Metabolites managing excess manganese: The SgPAL2-regulated flavonoids in stylo
Munkhtsetseg Tsednee

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
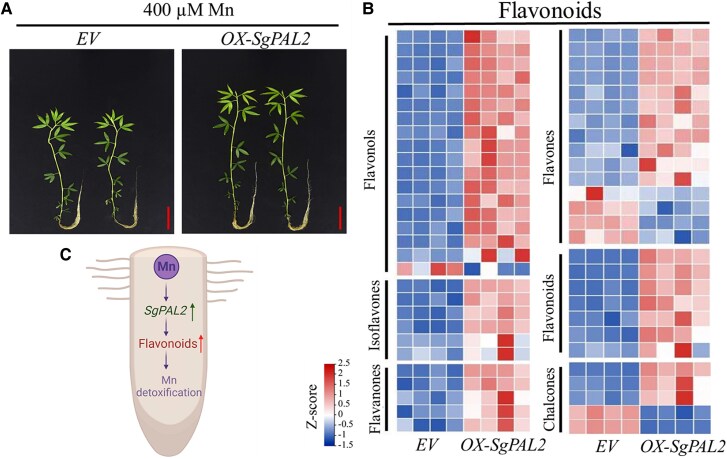 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhytochemical compounds biological activities · Plant Reproductive Biology · Fungal Biology and Applications
Plants require manganese (Mn) to support many biological processes such as respiration and photosynthesis. Mn is a cofactor of various enzymes, for example, Mn-superoxide dismutase (Alejandro et al. 2020). However, in excess, Mn causes oxidative stress, impaired photosynthesis, and disrupted enzyme activity, thereby reducing plant growth and productivity (Alejandro et al. 2020).
In acidic soil, Mn toxicity occurs commonly due to abundant presence of bioavailable Mn form (Mn^2+^) (Shao et al. 2017). Considering that 40% to 50% of the world's arable lands are acidic, located mainly in tropical and subtropical areas (Kochian et al. 2004), Mn toxicity challenges plant growth and development, especially crop production. But some plants, such as tropical legume stylo (Stylosanthes guianensis), tolerate excess Mn and adapt to acidic soils (Chen et al. 2015). Uncovering how these plants cope with Mn toxicity could provide insights into improving plant tolerance to excess Mn.
In Plant Physiology, Wang et al. (2025) investigated the Mn tolerance mechanisms in stylo genotypes and identified a metabolic regulatory gene and metabolites involved in Mn detoxification. Initially, the authors confirmed and worked with a Mn-tolerant genotype, RY5, that showed no defects in root growth and enhanced antioxidant activity in high Mn. These observations suggested that RY5 uses the internal Mn detoxification mechanism to tolerate high Mn contents.
Previously, specialized metabolites have been suggested to be involved in Mn tolerance in different plant species, for example, cowpea (Führs et al. 2012) and citrus (Zheng et al. 2024). Indeed, over 230 Mn-responsive metabolites could be identified in RY5 exposed to elevated Mn. Of these, the majority were flavonoids and phenolic compounds.
The authors used their earlier transcriptomic analyses in stylo with Mn supply (Jia et al. 2020) and looked at the mRNA accumulations of genes related to the phenylpropanoid/flavonoid pathway in both RY5 and TF0317 roots. From the RY5-specific genes, the authors then selected PHENYLALANINE AMMONIA-LYASE 2 (SgPAL2), whose expression increased over 4-fold in response to excess Mn, for its functional study.
Phenylalanine ammonia-lyase is a key enzyme in the phenylpropanoid pathway and catalyzes the conversion of phenylalanine to trans-cinnamic acid (Zhang and Liu 2015). PAL activity increased in RY5 roots, but not in TF0317 roots, in excess Mn treatment, and the inhibition of PAL activity decreased Mn tolerance in RY5. These results indicated the potential role of SpPAL2 in conferring Mn tolerance in the RY5 genotype.
