Pogo transposons provide tools to restrict cancer growth
Elina Zueva, Marianne Burbage

TL;DR
A gene from a transposon, POGK, acts as a tumor suppressor in aggressive breast cancer by controlling ribosome production and cell growth.
Contribution
The study reveals POGK's tumor-suppressive role in triple-negative breast cancer through its interaction with TRIM28 and regulation of ribosomal genes.
Findings
POGK, derived from a transposon, functions as a tumor suppressor in triple-negative breast cancer.
POGK interacts with TRIM28 to repress ribosomal genes and restrict cell growth.
An isoform switch in POGK leads to loss of its tumor-suppressive activity.
Abstract
Transposable elements provide material for novel gene formation. In particular, DNA transposons have contributed several essential genes involved in various physiological or pathological conditions. Here, we discuss recent findings by Tu et al. in Molecular Cell that identify Pogo transposon‐derived gene POGK as tumor suppressor in triple‐negative breast cancer (TNBC) by regulating ribosome biogenesis and restricting cell growth. An isoform‐switch in TNBC results in the loss of POGK capacity to recruit the epigenetic corepressor TRIM28 and to exert its repressive functions. These findings shed light on the potential for TE‐derived genes in providing new therapeutic opportunities for highly malignant TNBC. Here, we discuss findings by Tu et al. showing that POGK, a gene derived from the pogo DNA transposon, has tumor‐suppressor functions in triple‐negative breast cancer. POGK interacts…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
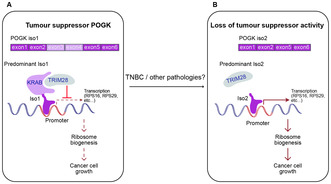 Figure 1
Figure 1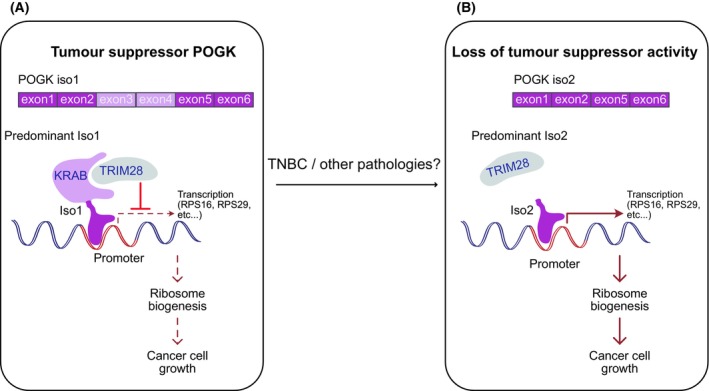 Figure 2
Figure 2- —Fondation ARC pour la Recherche sur le Cancer 10.13039/501100004097
- —Ligue Contre le Cancer 10.13039/501100004099
- —Agence Nationale de la Recherche 10.13039/501100001665
- —Paris Île‐de‐France Region—DIM ITAC
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCRISPR and Genetic Engineering · Chromosomal and Genetic Variations · RNA modifications and cancer
Transposable elements (TEs) constitute a significant portion of eukaryotic genomes. They are classified into two main categories. Class I retrotransposons move via an RNA intermediate using a copy‐and‐paste mechanism, while Class II DNA transposons use DNA intermediates through “cut‐and‐paste” or a “peel‐and‐paste” mechanisms [1]. Throughout evolutionary history, the parasitic propagation of TEs has profoundly reshaped genomic architecture. Coevolving with host genomes, TEs have contributed a wide array of regulatory elements such as promoters, enhancers, organizers of 3D genomic architecture, and drivers of novel splice isoforms, which occasionally encode proteins with altered cellular functions [2, 3, 4].
