the ecological settings, and the role of the three repellent ketones in discriminating non-preferred and preferred vertebrate hosts would depend on the skin background odor. As such geography, seasonality and a larger pool of vertebrates and sample size should be considered in investigating potential biomarkers that the vector may use to discriminate hosts for feeding. Additionally, the role of other sensory cues derived from physical sources including visual, thermal, tactile and acoustic combined with olfactory biomarkers should be investigated in future behavioral research for the integrated management of tsetse flies and African trypanosomosis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Channumsin M, Ciosi M, Masiga D, Auty H, Turner CM, Kilbride E, Mable BK (2021) Blood meal analysis of tsetse flies (Glossina pallidipes: Glossinidae) reveals higher host fidelity on wild com-pared with domestic hosts. Wellcome Open Res 6(213). 10.12688/wellcomeopenres.16978.110.12688/wellcomeopenres.16978.1PMC 851312334703903 · doi ↗ · pubmed ↗
- 2Dinno A (2024) Dunn.test: dunn's test of multiple comparisons using rank sums. R package version 1.3.6. https://CRAN.R-project.org/package=dunn.test. Accessed 15 Sept 2024
- 3FAO (2024) Programme Against African Trypanosomosis (PAAT), The Disease. Food and Agriculture Organization of the United Nations (FAO). Available: http://www.fao.org/paat/the-pro-gramme/the-disease/en/. Accessed 15 Sept 2024
- 4KWS (2024) Kenya Wildlife Service 2024, Amboseli National Park. https://www.kws.go.ke/amboseli-national-park. Accessed 15 Sept 2024
- 5Orubuloye OY, Mbewe NJ, Tchouassi DP, Yusuf AA, Pirk CW, Torto B (2024) An Overview of Tsetse Fly Repellents: Identification and Applications. J Chem Ecol 10.1007/s 10886-024-01527-510.1007/s 10886-024-01527-5PMC 1154371038976099 · doi ↗ · pubmed ↗
- 6R Core Team (2024) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org/. Accessed 15 Sept 2024
- 7Radulovic NS, Blagojevic PD, Palic RM, Zlatkovic BK, Stevanovic BM (2009) Volatiles from vegetative organs of the paleoendemic resurrection plants Ramonda serbica Panc. and Ramonda nathaliae Panc. at Petrov. J Serb Chem Soc 74(1):35–44. 10.2298/JSC 0901035 R
- 8Shaw APM, Cecchi G, Wint GRW, Cecchi G, Torr S, Waiswa C, Temesgen A, Eregae M, Abdi A, Muchina S, Mugasi S, Mattioli R, Mattioli RC, Robinson TP (2017) Intervening against bovine trypanosomosis in Eastern Africa: mapping the costs and benefits: mapping the costs and benefits. Programme Against African Try-panosomosis, Food and Agriculture Organistaion of the United Nations, Rome, Italy. Available: http://www.fao.org/3/a-i 7342 e. pdf. Accessed 15 Sept 2024