To confirm the SpPAL2 function, Wang et al. (2025) further generated OX-SgPAL2 overexpression and RNAi-SpPAL2 knockdown lines. Although all of the lines showed similar levels of Mn accumulation, the OX-SgPAL2 lines tolerated the high Mn (Fig. 1A) compared with the RNAi knockdown lines. Moreover, the antioxidant enzyme activities and 16 genes related to the biosynthesis of secondary metabolites significantly increased in OX-SgPAL2 overexpression lines. Together, these results demonstrated that SgPAL2 functions in Mn tolerance in stylo, with an anticipated role in metabolic regulations.
Lastly, to identify the *SpPAL2-*regulated metabolites, the authors performed metabolomic analyses in OX-SpPAL2 overexpression versus control lines and observed the enrichments of metabolites from the phenylpropanoid/flavonoid pathway in the overexpression lines (Fig. 1B). They further obtained 25 candidate metabolites, including 4 flavonoids, those abundances were also increased in RY5 roots under high Mn.
An exogenous supply of one of these candidate flavonoids, calycosin, enhanced the stylo root growth under excess Mn and decreased the ROS level triggered by excess Mn. Thus, the results confirmed that the *SpPAL2-*regulated calycosin flavonoid achieves Mn detoxification by reducing ROS accumulation in stylo roots (Fig. 1C).
In summary, Wang et al. (2025) have identified the critical metabolic regulatory gene, SpPAL2, and metabolites associated with Mn tolerance in stylo. This finding advances the understanding of excess metal detoxification and tolerance mechanisms in plants. Moreover, the study provides a promising target and route to improve plant tolerance to excess Mn through engineering the phenylpropanoid/flavonoid pathway.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alejandro S, Höller S, Meier B, Peiter E. Manganese in plants: from acquisition to subcellular allocation. Front Plant Sci. 2020:11:300. 10.3389/fpls.2020.0030032273877 PMC 7113377 · doi ↗ · pubmed ↗
- 2Chen ZJ, Sun LL, Liu PD, Liu GD, Tian J, Liao H. Malate synthesis and secretion mediated by a manganese-enhanced malate dehydrogenase confers superior manganese tolerance in Stylosanthes guianensis. Plant Physiol. 2015:167(1):176–188. 10.1104/pp.114.25101725378694 PMC 4281010 · doi ↗ · pubmed ↗
- 3Führs H, Specht A, Erban A, Kopka J, Horst WJ. Functional associations between the metabolome and manganese tolerance in. J Exp Bot. 2012:63(1):329–340. 10.1093/jxb/err 27621934118 PMC 3245468 · doi ↗ · pubmed ↗
- 4Jia YD, Li XY, Liu Q, Hu X, Li JF, Dong RS, Liu PD, Liu GD, Luo LJ, Chen ZJ. Physiological and transcriptomic analyses reveal the roles of secondary metabolism in the adaptive responses of Stylosanthes to manganese toxicity. BMC Genomics. 2020:21(1):861. 10.1186/s 12864-020-07279-233272205 PMC 7713027 · doi ↗ · pubmed ↗
- 5Kochian LV, Hoekenga OA, Pineros MA. How do crop plants tolerate acid soils? Mechanisms of aluminum tolerance and phosphorous efficiency. Annu Rev Plant Biol. 2004:55(1):459–493. 10.1146/annurev.arplant.55.031903.14165515377228 · doi ↗ · pubmed ↗
- 6Shao JF, Yamaji N, Shen RF, Ma JF. The key to Mn homeostasis in plants: regulation of Mn transporters. Trends Plant Sci. 2017:22(3):215–224. 10.1016/j.tplants.2016.12.00528087151 · doi ↗ · pubmed ↗
- 7Wang L, Li J, Liu L, Dong R, Liu GD, Rao IM, Chen ZJ. Phenylalanine ammonialyase 2 regulates secondary metabolism and confers manganese tolerance in Stylosanthes guianensis. Plant Physiol; 2025:197(1). 10.1093/plphys/kiaf 00539761536 · doi ↗ · pubmed ↗
- 8Zhang XB, Liu CJ. Multifaceted regulations of gateway enzyme phenylalanine ammonia-lyase in the biosynthesis of phenylpropanoids. Mol Plant. 2015:8(1):17–27. 10.1016/j.molp.2014.11.00125578269 · doi ↗ · pubmed ↗