Recent technological advances have revealed the remarkable ability of TEs to contribute to the emergence of novel functional genes, with estimates ranging from dozens to thousands [5]. The most striking examples include the ARC gene encoding a repurposed Gag protein derived from a Ty3/Gypsy retrotransposon and playing a critical role in synaptic plasticity [6], and the Syncytin, a retroviral envelope protein that is essential for placental morphogenesis [7]. Interestingly, DNA transposons contribute a disproportionately larger number of TE‐derived genes than retrotransposons. These elements encode transposase enzymes that recognize specific nucleotide sequences within the cognate transposons, enabling their mobilization. Both the DNA‐binding and catalytic domains of transposases can be captured for cellular functions. Transposase capture likely contributed to the development of the adaptive immune system in jawed vertebrates. The V(D)J recombination process, which generates virtually limitless diversity in antibodies and T‐cell receptors, relies on RAG1/2 recombinases. They catalyze DNA cutting and joining through chemistry closely related to DNA transposases, with the RAG1 sequence showing evolutionary links to enzymes of Transib DNA transposons [8], suggesting that Transib‐like elements gave rise to this groundbreaking evolutionary innovation. In this example, the transposase retains its original function; however, a more common evolutionary outcome is the creation of proteins with entirely novel functions. For instance, the transposase domains of pogo transposons, part of the Tc1/mariner superfamily that has invaded nearly all animal taxa, have been repeatedly domesticated for novel functions. In mammals, they gave rise to centromere‐associated protein CENPB, which binds α‐satellite repeats in a sequence‐specific way to facilitate kinetochore formation [9]. The fusion of transposase domains with host ZNF or Krüppel‐associated box (KRAB) domains led to the emergence of the POGZ and POGK proteins, respectively [5].
Change in expression of pogo‐derived genes is associated with various pathologies. For example, disruption of POGZ expression is implicated in autism and intellectual disability [10, 11]. CENPB is aberrantly expressed in several cancers, including lung and colorectal cancers [12, 13]. In a recent issue of Molecular Cell, Tu et al. describe a tumor‐suppressor function of POGK in triple‐negative breast cancer (TNBC) [14], the most aggressive subtype with very few known therapeutic targets. They analyzed a group of transposable‐element‐derived genes (TEGs) across diverse cancer cohorts from the Cancer Genome Atlas (TCGA), identifying POGK as the top‐altered TEG transcript, most significantly dysregulated in breast cancers. Immunohistochemical analysis revealed reduced POGK expression in breast cancer tissues compared to normal samples, with a marked decrease in TNBC relative to non‐TNBC subtypes. Functional assays demonstrated that POGK overexpression impaired TNBC cell growth in vitro and in both immune‐competent and immune‐compromised mice. This growth suppression was accompanied by a significant reduction in ribosomal gene transcription and ribosome biogenesis. The authors showed that the KRAB domain of POGK interacts with the global transcriptional co‐repressor TRIM28 [15] (Fig. 1A). This complex specifically binds to DNA to repress gene expression, particularly the ribosomal genes RPS16 and RPS29. Notably, the ability of POGK to suppress cell growth relies on this KRAB–TRIM28 interaction, as a KRAB domain mutant was unable to inhibit cell growth. Mechanistically, the study revealed that POGK downregulation in TNBC likely results from isoform switching. In TNBC cells, the full‐length POGK isoform (iso1, Fig. 1A) is replaced by a shorter isoform (iso2, Fig. 1B) lacking the KRAB domain. Although iso2 retains the same DNA‐binding motif as iso1, it fails to recruit TRIM28, rendering it incapable of repressing RPS16 and RPS29 transcription. Furthermore, DOX‐inducible expression of iso2, unlike iso1, did not impair TNBC cell growth either in vitro or in vivo (Fig. 1).
In conclusion, the study provides compelling insights into the mechanism of action of POGK, highlighting how its isoform switching undermines cancer defense mechanisms. The discovery that reduced POGK expression in TNBC arises from a shift from a full‐length isoform to a shorter one is particularly interesting. Identifying the molecular players that regulate this isoform balance will be critical for developing therapeutic strategies to restore the tumor‐suppressive POGK iso1 expression. Splicing is a highly intricate process involving core spliceosome components and cofactors that promote or repress spliceosome assembly. Recent studies suggest that the deregulation of splicing‐related proteins is a primary driver of altered splicing landscapes in cancer [16, 17]. This raises an important question about the identity of splicing factors that induce the POGK isoform switch. TNBC, particularly its glycolytic subtype MPS2, associated with higher tumor grade, exhibits profound dysregulation of splicing machinery proteins [18]. This makes TNBC potentially vulnerable to splicing‐targeting pharmacological interventions. Indeed, modulation of specific splicing‐related genes has been shown to induce TNBC cell death [19, 20, 21]. With the growing focus on pharmacological inhibition of splicing factors [22, 23] and on isoform switching drugs [24], correcting POGK isoform imbalance in TNBC offers a promising strategy to combat this aggressive cancer subtype.
The authors observed that POGK levels are both upregulated and downregulated across various cancer cohorts, raising another important question. How do the tumor‐suppressive functions of POGK‐iso1 manifest in TNBC subtypes or other tumor types, and to what extent is POGK expression predictive of patient survival or treatment response?
From a genomic perspective, Tu et al. provide the first evidence of a DNA transposon‐derived fusion protein acting as a tumor suppressor. As the investigation of TE‐derived proteins continues to advance, additional narratives will likely emerge, shedding light on these molecular chimeras born from the complex interplay between host genomes and their parasitic invaders.
Conflict of interest
The authors declare no conflict of interest.
Author contributions
EZ and MB both analyzed and discussed the Tu et al. paper, wrote and edited the manuscript.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bourque G , Burns KH , Gehring M , Gorbunova V , Seluanov A , Hammell M , et al. Ten things you should know about transposable elements. Genome Biol. 2018;19:1–12.30454069 10.1186/s 13059-018-1577-z PMC 6240941 · doi ↗ · pubmed ↗
- 2Fueyo R , Judd J , Feschotte C , Wysocka J . Roles of transposable elements in the regulation of mammalian transcription. Nat Rev Mol Cell Biol. 2022;23:481–497.35228718 10.1038/s 41580-022-00457-y PMC 10470143 · doi ↗ · pubmed ↗
- 3Arribas YA , Baudon B , Rotival M , Suárez G , Bonté PE , Casas V , et al. Transposable element exonization generates a reservoir of evolving and functional protein isoforms. Cell. 2024;187:7603–7620.39667937 10.1016/j.cell.2024.11.011 · doi ↗ · pubmed ↗
- 4Pasquesi GIM , Allen H , Ivancevic A , Barbachano‐Guerrero A , Joyner O , Guo K , et al. Regulation of human interferon signaling by transposon exonization. Cell. 2024;187:7621–7636.e 19. 10.1016/j.cell.2024.11.016 39672162 PMC 11682929 · doi ↗ · pubmed ↗
- 5Feschotte C , Pritham EJ . DNA transposons and the evolution of eukaryotic genomes. Annu Rev Genet. 2007;41:331–368.18076328 10.1146/annurev.genet.40.110405.090448 PMC 2167627 · doi ↗ · pubmed ↗
- 6Zhang W , Wu J , Ward MD , Yang S , Chuang YA , Xiao M , et al. Structural basis of arc binding to synaptic proteins: implications for cognitive disease. Neuron. 2015;86:490–500.25864631 10.1016/j.neuron.2015.03.030PMC 4409568 · doi ↗ · pubmed ↗
- 7Mi S , Lee X , Li XP , Veldman GM , Finnerty H , Racie L , et al. Syncytin is a captive retroviral envelope protein involved in human placental morphogenesis. Nature. 2000;403:785–789.10693809 10.1038/35001608 · doi ↗ · pubmed ↗
- 8Kapitonov VV , Jurka J . RAG 1 core and V(D)J recombination signal sequences were derived from Transib transposons. P Lo S Biol. 2005;3:e 181.15898832 10.1371/journal.pbio.0030181 PMC 1131882 · doi ↗ · pubmed ↗